
滇产单子叶植物的多样性格局与保护
2024-03-19 08:14文妍莉
植物研究 2024年2期
文妍莉 李 嵘
(1.中国科学院昆明植物研究所,东亚植物多样性与生物地理学重点实验室,昆明 650201; 2.中国科学院大学,生命科学学院,北京100049)
生物多样性是指地球上所有生命体与其生存环境相互作用形成的生态复合体[1],它是人类赖以生存和发展的基础。全球生物多样性的分布极不均衡,热带和山地的生物多样性显著高于其他区域[2-3]。然而,由于人类活动、气候变化和资源过度利用等的影响,生物多样性正急剧减少[4],因此,揭示生物多样性的分布格局及其形成原因是阻止和逆转生物多样性丧失的重要途径之一。
物种丰富度是反映生物多样性丰富程度的重要参考指标,能准确揭示各地区的生物复杂性和多样性[5],例如,Liu 等[6]利用物种丰富度揭示了秦岭—大巴山南北坡的生物多样性聚集度。近年来,高通量DNA 测序技术的发展和区域生命之树的建立,为从系统发育的角度评估生物多样性的组成提供了方法和途径。系统发育多样性是最早用来标识生物多样性组成的进化指标[7],它用区域内所有物种在系统发育树上的最短枝长之和来度量。Zhou 等[8]基于樟科(Lauraceae)植物的系统发育关系,并结合其地理分布,探讨了中国樟科植物系统发育多样性的分布格局,揭示了植被过渡区具有较高的进化组成多样性。这类方法改变了以往将物种视为独立个体的做法,强调物种间的演化关系,因而更好地反映生物多样性在进化维度上的多样性[9-12]。
特有现象是生物多样性在进化过程中对不同区域自然环境适应和演化的结果[13],对于理解生物多样性的形成与保护具有重要意义。Huang等[14]根据中国特有种子植物的地理分布,并结合物种间的系统发育关系,识别出我国生物多样性保护的19个热点和空白区。加权特有性和系统发育特有性是常用的量化分类学和进化历史特有现象的指标[15-16]。
近年来,区域物种的受威胁状态在生物多样性保护领域受到越来越多的关注[17]。Hills等[18]利用进化特异性全球濒危度指数对南非薯蓣属(Dioscorea)植物多样性进行分析,识别了南非重要的生物多样性保护优先区。进化特异性全球濒危度指数是将物种进化特异性与物种濒危等级相结合[19],用以量化生物多样性的区域优先保护级别[20]。
随着全球数据的开放共享,综合生物多样性、特有性及濒危度,从多个视角理解生物多样性的形成与演变,可为生物多样性的保护提供多维证据。如Zhang 等[21]对中国西南地区的维管植物多样性进行研究发现,植物的多样性、特有性和濒危度的地理分布格局之间的重合度较低,这使得植物多样性的综合保护变得尤为困难。为更好地理解生物多样性的分布格局和形成原因,有必要从不同类群、不同维度进行综合研究,以便更精准地制定有效的生物多样性保护策略。
云南地跨喜马拉雅、印度—缅甸、中国西南山地3个全球生物多样性热点地区[22],富含丰富的珍稀濒危和特有生物物种[14,23-26]。植物谱系组成多样,包含木兰分支、单子叶植物、真双子叶植物等多个被子植物演化支系[27]。其中,单子叶植物是云南植物区系的重要组成部分,约占云南被子植物的15%[28]。因生活史短、进化速率高,其演化历史有别于其他被子植物分支[29],也因其适应性强,分布极为广泛,多是林下或林中树上的伴生植物,或是高山、亚高山草甸的主要建群植物[30]。本研究以云南单子叶植物为例,基于物种的地理分布及其系统发育关系,从分类学和系统发育的角度探讨单子叶植物的多样性、特有性及受威胁程度的分布格局,并整合云南自然保护地的分布,辨识单子叶植物的保护空缺,为云南生物多样性保护提供科学指导。
1 材料与方法
1.1 研究地区
