
野生藏酋猴的体重及其影响因素
2024-02-26 10:36孙宇夏梦怡夏颖娜汪京京卢家凯刘申岐夏东坡王希孙丙华
兽类学报 2024年1期
孙宇 夏梦怡 夏颖娜 汪京京 卢家凯刘申岐 夏东坡 王希 孙丙华*
(1 安徽大学资源与环境工程学院,合肥 230601)(2 安徽省生物多样性与短尾猴行为生态学国际联合研究中心,合肥 230601)(3 安徽大学生命科学学院,合肥 230601)
灵长类动物的身体形态特征能够体现生理代谢、营养状态、生长发育和系统发生等重要信息,因此受到进化生物学、生理生态学、保护生物学和动物生态学等研究领域的极大关注 (Kagayaet al., 2010)。体重是灵长类动物的主要体征指标之一,与物种特征 (Fleagle, 2013)、健康状况、生殖发育和能量代谢密切相关 (邹如金等,1999)。同时,体重对灵长类动物的行为和社会结构也有着重要影响,体重的差异将导致种内竞争和配偶选择策略的不同,以及社会等级结构的形成 (Silket al., 2003)。因此,分析野生灵长类动物的体重及其动态变化规律,不仅可以了解体重所隐含的物种特征和个体健康状态的生物学信息,也有利于理解动物的个体发育规律和行为生态适应机制,对野生灵长类动物保护与管理具有重要指导意义(Struhsaker, 2010; Wich and Singleton, 2017)。
灵长类动物的体重与个体发育阶段、食物资源、繁殖周期和健康状态都有直接的关系,受形态、生理、生态和进化等多种因素的影响。在灵长类动物群体内部,体重的动态变化是一种普遍现象,这在很大程度上取决于个体获取食物资源的种类、分布和质量 (Mori, 1979)。生活在食物资源季节性变化环境中的灵长类,其体重也表现出明显的季节波动,食物资源丰度和质量较高的季节,通常也是个体拥有最大体重的时期 (Lewis and Kappeler, 2005)。另外,个体对高质量食物资源的占有能力对其体重也具有一定的影响。例如当食物资源有限时,高顺位个体能够获得更多的高质量食物资源,这使得它们拥有更大的体重。在猕猴 (Macaca mulatta) (Sueuret al., 2012) 和东非狒狒 (Papio anubis) (Packer, 1979) 的社群中,雌雄个体的体重都与他们的社会顺位呈正相关。但在黑猩猩 (Pan troglodytes) 中,仅雌性个体的体重与社会顺位存在正相关关系 (Wrangham, 1977),暗示社会顺位对灵长类体重的影响可能存在更多的社会生态因素。体重的性二型现象在灵长类动物中也普遍存在,通常雄性体重显著大于雌性,且一雄多雌交配模式的物种比单配制的种类表现出更明显的雌雄性二型现象 (Payseuret al., 1999)。体重的这种性二型现象与个体发育、栖息环境、食物资源和系统发育等因素密切相关 (Leigh and Shea,1995; Rehg and Leigh, 1999)。
中国现有28 种灵长类动物,是世界上野生灵长类种类最为丰富的国家之一。以往针对中国野生灵长类的体重监测和研究较少,仅对藏酋猴(Macaca thibetana)、川金丝猴 (Rhinopithecus roxellanae) 和猕猴等物种的体重进行了初步监测与报道 (梁冰等,2001;赵远等,2012),有关我国野生灵长类不同物种的体重数据、动态变化规律及其影响因素的研究仍待加强。藏酋猴是中国特有的灵长类动物,隶属于猴科 (Cercopithecidae) 猕猴属(Macaca),已被列入《国家重点野生动物保护名录》(Ⅱ级) 和《世界濒危动物红皮书》(易危等级)。该物种具有混交型的社会体制,表现出全年交配、季节性繁殖特点 (李进华,1999)。前期基于黄山部分自然死亡个体的监测数据表明,藏酋猴是猕猴属中体重最大的物种 (李进华,1999)。研究发现,峨眉山的藏酋猴体重与栖息地食物资源的季节波动密切相关 (Zhao, 1994)。迄今为止,在群体水平上,仍缺乏藏酋猴体重的长期监测数据,限制了对该物种的体重发育动态及其与社会生态因素间关系的理解。
