
结核分枝杆菌潜伏感染抗原Rv2628c-Rv1737c融合蛋白的原核表达与免疫原性分析
2024-02-23 01:56解建辉孙卫国张灵霞
中国人兽共患病学报 2024年1期
解建辉,李 昆,孙卫国,贺 雄,朱 琰,张灵霞
结核病(tuberculosis, TB)是由结核分枝杆菌(Mycobacteriumtuberculosis,MTB)感染引起的一种慢性传染性疾病,我国每年约3.7万人死于结核病,是结核病高负担国家之一,其中30 %的病例由于缺乏高灵敏度和特异度的诊断方法而无法确诊,没有得到适当的诊断和治疗[1]。目前全世界将近有三分之一的人口可能感染有MTB[2]。结核病的发生、发展进程主要是由 MTB与机体免疫系统之间的相互作用所决定的,当MTB 逃避人体的免疫攻击或者与人体免疫处于一种动态平衡后,可在体内处于持续感染或呈潜伏性感染(latent tuberculosis infection, LTBI)状态,早期识别和预防结核潜伏感染成为结核病控制的一个重要组成部分[3]。MTB 潜伏感染定义为机体对MTB抗原刺激产生的持久性的反应,临床上没有任何活动性结核病征象[4]。研究发现,在LTBI者中,存在可能有5%~10%会因为免疫力低下、并发感染等原因发展为活动性结核病[5]。LTBI没有特异性的临床症状和实验室诊断指标,目前尚缺乏统一的方法和金标准对其进行诊断;同时,针对LTBI还没有有效的保护性疫苗阻止LTBI发展为活动期结核病。
在结核病的发病和发展过程中,人体的机体免疫功能发挥了关键作用[6], 在机体淋巴细胞的免疫应答过程中,T 细胞接触结核分枝杆菌的特异性抗原后成为记忆性T细胞,当再次接触MTB的特异性抗原时,T细胞将分化成为效应T细胞的同时释放多种细胞因子,包括变化水平高的γ-干扰素(interferon-γ,IFN-γ)。因此,可以通过检测INF-γ 含量变化或者释放INF-γ 的 T 淋巴细胞的数量来判断生物个体是否感染MTB,MTB 基因组发现约4 000个开放阅读框,编码几千种蛋白分子,选择什么结核分枝杆菌刺激抗原就变得尤为重要,同时需要保证检测抗原的敏感性和特异性。当结核分枝杆菌进入休眠期时,一些与潜伏感染有关的基因包括Dos R(dormancy survival regulon)调节子进行表达[7],这些MTB潜伏感染相关抗原帮助结核杆菌对抗宿主免疫活性细胞,在LTBI人群中,这类潜伏抗原激发机体产生免疫反应。Rv2628c、 Rv1737c是受 Dos R调控的潜伏感染相关抗原,研究发现Rv2628c、 Rv1737c 能诱导潜伏感染者产生较强的 IFN-γ 反应[8-9]。本实验室前期以Rv2628c 抗原刺激样本外周血单核细胞(peripheral blood mononuclear cells, PBMCs)也发现,Rv2628c 抗原在 LTBI 患者组中引起的IFN-γ分泌水平高于TB 患者组以及健康人群组[10]。同时,在BCG菌体内Rv1737c基因表达减弱,但在休眠期的结核杆菌内正常表达[11]。本研究根据结核分枝杆菌抗原刺激TB感染人群释放INF-γ 原理,以ESAT-6/CFP10和MTB全菌裂解液蛋白为对照抗原,依据Rv2628c、 Rv1737c 抗原刺激特点,原核重组获得Rv2628c-Rv1737c融合蛋白,验证其可否特异性诱导MTB感染者PBMCs细胞产生高水平的IFN-γ,根据LTBI者中诱导的特异性IFN-γ水平显著高于活动期感染者的特性,以期区分活动期病人和LTBI 者;同时证实Rv2628c-Rv1737c可被MTB 感染者尤其是LTBI 者外周血T细胞特异性识别,从而进一步研究用于TB融合蛋白疫苗构建的靶抗原可行性。
