
无籽刺梨及其近缘种叶绿体基因组序列比较分析
2024-03-19 04:41黄嘉城段晓盟张晓琳冯家玉甘沛华何承忠
福建农林大学学报(自然科学版) 2024年1期
纵 丹, 黄嘉城, 段晓盟, 张晓琳, 冯家玉, 甘沛华, 何承忠
(1.西南林业大学云南省高校林木遗传改良与繁育重点实验室;2.西南林业大学 西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224)
无籽刺梨(RosasterilisS. D. Shi),又名搭钩刺梨、光枝无子刺梨、安顺金刺梨等,是蔷薇科(Rosaceae)蔷薇属(Rosa)植物。其由于无法产生可育种子而得名,是贵州特有的刺梨种,主要分布在贵州安顺、兴仁等地[1-2]。其果实清爽可口,富含维生素C等营养物质,因此被称为“维C之王” 。无籽刺梨还富含多种氨基酸和矿物元素,具有较高的营养价值和开发利用价值。此外,无籽刺梨的根系发达,生长迅速,因此在退耕还林等工程造林中被视为优选树种[2-3]。
刺梨(R.roxburghiiTratt),又名缫丝花,同属蔷薇科蔷薇属植物。季祥彪等[4]基于形态学比较分析认为,无籽刺梨可能是一种自然杂交种,它与刺梨在叶型、果形和花序等方面高度相似。单瓣缫丝花(R.roxburghiif.normalisRehd et Wils)是缫丝花的变型,是其野生原始类型,两者的形态和解剖结构基本相似[5]。文晓鹏等[6]结合形态学特征和RAPD分子标记对刺梨及部分近缘种的亲缘关系进行探讨,结果显示,无籽刺梨与贵州缫丝花(R.kweichowensisT. T. Yu &T. C. Ku)的遗传距离最近,并推测无籽刺梨可能来源于贵州缫丝花的高度雄性不育种。陈睿等[7]基于33个形态学性状对8种野生蔷薇的亲缘关系进行了研究,聚类结果显示,刺梨与紫花重瓣玫瑰[Rosarugosaf.plena(Regel) Byhouwe]和无籽刺梨聚在一组。李旦等[8]基于AFLP分子标记和DNA条形码分析结果认为,刺梨与无籽刺梨是独立的2个种。