本研究以黄山野生藏酋猴YA1 自然社会群体为对象,探讨群体水平下的野生藏酋猴体重变异及其影响因素。自1987 年以来,研究团队持续跟踪研究YA1 群体近40 年,积累了大量的群体和个体生活史资料。群体内的年龄结构清楚,可准确识别个体,便于行为和体重数据的采集。从出生到性成熟,藏酋猴历经了婴幼期、少年期和青年期3 个发育阶段。至青年期 (雌猴3 ~ 5 岁,雄猴4 ~ 7岁) 时,出现性二型,雄性迅速长大,长出尖锐的犬齿,雌雄个体参与社会交往的时间增加,如给成年个体提供更多的理毛。基于以往的研究,我们假设性别、年龄、社会顺位和季节波动对藏酋猴的体重大小具有显著影响。本文利用持续监测YA1 群内不同个体11 个月的体重和行为数据,结合已有的群体和个体生活史资料,拟检验以下3 个预测:(1) 藏酋猴体重存在雌雄性二型现象,成年雄性体重显著高于成年雌性,未成年雌雄个体的体重发育存在分化,雌性的快速发育期早于雄性;(2) 藏酋猴成年雌雄个体的体重与其社会顺位间存在显著的正相关关系;(3) 藏酋猴的体重在不同季节间存在显著差异,由于秋季和冬季是食物资源最丰富和最匮乏的两个时期,藏酋猴秋季和冬季的体重分别显著高于和低于其他季节。本研究旨在利用非损伤性的测量方法,获得藏酋猴在群体水平上的体重数据,分析未成年的体重发育动态,探讨影响体重大小的主要因素,为藏酋猴的保护和管理提供基础数据支撑。
1 研究方法
1.1 研究地点和对象
本研究于2022年7月至2023年5月在安徽黄山野生猴谷 (北纬30°29′,东经118°10′) 完成。研究区域平均海拔高度500 m,其中最高海拔1 310 m。植被分布垂直变化明显,海拔600 ~ 800 m以次生常绿阔叶林为主;海拔600 ~ 800 m以落叶、常绿阔叶混交林为主;海拔1 000 m 以上,上层为落叶林,下层为常绿林。气温存在季节和空间差异,山顶气候呈寒温带特征,全年平均气温7.8 ℃,最高25.6 ℃,最低-19.8 ℃;山腰为温带,山脚为亚热带,全年平均气温15.3 ℃,最高34.2 ℃,最低-13.9 ℃ (黄山志编纂委员会,1988)。研究对象为鱼鳞坑1号群体 (YA1群),由61只藏酋猴组成,包括27只成年个体 (雄性12只,年龄7 ~ 15岁;雌性15 只,年龄5 ~ 19 岁) 和34 只未成年个体 (雄性22只,年龄0 ~ 6岁;雌性12只,年龄0 ~ 4岁)。由于不同年龄、性别的藏酋猴在体型和毛色上有典型的不同,因此可根据体型大小、体表毛发和面部特征,并辅以明显的个体特征 (断指、兔唇等),对该群藏酋猴进行准确的个体识别 (李进华,1999)。
1.2 数据收集
利用带有蓝牙功能的地磅秤 (测量范围0 ~300 kg,精确度0.1 kg)、三脚架、摄像机和自制地磅秤平台,完成藏酋猴体重数据的采集。测量体重时,利用少量玉米引诱目标个体到地磅秤台面,当其身体全部位于台面上且示数稳定3 s 以上时,进行读数并记录。整个测量过程不需要捕获或接触藏酋猴个体,对其生活无任何影响。处于社会边缘的雄猴和婴猴胆小怕人,体重测量难度较大,故研究对象共包括24 只成年个体 (雄性9 只,年龄7 ~ 15岁;雌性15只,年龄5 ~ 19岁) (表1) 和24只未成年个体 (雄性16 只,年龄0 ~ 6 岁;雌性8 只,年龄0 ~ 4 岁)。记录成年藏酋猴间的攻击-屈服行为,将行为数据导入Excel,构建雄性和雌性的攻击-屈服矩阵 (Aggression-submission matrix),然后基于此矩阵计算DS (David’s score) 值,通过DS值确定雄性和雌性个体的等级顺位,DS值越大,等级顺位越高 (表1)。将测量个体体重数据、测量时间录入Excel表格,用于后续分析。