1 对象与方法
1.1 研究对象 收集解放军总医院第八医学中心结核病医学部2021年住院的30例活动期肺结核病人,男性15例、女性15例,年龄32~52岁,平均40岁。排除糖尿病、高血压等基础疾病以及其他合并症的临床期,所有结核病患者均进行胸部X 线检查,发现有浸润性病变,符合活动性结核病(active tuberculosis, ATB)诊断标准[12]。潜伏感染者样本选自总医院第八医学中心体检中心,无发热、咳嗽和咳痰等症状,影像学检查肺部无病灶,通过干扰素释放实验判定为潜伏感染者,共 20 例,男女各10例,年龄34~48岁,平均39岁。同时,选择体检中心健康体检者20例,男女各10例,年龄26~40岁,平均34岁,常规体检项目和胸部X线检查均显示正常。本研究通过单位伦理委员会审批,所有受试者均已签署知情同意书。
1.2 实验动物 实验动物SPF级雌性BALB/c小鼠30只,6~7周龄,体重16~18 g,从中国疾病预防控制中心实验动物中心购进。动物实验经单位伦理委员会审查通过。
1.3 试剂 大肠杆菌(Escherichiacoli,E.coli)DH5α、BL21(DE3)菌株和载体pET24a本实验室保存;全基因合成与测序由北京华大生物公司完成;限制性内切酶、T4连接酶、IPTG购自天根生化科技(北京)有限公司;淋巴细胞分离液Ficoll、Ni-NTA蛋白纯化树脂购自美国GE公司,本室灌装柱;TB感染T细胞检测试剂盒(TB-IGRA)购自万泰生物药业股份有限公司;蛋白质相对分子质量marker (美国sigma公司);RPMI-1640培养基和胎牛血清购于以色列BI公司;RIPA细胞裂解液购于美国Sigma公司;羊抗人IgG、酶标二抗、BCA蛋白定量试剂盒和牛血清白蛋白(BSA)购于北京索莱宝公司;SYBR荧光定量试剂盒和反转录试剂盒均购自TaKaRa公司;其它试剂为进口或国产分析纯。
1.4 MTB抗原准备
1.4.1 Rv2628c-Rv1737c蛋白原核重组与纯化 根据GenBank中公布的Rv2628c(Np-217144.1)和Rv1737c(Np-216253.1)蛋白氨基酸序列,参照E.coli体内表达密码子喜好特点,采用Rv2628c-Linker-Rv1737c融合模式对所需融合蛋白对应核酸密码子序列进行全基因合成,表达载体为pET24a,限制性酶切位点为NdeI和XhoI,柔性Linker氨基酸序列为GGGGSGGGGS。全基因核酸序列由北京华大基因公司合成,构建亚克隆表达载体pET24a-Rv2628c-Rv1737c,通过热激法转化E.coliBL21(DE3),融合蛋白的C端自带有6×His标签,通过柱上复性和亲和纯化对表达的重组蛋白进行纯化。