邓亨宁等[9]基于叶绿体基因片段和核基因片段的综合分析结果认为,无籽刺梨起源于长尖叶蔷薇(R.longicuspisBertal)和刺梨的天然杂交种,且长尖叶蔷薇为母本。陈兴银等[10]基于形态学特征比较分析表明,无籽刺梨与贵州缫丝花和刺梨的亲缘关系较近,内转录间隔区(internal transcribed spacers,ITS)的序列分析则表明,无籽刺梨与长尖叶蔷薇聚为一支。Han et al[11]基于叶绿体全基因组构建最大似然(maximum likelihood,ML)树,结果表明,无籽刺梨与中国月季(R.chinensisJacq)或单瓣月季(R.chinensisvar.spontanea)亲缘关系较近。综合上述分析,不同研究方法得出的结果在无籽刺梨的分类地位存在一定的差异。
叶绿体是绿色植物进行光合作用和碳固定的场所。作为半自主性细胞器,叶绿体具有一套完整的基因组,其遗传方式以母系为主[12-13]。大多数植物的叶绿体基因组呈典型的四分体结构,包括两个反向重复区(inverted repeat region, IR)、大单拷贝区(large single copy region, LSC)和小单拷贝区(small single copy region, SSC)[13]。长度一般120~160 kb,但最新研究结果表明,部分植物的叶绿体基因组长度可达到170 kb[14-16]。与核基因组和线粒体基因组相比,叶绿体因基因组较小、不易发生重组、进化速率较慢且序列较为保守等特征,在探究物种起源、演化以及物种间亲缘关系分析等方面具有重要应用[17-20]。截至2022年12月,研究者已经对大约110个蔷薇属植物(包括种、变种、杂交种和品种)的叶绿体基因组进行了测序工作。
本试验采用比较基因组学的研究方法,对无籽刺梨、刺梨、贵州缫丝花、单瓣缫丝花、长尖叶蔷薇和中国月季的叶绿体全基因组进行了分析。研究内容包括编码基因的组成、单拷贝(single copy, SC)区与IR区边界的比较、简单重复序列(simple sequence repeats,SSR)的鉴定以及基因组范围内的高变区序列的鉴定。此外,基于叶绿体全基因组序列,重建了蔷薇属植物的系统进化树,旨在揭示无籽刺梨及其近缘种之间的亲缘关系,并为蔷薇属植物的系统进化研究提供重要的分子标记。