表1 藏酋猴目标个体的年龄、性别和等级顺位Table 1 Information of age, gender and social rank of target animal individuals of Tibetan macaques
本研究的数据收集和研究内容符合《中华人民共和国野生动物保护法》的相关规定和安徽省黄山市林业局的监管要求。
1.3 统计分析
使用广义线性混合模型 (GLMM) 分析性别、年龄、顺位和季节因素对成年藏酋猴体重的影响,并进一步检验各因素对藏酋猴体重变化的影响。由于处在生长发育阶段的未成年个体体重变化较大,故未做分析。对同一年龄不同性别的藏酋猴体重数据,采用Kolmogorov-Smirnov (K-S) 检验数据是否符合正态分布,若符合正态分布,使用独立样本T检验,若不符合正态分布,使用Mann-WhitneyU检验探究同一年龄组不同性别间的体重是否存在差异;使用Spearman 相关性分析,验证体重与年龄和等级顺位的关系;使用Kruskal-WallisH检验分析不同季节间成年藏酋猴体重是否存在显著变化,进一步使用配对检验分析两季节间藏酋猴体重是否存在显著性差异。以上检验均为双尾检验,显著性阈值设置为0.05,文中体重数据展示均为平均值 ± 标准差 (mean ± SD)。
所有统计分析均在SPSS 27.0 和R 4.1.3 中进行,使用Origin 2023绘图。本研究始于2022年7月,经历夏季、秋季、冬季和春季,故文中与季节因素相关的图表均按照自然的季节顺序展示。10月未追踪到猴群,所以该月份体重变化数据缺失。
2 结果
2.1 藏酋猴体重变化
雄性和雌性成年藏酋猴的平均体重分别为(18.71 ± 2.33) kg 和 (11.79 ± 1.97) kg;未成年雄性(0 ~ 6岁) 的体重变化范围是0.60 ~ 14.20 kg,未成年雌性 (0 ~ 4岁) 的体重变化范围是 0.60 ~ 8.90 kg。结果表明,在群体水平上,年龄 (P< 0.001)、性别 (P< 0.01)、等级顺位 (P< 0.001) 和季节 (P<0.001) 对成年藏酋猴体重变化均存在显著影响。
同一个体的10 个月数据分析结果表明,调查期间藏酋猴的体重均呈现出先上升后下降再上升的趋势,成年雌雄个体体重在不同季节有波动,但总体趋势不变;未成年个体体重在10 个月内亦有波动,但总体呈增长的趋势 (图1)。

图1 藏酋猴体重的月份变化Fig. 1 Monthly variation in body weight of Tibetan macaques
2.2 藏酋猴体重与年龄的关系
未成年藏酋猴的体重增长速率在不同年龄组间存在显著变化 (雄性:H= 300.83,df= 5,P<0.001;雌性:H= 187.16,df= 3,P< 0.001)。2 岁前,雄性的体重显著高于同龄雌性;2 ~ 3 岁,雌性体重快速增加,逐渐超过同龄雄性;3 岁后,雄性体重快速增加,又再次显著高于同龄雌性。未成年不同年龄段雌雄个体体重增长速度也存在显著性差异,未成年雄性个体,1 ~ 3 岁体重增长速度较慢,3 ~ 5岁增长速度加快,为快速发育期,5 岁后增长速度减慢;未成年雌性个体,1 ~ 2 岁体重增长速度较为缓慢,2 ~ 3 岁增长较快,为快速发育期,3 岁后增长速度下降 (图2)。
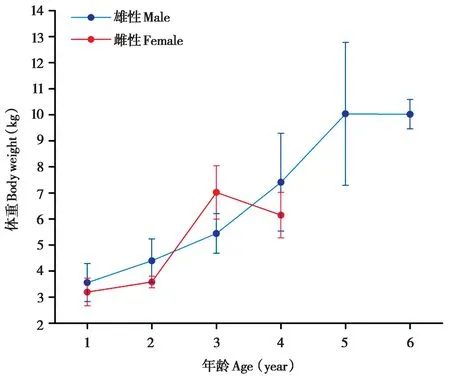