1.4.2 MTB H37Rv全菌裂解液的制备保存 方法为将MTB临床标准株H37Rv(ATCC 25618) 复苏后用7H9培养基于P2实验室37 ℃条件下进行液体培养3周,4 ℃条件下,5 000 r/min离心收集沉淀菌体,65 ℃条件下水浴30 min,再用无菌PBS进行充分重悬,冰浴条件下进行超声破碎,12 000 r/min离心收集上清,为H37Rv全菌裂解液,BCA定量分装后-80 ℃冰箱保存。
1.4.3 ESAT-6/CFP10多肽合成 由于无法购得标准品,同时前期实验发现以重组ESAT-6-CFP10融合肽作为抗原刺激时产生较多的非特异性,本研究MTB抗原ESAT-6和CFP10多肽由北京赛百盛公司合成,其中每条多肽长度为15个氨基酸,相邻片段重复5个氨基酸,共34条多肽[13],以PBS溶解保存。
1.5 人外周血单个核细胞的分离与刺激 采集各研究样本抗凝血2 mL,按照等比比例以RPMI 1640基础培养基稀释,然后使用等比Ficoll密度梯度离心(室温1 000×g,20 min),吸取中间细胞层获取PBMCs,再加入10 mL RPMI 1640不完全培养液充分洗涤,离心(室温400×g,5 min),弃上清,沉淀即为PBMCs。抗原刺激时,按照1×106/mL接种PBMCs。实验抗原刺激浓度分别为:Rv2628c-Rv1737c融合蛋白浓度为5 μg/mL;全菌裂解液(lysate)浓度为8 μg/mL;ESAT-6/CFP10多肽(pool)浓度为34 μg/mL(每条多肽浓度为1 μg/mL),刺激20 h后,收集细胞,加入Triozl,充分震荡,使细胞溶解于Triozl,放置-80 ℃保存,实验方法参照文献[13]进行。
1.6 PBMCs细胞总RNA的抽提与cDNA合成 常规乙醇沉淀法提取总RNA:Triozl细胞混合物进行室温融化,加入0.2 mL氯仿,剧烈振荡15 s,室温放置3 min;11 000 r/min、4 ℃条件下离心15 min,取上清到无RNAase EP管内,加入异丙醇0.5 mL进行上下颠倒混匀,静置15 min;11 000 r/min、4 ℃条件下离心15 min,弃上清,使用冰浴的1 mL无核酸酶水稀释的75%乙醇清洗RNA沉淀;干燥数分钟后加入无RNase水20 μL,震荡混匀;以Agilent 2100对RNA样本的浓度、纯度和完整性进行检测,-80 ℃冰箱保存。吸取样本RNA至10 μL反应体系中进行反转录成cDNA,反转录条件为:37 ℃温浴15 min复制,85 ℃条件下5 s。将反应产物按1∶10进行稀释后分装,-20 ℃保存备用。
1.7 IFN-γ mRNA的检测 登录NCBI网站,借用primer-BLAST程序设计IFN-γ的荧光定量PCR引物,合成的序列如下: 上游5′-GAGTGTGGAGACCATCAAG-3′,下游5′-TGAGTTCATGTATTGCTTTG-3′;实验以看家基因GAPDH作为内参,合成引物为: 上游5′-GCACCGTCAAGGCTGAGAAC-3′,下游5′-TGGTGAAGACGCCAGTGGA-3′。以SYBR Green I 荧光定量试剂盒检测mRNA的表达。反应体系为25 μL,包含cDNA模板2 μL,水9.5 μL,SYBR Green MIX 12.5 μL,上下游引物稀释浓度为10 μmol/L,反应体系各加入0.5 μL。PCR反应条件为: 95 ℃变性5 min; 94 ℃变性15 s,60 ℃退火与延伸30 s,共计40个循环。