云 南 地 处 中 国 西 南,位 于21°8′32″~29°15′8″N,97°31′39″~106°11′47″E。由于新近纪以来的造山运动,使得境内河谷深切、地势陡峭,整体自东南向西北抬升[31]。由于受太平洋季风和印度洋季风的双重影响,云南气候表现出年温差小、日温差大、雨量充沛、干湿季分明等特点[32]。古南大陆成分、古北大陆成分、古地中海成分及东亚成分是云南植物区系的主体[33]。植被类型多样,包括热带雨林、亚热带常绿阔叶林、针阔叶混交林、高山和亚高山灌丛和草甸等多种类型[30]。
1.2 数据来源
根据《云南植物志》[28]《云南省生物物种名录(2016版)》[34]等资料,对云南单子叶植物物种数据库进行补充完善,该数据库包括植物学名、中文名、科属位置、地理分布等信息。利用Flora of China(http://www.efloras.org/,FOC)、Plants of the World Online(https://powo.science.kew.org/,POWO)、World Flora Online(https://www.worldfloraonline.org/,WFO)、中国数字植物标本馆(https://www.cvh.ac.cn/,CVH)、中国国家标本资源平台(http://www.nsii.org.cn/,NSII)等线上数据库对植物学名、地理分布信息进行修订。科属范畴以被子植物分类系统为依据[27],物种学名以Flora of China 为标准,地理分布以行政县为单元。将亚种、变种等种下等级归并到相应的种内,同时排除栽培、归化和外来入侵物种。此外,根据《IUCN 物种红色名录的濒危等级和标准:3.1 版》[35],结合《中国高等植物受威胁物种名录》[36],将云南单子叶植物的受威胁状态归为易危、濒危和极危。
为减少县域面积大小对生物多样性度量的影响,使用地理信息系统ArcGIS 10.2 将云南划分为4 374个10 km×10 km 的等面积栅格。以云南省地图做底图,将植物县域分布记录根据经纬度匹配到相应的等面积栅格中,最终获得929 114 条物种栅格分布记录。底图矢量数据来源于国家基础地理信息中心(http://www.ngcc.cn/ngcc/)1∶100 万矢量地图数据,地理坐标参考系统为WGS_1984_UTM_Zone_47N。
1.3 系统发育树构建
基于GBOTB 系统发育树[37],利用R 3.6.2 中的V.PhyloMaker 程序包[38],根据云南单子叶植物物种数据库构建滇产单子叶植物系统发育树。参考V.PhyloMaker 中的Scenario 3 方法,对于骨架树中缺失的物种,根据其所在科或属的系统位置进行添加[38]。
1.4 多样性度量指标
采用物种丰富度、系统发育多样性、标准化系统发育多样性、加权特有性、系统发育特有性度量云南单子叶植物多样性。
物种丰富度(RS)为每个栅格内所有物种之和,计算公式如下:
式中:K表示栅格单元中的物种总数量。
系统发育多样性(DP)是指连接系统发育树根部到支端的枝长之和[7],计算公式如下:
式中:C是一组分支,表示一个栅格单元中所有类群到系统发育树根部的最短生成路径;c是一个分类单元的分支;Lc是分支c所代表的分支长度。
研究表明,物种丰富度和系统发育多样性显著相关[39],为消除物种丰富度对系统发育多样性的影响,采用标准化的方法量化每个栅格的系统发育组成,也即标准化系统发育多样性(DP,s)[40],计算公式如下:
式中:DP,o表示每个栅格内系统发育多样性的观测值,DP,r表示随机化每个栅格的系统发育多样性,σDP,r表示随机化后的每个栅格的系统发育多样性标准差。若DP,s为正,表明栅格系统发育组成多样性相对较高;反之,栅格系统发育组成多样性相对较低[41]。
加权特有性(EW)是指限定在特定范围的物种数量[15],根据物种的分布范围,为每个物种设置权重,分布范围越小,权重越高,计算公式如下:
式中:n表示栅格的物种数量;Wi表示i物种在该栅格内的权重。
系统发育特有性(EP)是栅格内每个分支分布范围的倒数加权的分支长度之和[16],计算公式如下:
式中:C是分类群到系统发育树根部的最短路径的分支集合;Lc是分支c的长度;Rc是分支c所有后代物种的全局范围大小,其范围大小用所占据的栅格单元总数来衡量,而不重复计算栅格。
使用受威胁指数(IT)和进化特异性全球濒危度指数(IEDGE)度量云南不同栅格单子叶植物的受威胁程度。
受威胁指数(IT)是每个栅格内受威胁物种数量占栅格内全部物种的比例,计算公式如下:
式中:RT为栅格内受威胁物种数量,即极危、濒危和易危物种的总和,RS为栅格内物种总数。
进化特异性全球濒危度指数是将各栅格内每一物种的进化特异性(ED)与全球受威胁等级(GE)进行综合分析得到的指标[19],计算公式如下:
式中:ED是基于GBOTB 骨架树构建的云南单子叶植物系统发育树计算获得的进化特异性值;GE是受威胁物种的分值(易危1分、濒危2分、极危3 分)。最后,根据栅格内每个物种的IEDGE,计算各栅格所有物种IEDGE的平均值作为各栅格单子叶植物的受威胁程度。
物种丰富度与系统发育多样性(标准化系统发育多样性)、加权特有性与系统发育特有性、受威胁指数与进化特异性全球濒危度指数的相关性分析采用Pearson 指标。上述所有分析利用R 3.6.2 软件中的函数cor()、picante 程序包[42]和Biodiverse 3.2软件完成。
1.5 保护空缺分析
将云南单子叶植物的多样性、特有性及受威胁程度的前30%和5%定义为热点区域,表示云南单子叶植物的重点保护区域。使用ArcGIS 10.2软件,在同一图层文件夹下依次打开物种丰富度热点区域、系统发育多样性热点区域、物种特有性热点区域、系统发育特有性热点区域、受威胁物种热点区域、进化特异性及全球受威胁度热点区域和云南自然保护地图层,热点区域图层中未被自然保护地所覆盖的区域即为云南单子叶植物的保护空缺。
2 结果与分析
2.1 云南单子叶植物的多样性格局
据统计,云南有单子叶植物35 科459 属2 267种及38 亚种和101 变种。其中,局限分布于云南的特有种有317种,受威胁物种436种(极危52种、濒危166种、易危218种)。
云南单子叶植物的物种丰富度与系统发育多样性显著正相关(R=0.99,P<0.001;见图1A),与标准化系统发育多样性显著负相关(R=-0.42,P<0.001;见图1B);云南单子叶植物的加权特有性与系统发育特有性显著正相关(R=0.98,P<0.001;见图1C);云南单子叶植物的受威胁指数与进化特异性全球濒危度指数显著正相关(R=0.64,P<0.001;见图1D)。云南西北部、南部和东南部的物种组成和系统发育组成多样性和特有性均较高(见图2A~D);然而,当消除物种丰富度对系统发育多样性的影响后,除西北部和东南部的标准化系统发育多样性较低外,其他区域的标准化系统发育多样性均较高(见图3)。
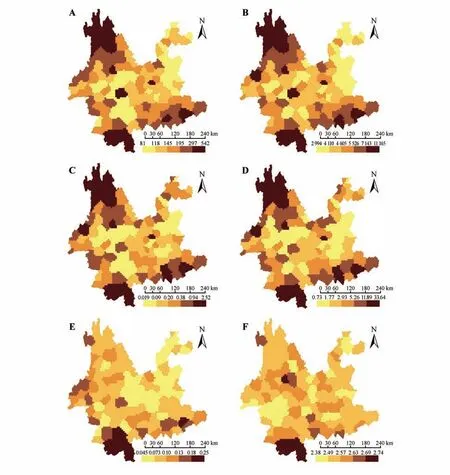
图2 云南单子叶植物的多样性格局A.物种丰富度;B.系统发育多样性;C.加权特有性;D.系统发育特有性;E.受威胁指数;F.进化特异性全球濒危度指数。Fig.2 Diversity patterns of Yunnan monocotyledons A.Species richness;B.Phylogenetic diversity;C.Weighted endemism;D.Phylogenetic endemism;E.Threatened index;F.Evolutionary distinctiveness and globally endangered index.