1 材料与方法
1.1 无籽刺梨及其近缘种叶绿体基因组序列
从NCBI基因组数据库(https://www.ncbi.nlm.nih.gov/)下载6种蔷薇属的叶绿体基因组序列。具体物种信息为:无籽刺梨(MW007387)、刺梨(KX768420)、贵州缫丝花(MZ261861)、中国月季(MH332770)、长尖叶蔷薇(MZ261890)、单瓣缫丝花(MZ261869)。
1.2 叶绿体基因组序列组成和编码基因
利用Geneious软件分别确定6种植物叶绿体基因组序列LSC、SSC和IR区的长度,GC含量以及编码基因的类型和数目。
1.3 IR区边界的收缩与扩张分析
利用IRScope工具(https://irscope. shinyapps. io/irapp/)[21]对各区域连接位点进行可视化分析,直观呈现6个叶绿体基因组边界区域在IR、SSC和LSC之间的变化。
1.4 SSR分析
利用MISA软件(http://misaweb.ipk-gatersleben.de/)分析6种植物叶绿体基因组SSR位点。参数设置:单核苷酸最低重复次数为10次,二核苷酸最低重复次数为6次,三核苷酸、四核苷酸、五核苷酸和六核苷酸的重复次数均为5次。
1.5 叶绿体基因组高变区分析
采用DnaSP V5软件[22]并利用滑动窗口分析的方法计算蔷薇属40个叶绿体基因组(包括37种植物)(表1)的核苷酸多态性(nucleotide diversity)Pi。参数设置为窗口长度600 bp,步长200 bp,基于R语言绘制折线图,同时结合Geneious软件分析蔷薇属植物叶绿体基因组内高变区片段。
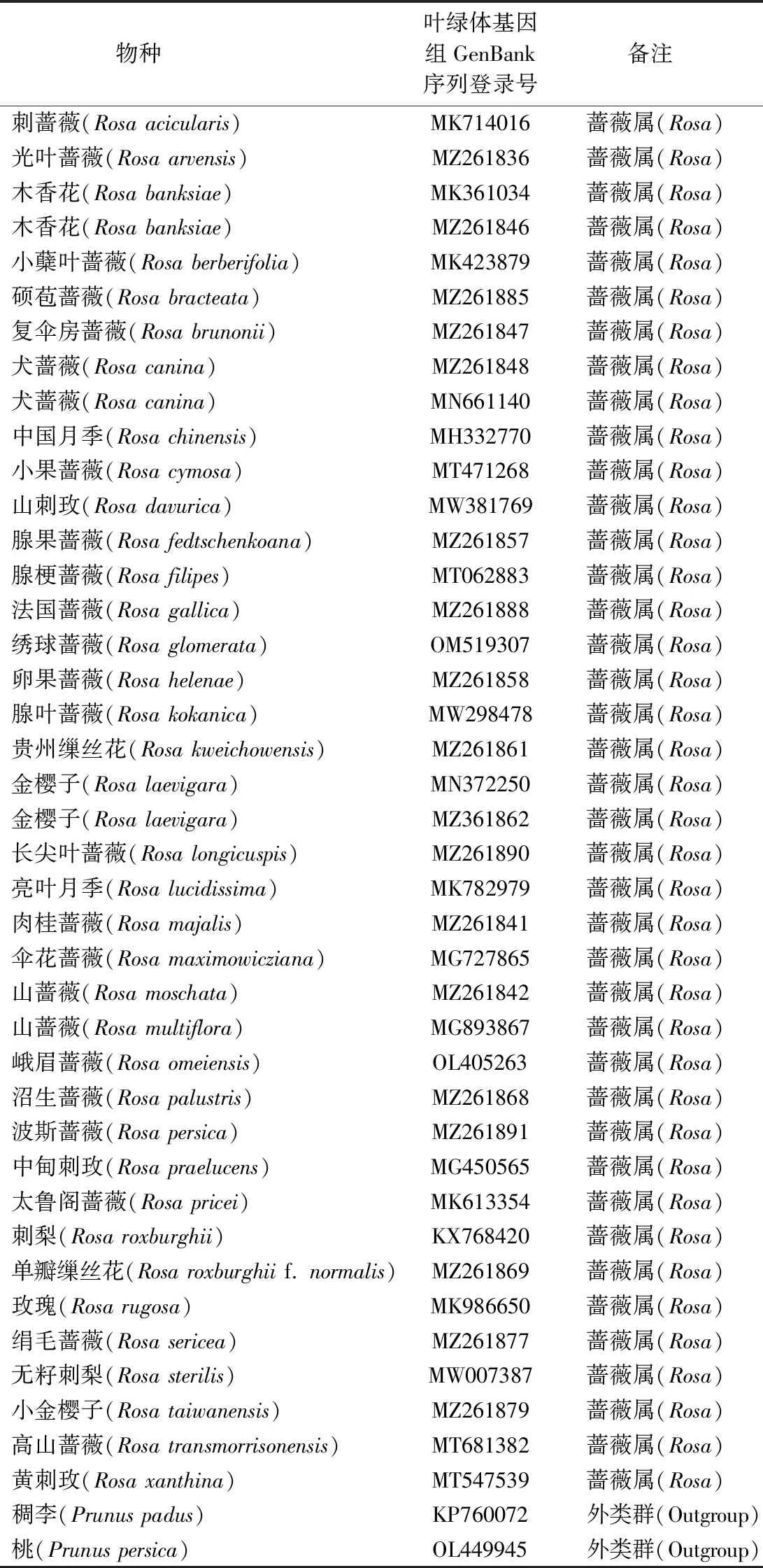
表1 用于高变区及系统发育分析的叶绿体基因组Table 1 Chloroplast genome of Rosa species for hypervariable region and phylogeny analysis