1.8 实验动物分组及免疫 对30只16~18 g雌性BALB/c小鼠,随机分为PBS、BCG、佐剂组(弗氏不完全佐剂)、融合蛋白(Rv2628c-Rv1737c)+佐剂组和BCG+融合蛋白+佐剂组,每组6只,经皮下2点注射免疫。用PBS配制2 mg/mL的融合蛋白,与DMT佐剂等体积混合,乳化,制备融合蛋白+DMT佐剂混悬液。BCG组注射1次;PBS和融合蛋白+DMT佐剂组每1周注射1次,共4次;BCG+融合蛋白+DMT佐剂组在免疫BCG1周后,用融合蛋白+DMT佐剂加强免疫3次,间隔1周。各组每次注射剂量均为100 μL/只,BCG注射剂量约为1×106CFU/只。免疫后6周,将各组小鼠处死,取脾脏提取淋巴细胞,摘眶采血,收集血清。
1.9 ELISA法检测小鼠血清结核特异性抗体水平
取融合蛋白以包被缓冲液进行稀释至10 μg/mL,96孔酶标板每孔加100 μL,酶标板置湿盒中放置4 ℃过夜;取出酶标板,吸去包被液,洗板3次;每孔加入1∶100稀释的血清100 μL,37 ℃孵育2 h;洗板3次;将HRP标记的羊抗鼠IgG抗体1∶2 000稀释,每孔加入100 μL,37 ℃孵育2 h;洗板3次;加入等比混合的显色液A、B液,每孔100 μL,避光放置30 min;每孔加入终止液50 μL,以空白孔为零点,在酶标仪检测波长450 nm的吸收值。
1.10 ELISA法检测小鼠脾细胞培养上清中抗原特异性Th1型细胞因子水平 在24孔板中每孔加入2.5×106个小鼠脾细胞,RPMI 1640培养基、MTB全菌裂解液、融合蛋白分别作为阴性、阳性和实验组刺激物。阴性对照孔加入RPMI 1640完全培养基,20 μL/孔;实验孔加入20 μL融合蛋白,终浓度为10 μg/孔;阳性对照孔加20 μL MTB全菌裂解液,终浓度为10 μg/孔。置37 ℃培养48 h,400×g离心10 min,收集上清。采用鼠细胞因子ELISA检测试剂盒进行检测,具体操作按说明书进行。
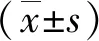
2 结 果
2.1 潜伏感染抗原Rv2628c-Rv1737c原核重组与纯化 对Rv2628c-Rv1737c蛋白密码子核酸序列根据原核系统表达特点进行优化,全基因合成后,克隆到表达载体为pET24a,测序正确,成功构建原核表达载体pET-28a-Rv2628c-Rv1737c,转化E.coli感受态细胞BL21(DE3)进行原核重组表达,以12%SDS-PAGE电泳发现重组蛋白以包涵体的形式存在(图1)。包涵体经尿素溶解后,进行柱上复性,以120 mmol/L咪唑洗涤目的蛋白,12%SDS-PAGE电泳分析纯度。结果如图2所示,纯化后的重组蛋白纯度在90%以上,分子量大约为57 kDa(Rv2628c分子量为13 kDa、Rv1737c分子量为43 kDa)同预期结果一致,BCA蛋白浓度测定融合蛋白浓度为2.5 mg/mL。

注:M.低分子量蛋白标准;1.未诱导的全菌体; 2.诱导的全菌体; 3.超声沉淀; 4.超声上清。图1 Rv2628c-Rv1737c蛋白原核表达形式SDS-PAGE分析Fig.1 Analysis of Rv2628c-Rv1737c protein expression in a prokaryotic system