图2 未成年藏酋猴体重随年龄的变化Fig. 2 The varies of body weight with age of juvenile Tibetan macaques
成年雄性藏酋猴的体重显著高于雌性 (Z=-17.66,df= 2,P< 0.001)。进入成年期后,雌雄个体的体重均呈现进一步增加的现象,8 ~ 9岁达到稳定状态,10 岁后体重略有下降,12 岁后体重在一定范围内上下波动,但基本保持稳定 (图3)。
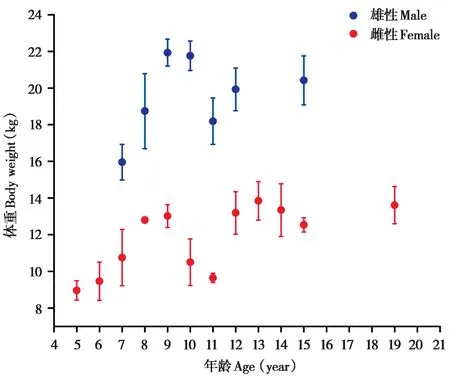
图3 不同年龄成年藏酋猴体重变化Fig.3 Age-related changes in body weight of adult Tibetan macaques
将成年藏酋猴按年龄分为年轻成年组 (雄性:7 ~ 10岁;雌性:5 ~ 10岁)、中年组 (10 ~ 15岁) 和老年组 (≥ 15岁),使用非参数检验探究藏酋猴在不同年龄组间的体重变化,结果表明,不同年龄组的成年藏酋猴体重存在显著性差异 (雄性:H=48.61,df= 2,P< 0.001;雌性:H= 69.14,df= 2,P< 0.001),雌性藏酋猴进入老年后体重显著高于中年期,但雄性藏酋猴体重在中年期和老年期无显著差异 (雌性:Z= 4.90,df= 1,P<0.001;雄性:Z= -1.03,df= 1,P= 0.303)。
2.3 藏酋猴体重与等级顺位的关系
本研究共记录到成年藏酋猴间的攻击-屈服行为805次,针对9只雄性和15只雌性成年藏酋猴的体重和DS值的Spearman相关性分析证实,成年雄性和雌性的体重都与DS 值存在显著正相关关系(雄性:R= 0.696,P< 0.001;雌性:R= 0.492,P< 0.001) (图4)。结果表明,成年雄性和雌性藏酋猴的体重均随等级顺位的提高显著增加。

图4 成年藏酋猴体重与等级顺位的关系. A:雄性;B:雌性Fig. 4 Correlation between body weight and social rank in adult Tibetan macaques. A: Male; B: Female
2.4 藏酋猴体重与季节的关系
成年雄性和雌性藏酋猴的体重数据检验结果为非正态分布。Kruskal-WallisH检验分析结果显示,雄性和雌性的平均体重均存在显著的季节差异 (雄性:H= 14.22,df= 3,P= 0.003;雌性:H= 10.32,df= 3,P= 0.016)。配对检验结果表明,雄性藏酋猴的平均体重在冬季显著低于夏季 (Z= 2.02,df= 1,P=0.043)、秋季 (Z= 3.42,df= 1,P< 0.001) 和春季(Z= -2.72,df= 1,P= 0.007);雌性藏酋猴的平均体重在秋季显著高于春季 (Z= -2.77,df= 1,P= 0.006)和冬季 (Z= -2.08,df= 1,P= 0.038);雄性和雌性藏酋猴的平均体重均在秋季达到最大值 (图5)。
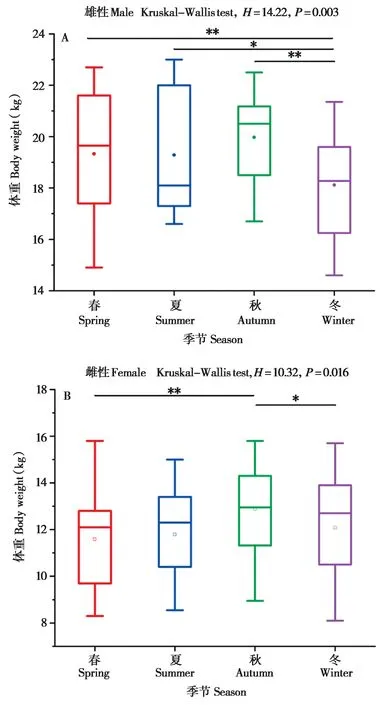
图5 成年藏酋猴体重的季节差异. A:雄性;B:雌性Fig. 5 Seasonal variation in body weight of adult Tibetan macaques.A: Male; B: Female
进一步分析了7 只怀孕与5 只未怀孕雌性平均体重在不同季节的变化,结果表明,怀孕雌性的平均体重在春季显著低于秋季 (Z= -2.03,df= 1,P= 0.042) 和冬季 (Z= -2.21,df= 1,P= 0.027) (图6A);未怀孕雌性的平均体重在不同季节无显著性差异 (P> 0.05) (图6B)。