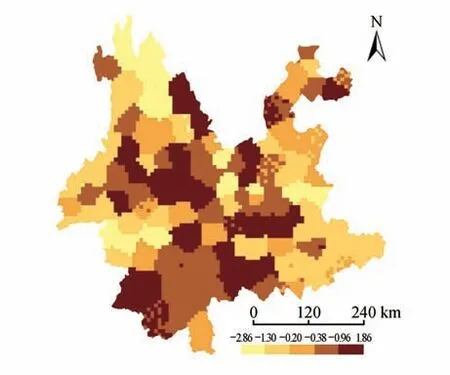
图3 云南单子叶植物的标准化系统发育多样性Fig.3 Standardized phylogenetic diversity patterns of Yunnan monocotyledons
2.2 区域受威胁状况与重点保护区域
Pearson相关性分析表明受威胁指数与进化特异性全球濒危度指数呈显著正相关(R=0.64,P<0.001),且二者的分布格局均显示云南西北部和南部是单子叶植物受威胁程度最为严重的区域(见图2E~F)。云南单子叶植物的多样性、特有性、受威胁程度与自然保护地的图层叠加显示,云南西北部、南部和东南部是单子叶植物的重点保护区域(见图4)。

图4 云南单子叶植物的重点保护区域Fig.4 Key protected areas of Yunnan monocotyledons
3 讨论
3.1 云南单子叶植物多样性的演化历史
研究表明云南单子叶植物的物种丰富度与系统发育多样性、加权特有性与系统发育特有性均呈显著正相关(见图1A,C),云南的西北部、南部和东南部是植物多样性和特有现象最为丰富的地区(见图2A~D),这与前期不同类群的研究结果一致,如云南被子植物[43]、云南被子植物蔷薇分支[25]、云南被子植物菊类分支[26]。然而,使用零模型去除物种丰富度对系统发育多样性的影响后,发现云南单子叶植物的谱系多样性在西北部和东南部较低,而在其他区域较高(见图3)。新生代以来,地质历史事件如印度板块与欧亚板块的碰撞,对云南西北部地理环境的塑造有着深远的影响,加之中新世以来印度洋西南季风的增强,更是为植物提供了多样的生存环境,成为百合属(Lilium)、鹿药属(Maianthemum)、豹子花属(Nomocharis)、贝母属(Fritillaria)等类群的分化中心[44],而这些类群大都来源于相近的谱系分支如百合科(Liliaceae),导致该区域单子叶植物的物种多样性丰富,而系统发育多样性却较低。云南南部和东南部地理位置特殊,地质环境和气候条件比较稳定,且未受到第四纪冰期—间冰期的影响[45],是诸多喜热喜湿类群,如豆蔻属(Amomum)、石斛属(Dendrobium)、蜘蛛报蛋属(Aspidistra)、芭蕉属(Musa)、万代兰属(Vanda)、水玉簪属(Burmannia)等的重要聚集地[46],因此,这些地区的物种组成较为多样。然而,云南东南部主要为喀斯特地貌,由于环境的过滤作用和植物自身的系统发育保守性[47],组成该区域的植物主要来自相同的进化支系,如天南星科(Araceae)的芋头属(Colocasia)、崖角藤属(Rhaphidophora)、石柑子属(Pothos)、犁头尖属(Typhonium)等,导致云南东南部单子叶植物的物种组成多样,但系统发育组成却较单一;云南南部地形复杂,为不同植物类群的生存和繁衍提供了良好的条件,这些植物不但种类丰富,如海芋属(Alocasia)、兜兰属(Paphiopedilum)、开口箭属(Campylandra)、姜属(Zingiber)、省藤属(Calamus)、龙竹属(Dendrocalamus),而且来源于不同的进化谱系,如天南星科(Araceae)、兰科(Orchidaceae)、百合科(Liliaceae)、姜科(Zingiberaceae)、棕榈科(Arecaceae)、禾本科(Poaceae)等,从而导致云南南部具有较高的单子叶植物物种组成和系统发育组成多样性。
3.2 云南单子叶植物多样性的保护