注:M.低分子量蛋白标准;1.上样前溶解的包涵体; 2和3.纯化后Rv2628c-Rv1737c。图2 Rv2628c-Rv1737c 蛋白柱上复性与亲和纯化 SDS-PAGE 分析Fig.2 SDS-PAGE analysis of Rv2628c-Rv1737c protein after column renaturation and affinity purification
2.2 健康者外周血PBMCs受不同MTB抗原刺激前后IFN-γ mRNA表达差异 刺激前健康者IFN-γ mRNA表达量为3.9±1.2,经ESAT-6/CFP10(pool) 和Rv2628c-Rv1737c抗原刺激后,PBMCs中 IFN-γ mRNA表达量分别为4.0±1.3(t=1.44,P=0.16)和3.9±1.3(t=0.008,P=0.99),刺激前后差异无统计学意义(P>0.05);但健康者PBMCs经MTB全菌裂解液混合物刺激后,PBMCs中 IFN-γ mRNA表达量出现明显上升,由刺激前的3.9±1.2上升到刺激后的14.7±3.5(t=17.8,P<0.001)。见图3。

图3 健康者PBMCs 经不同 MTB 抗原刺激前后IFN-γ mRNA表达差异Fig.3 IFN-γ mRNA expression differences in PBMCs from healthy individuals after stimulation with diverse MTB antigens
2.3 ATB患者PBMCs 受不同MTB抗原刺激IFN-γ mRNA表达差异 活动期结核病患者PBMCs经不同的MTB抗原刺激后,IFN-γ mRNA的表达量均出现明显变化,其中经ESAT-6/CFP10(pool) 抗原多肽刺激后,PBMCs中 IFN-γ mRNA表达量上升为16.7±5.3;经潜伏感染相关抗原Rv2628c-Rv1737c刺激后,PBMCs中 IFN-γ mRNA表达量上升为128±18.9;经MTB全菌裂解液刺激后,PBMCs中 IFN-γ mRNA表达量上升为217±34.1。ATB患者PBMCs 经不同抗原刺激,IFN-γ mRNA 的表达量差异有统计学意义(F=175.1,P<0.001)。结果见图4。
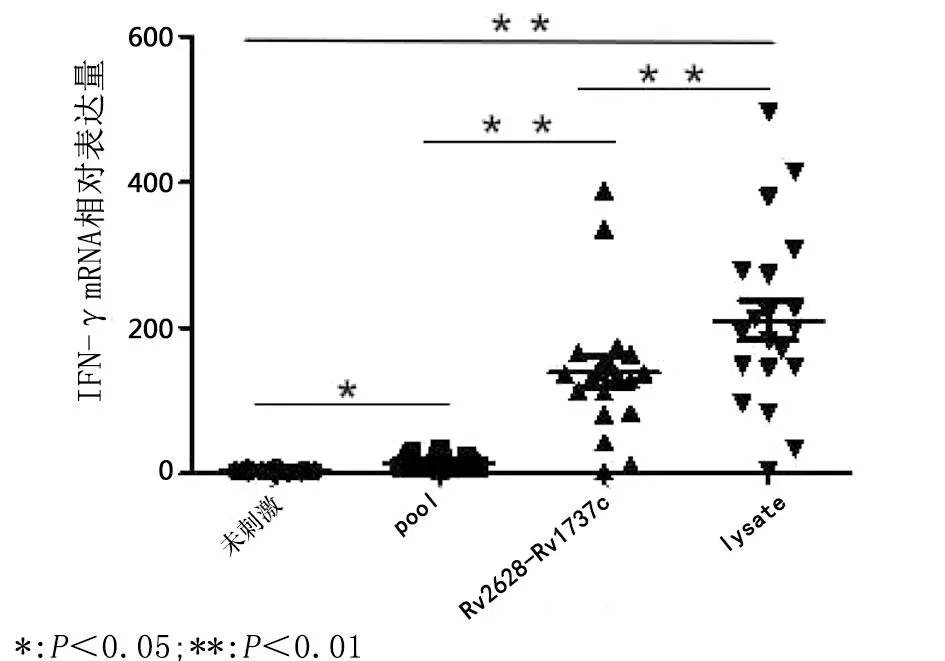
图4 ATB患者 PBMCs 经不同 MTB 抗原刺激前后IFN-γ mRNA 表达差异Fig.4 IFN-γ mRNA expression differences in PBMCs in patients with ATB after stimulation with diverse MTB antigens
2.4 LTBI者PBMCs 受不同MTB抗原刺激IFN-γ mRNA表达差异 与活动期结核病患者PBMCs相似,潜伏感染结核病患者PBMCs 经不同的MTB抗原刺激后,IFN-γ mRNA的表达量均出现明显变化,其中,经ESAT-6/CFP10(pool)抗原多肽刺激后PBMCs中 IFN-γ mRNA表达量上升至14.7±4.9;经潜伏感染相关抗原Rv2628c-Rv1737c刺激后,PBMCs中 IFN-γ mRNA表达量上升至150±28.3;经MTB全菌裂解液刺激后,PBMCs中 IFN-γ mRNA表达量上升至198±31.6。LTBI者PBMCs 经不同抗原刺激,IFN-γ mRNA 的表达量差异有统计学意义(F=369.9,P<0.001)。结果见图5。