1.6 蔷薇属植物的系统进化树构建
从NCBI基因组数据库另外下载蔷薇属34种植物的叶绿体基因组序列,以蔷薇科稠李属(Prunus)的稠李(P.padus)和桃(P.persica)作为外类群(表1),采用MAFFT软件[23]对叶绿体基因组序列进行比对分析,其结果经过BioEdit软件进一步优化处理。优化后采用IQ-TREE软件构建蔷薇属植物的ML树[24],基于IQ-TREE内置的ModelFinder[25]筛选出建树的最佳模型TVM+F+R9,重复次数为1 000,最后将建树结果采用Figtree软件进行美化。
2 结果与分析
2.1 叶绿体基因组大小与结构
从NCBI基因组数据库下载了6种植物(无籽刺梨、刺梨、贵州缫丝花、中国月季、长尖叶蔷薇和单瓣缫丝花)的叶绿体基因组序列,并对其序列和编码基因进行了统计和比较(表2)。这些蔷薇属植物的叶绿体基因组长度为156 561~156 749 bp。其中,刺梨的叶绿体全基因组序列最长,为156 749 bp,而无籽刺梨和贵州缫丝花的叶绿体全基因组序列最短,为156 561 bp。与长尖叶蔷薇相比,无籽刺梨和贵州缫丝花的叶绿体全基因组序列长度分别短了95和125 bp,而与中国月季相比仅相差30 bp。这6种蔷薇属植物的叶绿体基因组的平均GC含量均为37.2%。另外,LSC、SSC和IR区长度分别为85 701~85 852 bp、18 746~18 792 bp和26 044~26 063 bp,IR区GC含量均高于LSC区和SSC区(表2)。
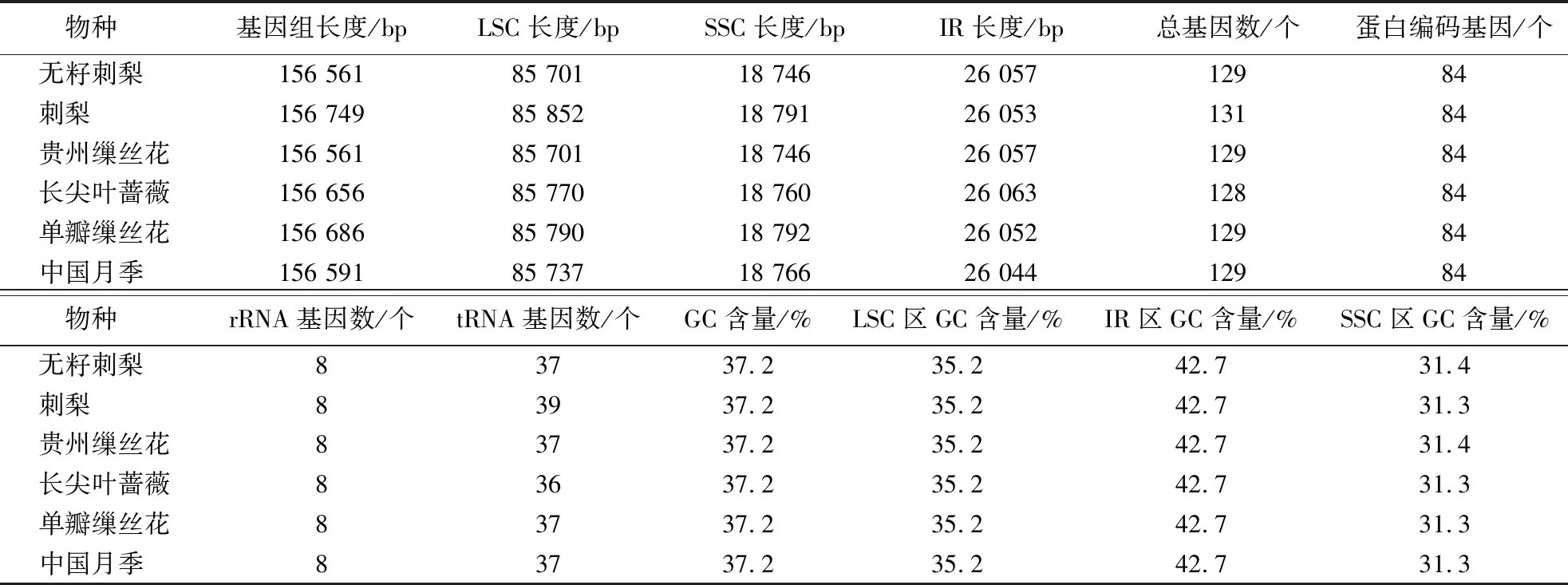
表2 6种蔷薇属植物的叶绿体基因组Table 2 Summary of chloroplast genome sequences of 6 Rosa species