对受威胁指数、进化特异性全球濒危度指数的联合分析,发现云南西北部和南部是受威胁物种数量最多且受威胁程度最高的区域(见图2E~F)。已有研究显示,由于社会发展、人为干扰、外来物种入侵,尤其大型工程带来的生态环境破坏是植物受威胁的重要原因[48]。此外,Zhang 等[49]的研究表明,由于高额经济利益驱使,一些重要的野生植物自然资源,如石斛属(Dendrobium)和兜兰属(Paphiopedilum)的种群数量稀少且分布零星,人为采挖及栖息地破碎化是这些类群面临的最大威胁。除外界客观因素外,植物自身的特性,如生长缓慢、遗传变异、萌芽率低等[50]也是导致植物受威胁的主要原因。我国东北、华北分布的大花杓兰(Cypripedium macranthos)由于基因交流受限,群体内遗传多样性较低[51],导致种群濒临灭绝。另外,Yildiz 等[52]对土耳其珍稀特有贝母属植物(Fritillaria aurea)的繁殖生物学研究表明,高度自交不亲和及传粉者依赖性使其生存受到严重威胁。
区域性物种丰富度、系统发育多样性及植物受威胁程度与自然保护地的叠加分析表明,云南西北部、南部及东南部是滇产单子叶植物的重点保护区域(见图4)。这些地区不但植物的种类、特有化程度及系统发育组成较高,而且受威胁物种的种类组成和区域受威胁程度(东南部除外)也较高。为应对人类活动、气候极端事件等引起的第六次生物大灭绝,加强对这些地区的生物多样性保护势在必行。具体而言,云南南部的单子叶植物谱系多样性较高,即其现代植物区系构成含有较为多样的演化支系[53]。通过加强云南南部单子叶植物多样性的保护,不但可以保护物种、特有种及受威胁物种的多样性,还可以保护复杂多样的植物区系演化历史。而对于云南西北部和东南部来说,虽然单子叶植物的谱系多样性较低,但这些谱系或主要由新近起源的类群构成(如西北部)[54],或是古老植物类群的孑遗(如东南部)[55],新兴类群是未来演化的基础,古老孑遗类群是植物进化密码的载体。加强云南西北部、东南部植物多样性的保护,不仅可以保护植物的种类组成、特有化水平及受威胁物种的多样性,还可以最大化保护植物区系的演化密码和进化潜能。尽管目前自然保护地的规划尚不能涵盖所有重点保护区(见图4),但通过提升现有自然保护地的连通性、规划布局的全能性,仍可以对植物多样性实施多维度、综合性的保护(包括演化历史、进化潜能和演化密码的保护)。
3.3 演化历史在生物多样性保护中的作用
生物多样性是从宏观(生态系统多样性)到微观(遗传多样性)、从群体到个体的自然综合体[1],生物多样性保护是多层次、多维度的综合性保护[56]。然而,由于技术、方法、数据等因素的制约,早期生物多样性的保护主要关注物种维度和宏观尺度的保护,忽视了演化历史(遗传多样性)维度的保护。近年来,随着区域性生命之树的构建、全球数据的开放共享、机器学习、深度学习、人工智能等新技术、新方法的涌现[37,57],生物多样性保护既面临时代的机遇,也面对时代的挑战,不同保护策略的选择对保护结果具有根本性影响,基于演化历史确立的保护优先级和空白区域识别,有助于提高保护策略的效率[58]。如Shan等[59]利用系统发育指标,补充完善了国家重点保护野生药用物种名录;Yang等[60]结合系统发育方法,确立了我国种子植物多样性优先保护区域;因此,未来生物多样性的保护应在不摒弃物种层面的基础上,从演化历史的维度,加强大数据、新技术、新方法在生物多样性保护中的应用,开展综合性保护。
猜你喜欢
云南画报(2021年11期)2022-01-18
科技创新与应用(2021年31期)2021-11-09
云南画报(2020年12期)2021-01-18
云南画报(2020年9期)2020-10-27
红领巾·探索(2020年5期)2020-05-19
非公有制企业党建(2019年7期)2019-07-19
小学科学(学生版)(2018年9期)2018-09-21
家教世界(2017年11期)2018-01-03
英语学习(2015年2期)2016-01-30
弹箭与制导学报(2015年1期)2015-03-11