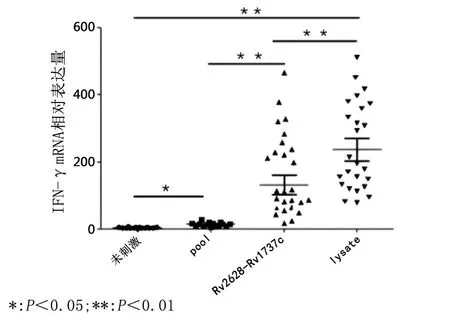
图5 LTBI者PBMCs 经不同MTB抗原刺激前后IFN-γ mRNA表达差异Fig.5 IFN-γ mRNA expression differences in PBMCs in patients with LTBI after stimulation with diverse MTB antigens
2.5 Rv2628c-Rv1737c刺激不同人群血清PBMCs后IFN-γ mRNA表达差异 与刺激前相比,ATB和LTBI者的PBMCs在经过Rv2628c-Rv1737c刺激后,IFN-γ mRNA的表达量均出现显著升高,分别上升到128±18.9(t=28.17,P<0.001)和150±28.3(t=26.11,P<0.001),3组样本经Rv2628c-Rv1737c 刺激后, IFN-γ mRNA 的表达量差异有统计学意义(F=486.2,P<0.001)。见图6。
图6 Rv2628c-Rv1737c刺激不同人群PBMCs IFN-γ mRNA表达差异Fig.6 IFN-γ mRNA expression differences in PBMCs in different populations after stimulation with Rv2628c-Rv1737c antigens
2.6 各组小鼠血清中抗原特异性IgG抗体水平 免疫后6周,处死小鼠,用MTB全菌裂解液和 融合蛋白分别检测各小鼠血清抗体水平。PBS组小鼠用MTB全菌裂解液和融合蛋白检测,均无明显特异性 IgG 抗体产生;用融合蛋白检测,BCG组抗体水平与PBS组差异无统计学意义(t=1.23,P>0.05),融合蛋白+DMT佐剂组和BCG+融合蛋白+DMT佐剂组抗体水平较PBS组显著升高(t融合蛋白+佐剂=15.92,tBCG+融合蛋白+佐剂=16.02,均P<0.01)。用MTB全菌裂解液检测时,BCG组、融合蛋白+DMT佐剂组和BCG+融合蛋白+DMT佐剂组与PBS组比较,抗体水平均有显著提升(tBCG=12.5,t融合蛋白+佐剂=6.6,tBCG+融合蛋白+佐剂=16.9,均P<0.01),结果如图7所示。

图7 Rv2628c-Rv1737c免疫小鼠产生IgG抗体值分析Fig.7 Analysis of IgG antibody levels in mice immunized with Rv2628c-Rv1737c
2.7 各组小鼠脾细胞特异性Th1型细胞因子水平 处死小鼠后分离、培养脾淋巴细胞,用MTB全菌裂解液和融合蛋白刺激时,PBS组均分泌最低水平的IL-2、IFN-γ 和 TNF-α;佐剂组与PBS组分泌IL-2(t=2.02,P>0.05)、IFN-γ(t=1.16,P>0.05)、TNF-α(t=1.47,P>0.05)无差异,其他各组分泌 IL-2(tBCG=5.8,t融合蛋白+佐剂=30.5,tBCG+融合蛋白+佐剂=42.5)、IFN-γ(tBCG=30,t融合蛋白+佐剂=30.5,tBCG+融合蛋白+佐剂=42.5)和TNF-α(tBCG=8.8,t融合蛋白+佐剂=12,tBCG+融合蛋白+佐剂=39)与PBS组差异均有统计学意义(均P<0.05)。