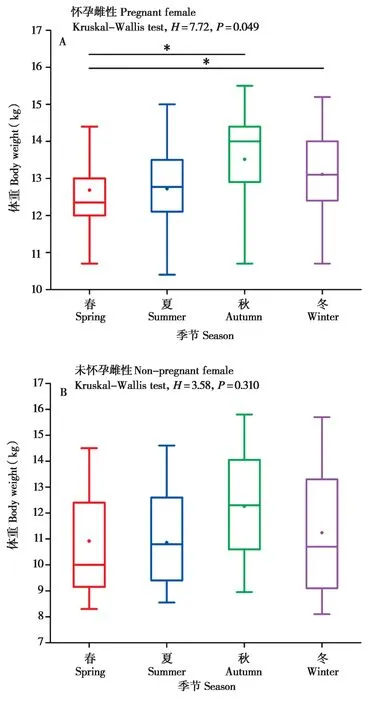
图6 雌性藏酋猴体重的季节差异. A:怀孕雌性;B:未怀孕雌性Fig. 6 Seasonal variation in body weight of female Tibetan macaques. A: Pregnant female; B: Non-pregnant female
3 讨论
3.1 藏酋猴体重与年龄的关系
本研究发现,藏酋猴体重发育为异速生长,不同生长阶段的增长速度明显不同,主要表现为先上升后下降的特点。另外,雌雄个体的快速发育期存在明显分化,雌性较雄性更早进入快速发育期。这一结果与先前对食蟹猴 (Macaca fascicularis) 的研究结论相似。未成年雄性食蟹猴的体重在4 ~ 5 岁为快速增长期,14 ~ 15 岁达到峰值;雌性个体出生后就快速增加,12 ~ 13岁达到峰值 (李冰,2006),造成这种现象的原因可能是不同发育阶段对营养的需求不同 (Setchell and Lee, 2004;Zmoraet al., 2018)。同一种灵长类动物,雄性和雌性出生时的大小和形态大致相同,但由于不同性别特有的生长策略,在发育过程中产生了分化(Badyaev, 2002)。进入青年期后,个体开始快速生长,为了获取更多的食物资源以及成年后能够更好地融入群体,个体必须拥有较大的体重和体型,才能在群体中占据优势地位。猕猴雄性体型的增加是通过延长初始生长期和提高生长速度来实现,未成年雄性较雌性拥有更长且生长速率更快的发育期,这是多夫多妻制交配系统物种的典型特征(Turcotteetal., 2022)。藏酋猴也可能存在相似的策略。多配偶制被认为反映了雄性的风险规避策略,其中延迟成熟意味着延迟雄性间竞争 (Wiley,1974; Jarman, 1983; Leigh and Terranova, 1998),有利于种群的生存和繁衍。
藏酋猴进入成年后体重仍会继续增加直至达到峰值,在中年期前后上下波动并逐渐稳定。藏酋猴在成年后为了能够更好地融入社群,短时间内体重会迅速增加以提高自身竞争力,从而获取更多的食物资源和交配机会;雄性藏酋猴体重在成年后显著高于雌性,造成这种现象的原因可能是非人灵长类动物本身具有性二型 (Plavcan,2001),雄性和雌性个体在体重的选择目的上存在一定差异。通常,雄性体重和体型较大个体会赢得战斗 (Gordon, 2006),进而具有更高的繁殖成功率,因此与较大体重相关的基因会在种群中传播开来;相比之下,雌性倾向于争夺食物等物质资源,而不是配偶 (Stercket al., 1997),因此雌性通常不会在体重上经历相同的选择目的 (Lindenfors,2002),但体重较大的雌性在争夺资源或保护后代方面往往表现更好,能够产生更大体重的后代和更好的乳汁。
3.2 藏酋猴体重与社会等级的关系
本研究发现,成年雄性和雌性藏酋猴体重均随等级顺位的升高显著增加,这一结果与猕猴以及一些原猴类动物类似。雄性个体通过打斗获取交配优先权和高质量食物资源,体重大、身型威猛的个体有更大的获胜机会,进而占据更高的等级顺位,获取更多的食物资源和交配机会(Highametal., 2011)。已有研究表明,不仅成年雄性猕猴的体重与等级顺位正相关 (Tokuda and Jensen, 1969),成年雌性的体重、头长、顶臀长也随社会等级序位增加而显著增大 (张茹帆等,2021)。一些狐猴科 (Lemuridae) (Marolfetal., 2007) 和倭狐猴属 (Microcebus) (Pochronet al., 2003) 的原猴类也表现出不同等级的雌性社会支配地位,其中,拥有较大体重的雌性控制着社会准入和资源分配(Kamilar and Cooper, 2013)。