经过对6种植物叶绿体基因组编码基因进行比较分析,发现无籽刺梨、贵州缫丝花、单瓣缫丝花和中国月季共编码129个基因,包括84个蛋白编码基因、8个rRNA基因和37个tRNA基因。而长尖叶蔷薇共编码128个基因,刺梨共编码131个基因。与前述的4种植物相比,长尖叶蔷薇的差异在于tRNA基因数目少1个,而刺梨的差异在于tRNA基因数目多2个,共有39个tRNA基因。
2.2 IR区边界的收缩与扩张
IR区作为叶绿体基因组中最保守的区域,其边界的扩张和收缩可导致叶绿体基因组长度发生改变[26]。本试验将6种植物的叶绿体基因组的IR/SSC区的边界情况进行了比较分析(图1)。结果表明:这些植物的4个边界基因类型和排列顺序完全一致,未出现基因的扩张或收缩,SSC/IRb边界(junction of SSC/IRb, JSB)和SSC/IRa边界(junction of SSC/IRa, JSA)均位于ycf1基因内部,且向IR区扩张的长度均为1 106 bp;除了单瓣缫丝花的rps19基因距离LSC/IRb的边界(junction of LSC/IRb, JLB)为15 bp外,其余树种的距离均为14 bp。此外,LSC/IRa区的边界(junction of LSC/IRa, JLA)位于trnH附近,单瓣缫丝花的JLA与trnH的距离为9 bp,而无籽刺梨、刺梨、中国月季、长尖叶蔷薇和贵州缫丝花JLA与trnH的距离分别为3、3、3、2、2 bp,表明无籽刺梨及其近缘种之间的叶绿体基因组结构具有高度保守性。
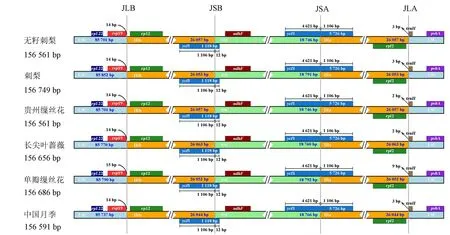
图1 叶绿体基因组边界分析Fig.1 Analysis on chloroplast genome boundary
2.3 SSR
利用MISA程序对无籽刺梨及其近缘种叶绿体基因组中SSR序列进行检测(图2),在6个物种的叶绿体基因组中,发现了3种类型的SSR序列重复基序,分别是单核苷酸(P1)、二核苷酸(P2)和复合型(C)。其中,单核苷酸重复类型最多,主要由A和T碱基重复组成,二核苷酸重复类型相同且均为AT碱基重复,表明无籽刺梨及其近缘种具有较强的AT偏向性。对无籽刺梨及其近缘种叶绿体基因组SSR序列进行比较分析表明,6种植物SSR类型和数目高度相似,除无籽刺梨和贵州缫丝花的SSR位点数量为50个(包括38个P1+3个P2+9个C)外,其他4种植物的SSR总数均为49个(图2)。