雌性优势等级可以改变母亲对后代的投资,进而影响自身以及子代的体重。例如在猕猴中,顺位较高的雌性,其后代拥有更大的体重 (Bowman and Lee, 1995)。雌性山魈(Mandrillus sphinx) 的等级顺位与其幼崽的体重呈显著正相关关系,且在断奶后,母亲等级对幼崽体重仍有影响 (Setchellet al., 2001)。灵长类成年后,雌雄个体会经历不同的自然选择 (Bercovitchet al., 2003),雌性个体体重增加可能是为了更好地抚育后代,为后代提供充足的营养和良好的生活环境,从而为顺利融入群体生活做好准备 (罗芳等,2011;刘峻杉等,2021)。
3.3 藏酋猴体重与季节的关系
成年雄性藏酋猴体重存在显著的季节波动,秋季最高,冬季最低,这一结果与峨眉山藏酋猴猴群的研究结果相同 (Zhao, 1994)。针对野生灵长类动物的长期监测结果表明,许多物种在不同季节会出现明显的体重波动。例如婆罗洲猩猩 (Pongo pygmaeus) 在食物资源丰富的季节体重会显著上升,在食物供应不足时由于无法获取足够的营养导致体重下降 (Vogelet al., 2011)。在黄山地区,藏酋猴的食物资源丰富度存在明显的季节变异,秋季食物资源最为丰富,冬季可取食的食物资源最少 (刘超等,2021);同时,由于冬季气温较低,藏酋猴消耗脂肪用以抵抗严寒,体重逐渐下降,因而雄性藏酋猴体重在秋季最高,冬季最低;随着翌年春季食物增多,气温回升,体重逐步恢复到稳定水平。但是,本研究在成年雌性藏酋猴中发现了不同的规律,雌性体重从秋季到翌年春季持续降低,造成这一结果的原因可能是繁殖季节的影响。在繁殖季节,雌性狐猴会消耗大量能量用于繁殖活动 (Brockman and Whitten, 1996)。同样,藏酋猴在繁殖季节可能会将能量用于繁殖活动而不是体重的维持,最终导致体重下降。在成年雌性藏酋猴中,怀孕个体的平均体重在冬季和春季间具有显著性差异,而未怀孕个体无显著性差异,这是由于怀孕个体除了消耗能量用于繁殖活动外,还需要消耗能量哺育后代。这与川金丝猴类似,在婴猴生活史早期,母亲对子代的照料投资维持在较高水平 (刘若爽等,2016)。另外,藏酋猴婴猴通常在春季出生,产仔后的成年雌性也在很大程度上降低自身的体重。上述因素可能是雌性藏酋猴在春季体重低于冬季的主要原因。
研究灵长类动物体重变化及其影响因素有利于深入理解灵长类发育、健康和生态适应性等科学问题,从而对野生灵长类保护提供支持。相比于实验室和动物园环境,本研究结果更能够反映自然状态下藏酋猴的体重变化,为后续研究提供基础数据。但是,由于婴猴大部分时间由母亲携带,独立活动时间相对较少,以及野外环境的不可控和采样难度的原因,位于社会边缘的个体和部分婴猴的体重难以测量,无法全面反映藏酋猴的体重变化,使本研究仍存在一定的局限性,有待进一步完善。
致谢:感谢黄山风景区管理委员会对野外采样工作给予的帮助,感谢本研究团队 (安徽大学行为生态学研究团队) 的每位成员对本研究的支持,感谢房东程海滨一家人和黄山风景区管委会谢升飞在野外生活中的照顾和关心。
猜你喜欢
科学与社会(2021年4期)2022-01-19
世界科学技术-中医药现代化(2021年5期)2021-11-05
中国房地产·综合版(2019年9期)2019-10-18
现代园艺(2018年2期)2018-03-15
森林与人类(2016年7期)2016-08-11
小天使·三年级语数英综合(2016年9期)2016-05-14
湖南中医药大学学报(2015年1期)2016-01-06
Coco薇(2015年11期)2015-11-09
世界海运(2015年8期)2015-03-11
中国医学科学院学报(2013年6期)2013-03-11