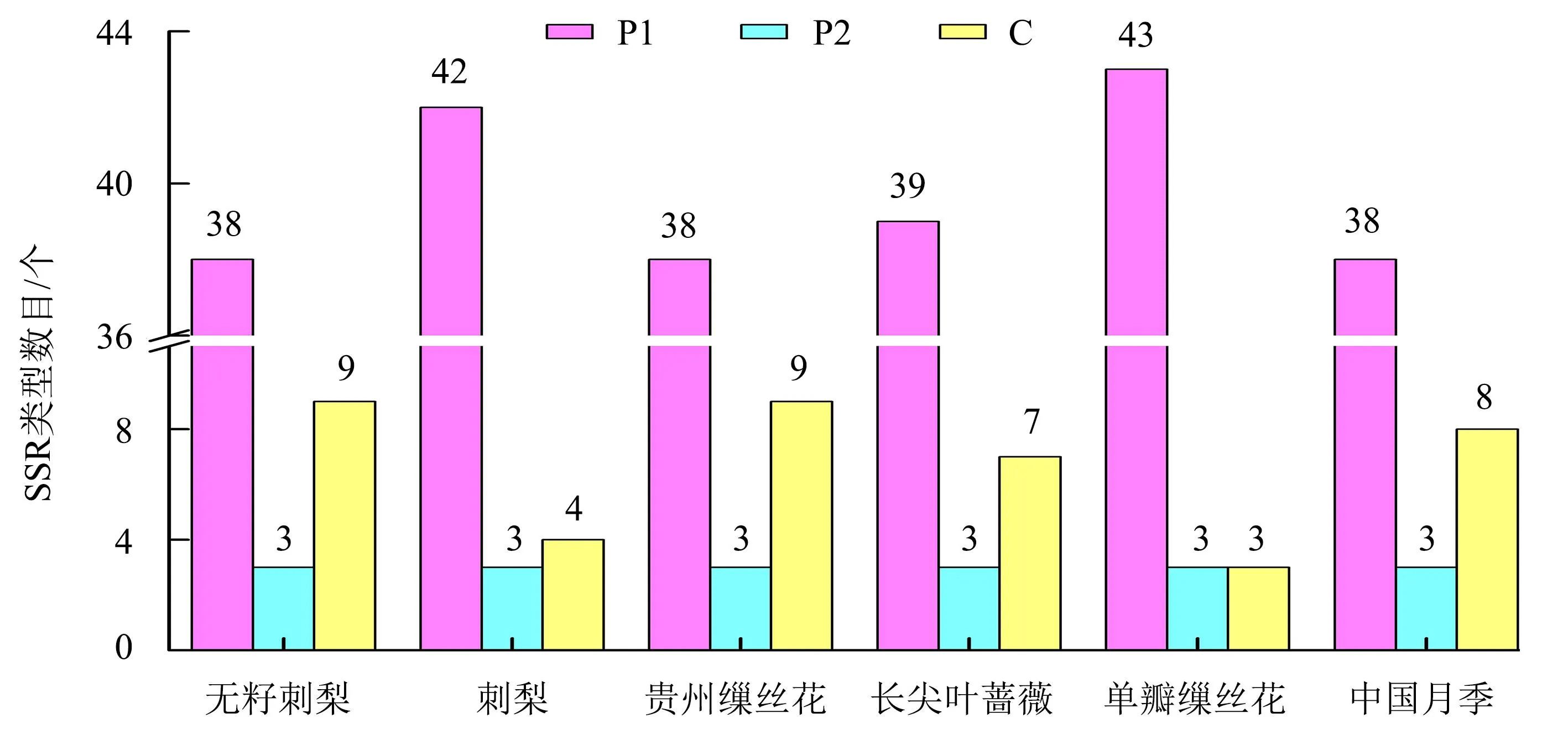
P1:单核苷酸SSR;P2:二核苷酸SSR;C:复合型SSR。
2.4 核苷酸多样性
采用DnaSP软件对40个蔷薇属植物的叶绿体基因组进行变异位点分析,发现共有3 753个多态位点。不同区段的叶绿体基因组Pi为0~0.015 6,平均值0.003 4。SC区变异核苷酸多样性高于IR区(图3),以Pi>0.01为阈值,筛选出了7个核苷酸变异较大的区段,包括trnK-trnQ、psbM-trnY、ycf3-rps4、rps4-trnL、psbE-petG、rpl16 intron和ycf1。除ycf1位于SSC区外,其余均位于LSC区。7个高变区片段中,5个为基因间隔片段,1个为内含子片段,1个为蛋白编码基因片段。这说明基因间隔区域表现出比编码区更高的核苷酸多样性。
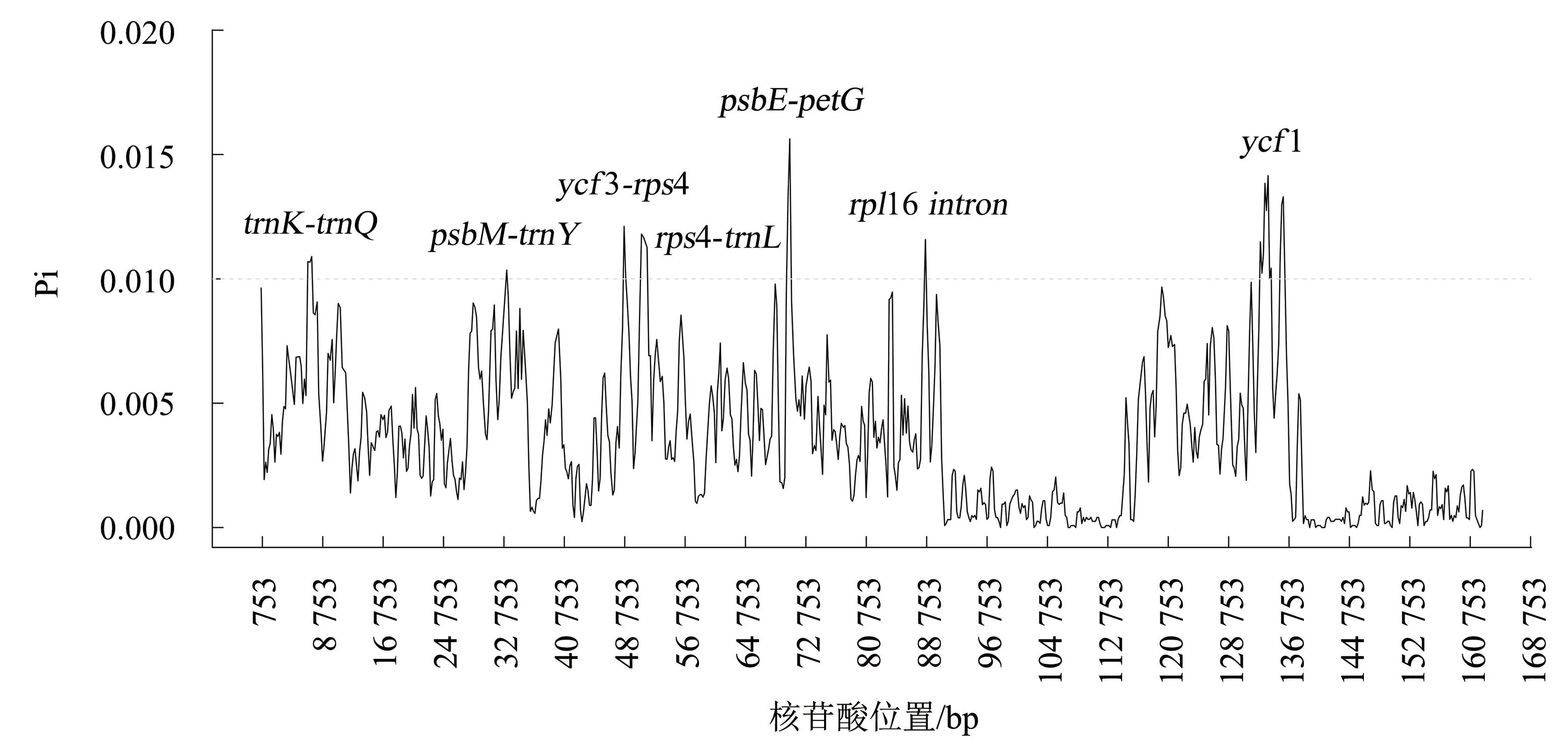
图3 蔷薇属40个物种的核苷酸多样性分析Fig.3 Analysis on nucleotide diversity of 40 Rosa species
2.5 系统发育
本试验共涉及40种蔷薇属植物,包括无籽刺梨及其近缘种。对这些植物的叶绿体全基因组进行了研究,并结合7个高变区片段的联合矩阵,利用ML法构建了系统发育树(图4和图5)。结果表明,基于高变区片段联合矩阵构建的系统发育树与基于叶绿体全基因组构建的系统发育树的拓扑结构完全一致。两者均表明无籽刺梨和刺梨位于不同的分支,为2个独立的种。其中,刺梨与单瓣缫丝花聚为一个分支,互为姊妹关系;无籽刺梨、贵州缫丝花、中国月季和长尖叶蔷薇聚为一支,且无籽刺梨与贵州缫丝花聚为一个高支持率的小支(支持率100%),并互为姊妹关系。

图4 基于叶绿体全基因组构建的系统发育树Fig.4 Phylogenetic tree based on complete chloroplast genome
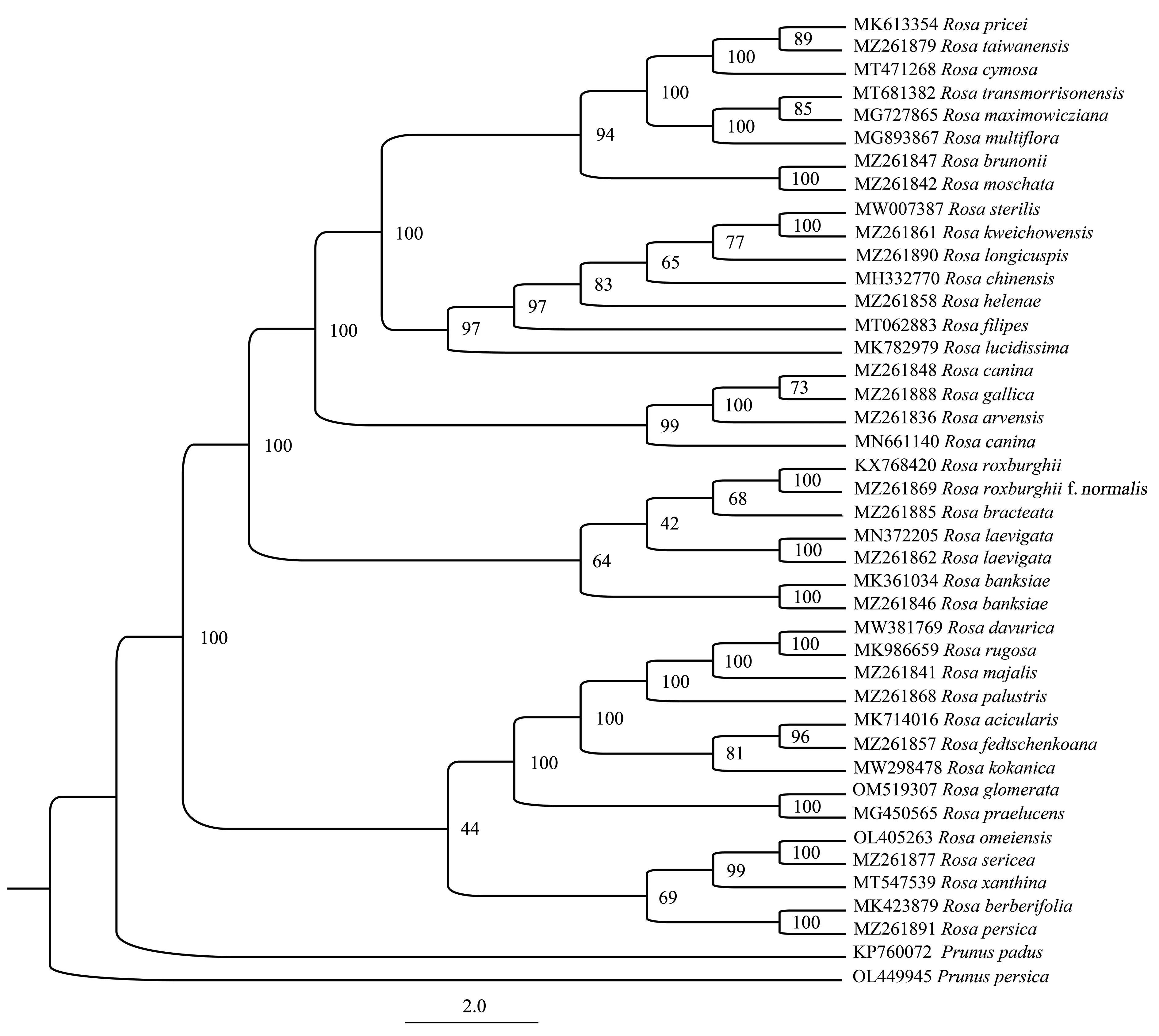
图5 基于高变区序列构建的系统发育树Fig.5 Phylogenetic tree based on hypervariable regions
3 讨论
植物叶绿体基因组通常由LSC、SSC、IRa和IRb组成,蔷薇属植物叶绿体基因组与绝大多数植物一样,呈闭合环状双链结构,具有典型的四分体结构[27]。本研究对无籽刺梨、刺梨、贵州缫丝花、中国月季、长尖叶蔷薇和单瓣缫丝花的叶绿体基因组进行了比较。结果表明,无籽刺梨及其近缘种之间叶绿体基因组高度相似,基因组长度为156 561~156 749 bp,总GC含量均为37.2%,且IR区GC含量高于LSC区和SSC区,4个rRNA基因位于IR区,这可能是导致IR区GC含量较高的原因[28]。IR区是叶绿体基因组中最保守的区域,在植物叶绿体基因组进化过程中,IR边界的收缩与扩张决定着植物叶绿体基因组的长度[26],此外,IR区的收缩和扩张频率也可反映物种之间的进化关系[29-30]。在本研究中,对这6种蔷薇属植物的叶绿体基因组进行了详细比较,发现各区域的稳定性较高,IR区的收缩或扩张对基因组长度影响较小。因此,推测这些植物叶绿体基因组大小的差异可能是由基因间隔区的插入或缺失所引起的[31]。
叶绿体基因组中的SSR(cpSSR)具有高复制率、共显性遗传、高多态性和母系遗传等特点,因此在物种鉴定、目的基因标记和系统发育研究中得到广泛应用[32-33]。对无籽刺梨及其近缘种cpSSR进行比较分析表明,6种植物的SSR类型和数目基本一致。其中,最常见的SSR位点是单核苷酸,主要由A/T组成,二核苷酸均为AT重复类型,进一步表明无籽刺梨及其近缘种叶绿体基因组偏好使用A或T碱基[34]。
在植物物种鉴定中,常选择进化速率较快的叶绿体基因片段和核糖体的ITS作为DNA条形码[35]。然而,蔷薇属植物叶绿体基因组高度保守的结构、基因数目和顺序,导致其基因或结构标记的变异程度较低,限制了蔷薇属物种亲缘关系的分析[26,36-37]。为了解决这个问题,Wu et al[38]和Wissemann et al[39]分别采用rbcL、matK、ndhC-trnV、atpB-rbcL、psbJ-petA、ndhJ-trnF、ndhF-rpl32作为DNA条形码分析蔷薇属系统发育关系,但部分序列的变异率较低,无法有效区分近缘种。因此,筛选变异率较高的高变区片段对于近缘种的鉴定和系统发育分析非常必要。本研究对40个蔷薇属植物叶绿体基因组进行比较分析,共检测到7个高变区片段,其中ycf1在其他植物中也具有较高的变异[40]。本试验将获得的这7个高变区片段联合进行系统发育树的构建,结果与全基因组序列构建的系统进化树一致,证明了这些高变区片段在蔷薇属物种亲缘关系研究中的有效性,可以进一步利用这些高变区片段作为候选标记,进行蔷薇属低分类阶元的系统发育和进化研究[41]。
本试验结果表明,无籽刺梨和刺梨是2个独立的种。其中,无籽刺梨与贵州缫丝花、长尖叶蔷薇和中国月季聚为一支,且贵州缫丝花与无籽刺梨互为姊妹关系,刺梨与单瓣缫丝花互为姊妹关系,此结果与文晓鹏等[6]、李旦等[8]研究结果一致;但与陈兴银等[10]、邓亨宁等[9]研究结果略有差异。这种差异可能源于样本数量的不同。此外,与单个叶绿体基因片段相比,完整的叶绿体基因组可以提供更多的信息位点,从而更有助于研究低分类水平物种之间的亲缘关系。这也可能是导致差异的原因之一[42]。
猜你喜欢
大众科学(2021年10期)2021-12-23
大众科学(2021年10期)2021-12-23
绿色科技(2020年5期)2020-05-29
四川蚕业(2020年4期)2020-02-10
现代园艺(2017年21期)2018-01-03
食品界(2017年7期)2017-08-24
饮食与健康·下旬刊(2016年12期)2017-01-05
儿童故事画报(2016年6期)2016-07-14
蚕桑通报(2015年1期)2015-12-23
丝绸(2014年4期)2014-02-28