融合蛋白刺激时,BCG组分泌各细胞因子水平均低于融合蛋白+佐剂组(tIL-2=3.56,tIFN-γ=17.4,tTNF-α=6.67,均P<0.05)和BCG+融合蛋白+佐剂组(tIL-2=4.38,tIFN-γ=25.6,tTNF-α=15.0,均P<0.05)。MTB全菌裂解液刺激时,融合蛋白+佐剂组分泌各细胞因子水平均低于BCG组(tIL-2=2.9,tIFN-γ=6.8,tTNF-α=12.5,均P<0.05)和BCG+融合蛋白+佐剂组(tIL-2=5.6,tIFN-γ=11.0,tTNF-α=4.0,均P<0.05)。实验结果见图8。

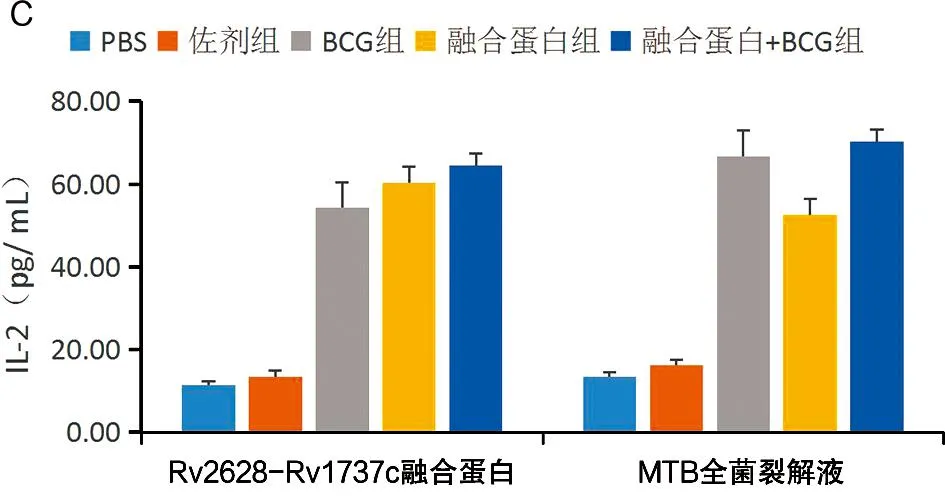
图8 各组小鼠脾细胞分泌抗原特异性IFN-γ(A)、TNF-α(B)和IL-2(C)水平Fig.8 Antigen specific IFN-γ(A),TNF-α(B), and IL-2(C) levels secreted by spleen cells from mice in each group
3 讨 论
LTBI无特异性的症状和体征指标,实验室诊断缺乏统一的方法和金标准。虽然LTBI者无传染性,但当机体因为消瘦、疲劳和其他并发症导致免疫力降低时,会导致 MTB大量增殖,发展为活动性结核,严重情况下引发全身播散性结核,如消化道结核、骨结核,尤其是引起结核性脑膜炎[14-15]。寻找快速、有效的诊断方法鉴别LTBI者并对其进行预防性治疗,是减少活动性结核发生的有效途径。IFN-γ 是机体对抗结核分枝杆菌感染的一类重要的Th1型细胞因子,主要由活化的T细胞和NK细胞产生[16],IFN-γ 对阻碍和杀灭机体胞内 MTB有重要作用。研究发现,IFN-γ 受体缺乏则可能使人群对TB 的易感性升高[17]。结核病患者在不同感染状态具有特异性的抗原特征谱,通过免疫分析可以区分不同感染状态[18]。本项研究中,选择 Rv2628c-Rv1737c融合蛋白作为 TB患者靶抗原,通过刺激不同样本人群PBMC细胞IFN-γ mRNA水平的检测,发现相对于刺激前,其可特异性诱导 MTB 感染者PBMC细胞产生高水平的 IFN-γ,尤其在LTBI者中诱导的特异性 IFN-γ水平显著高于ATB 感染者。实验发现,Rv2628c-Rv1737c相对于ESAT-6/CFP10抗原多肽具有较高的敏感性,对MTB全菌裂解液而言又具有较强的特异性,同时ATB和LTBI人群PBMCs受Rv2628c-Rv1737c融合蛋白刺激时,IFN-γ mRNA表达均出现上调,分别上升到128±18.9和150±28.3,利用Rv2628c-Rv1737c抗原刺激IFN-γ 表达差异特性以区分TB患者临床活动性期与潜伏感染期。本实验中出现样本受Rv2628c-Rv1737c抗原刺激后出现假阴性现象,存在原因可能是受试者因为自身免疫力低下的原因造成,同时样本的受融合抗原刺激后,阳性样本的IFN-γ 表达出现明显的个体差异,这需要在后续实验中进行条件优化,提高检测的平行性。
蛋白亚单位疫苗是研究新型有潜力的结核疫苗之一,筛选出能够被MTB感染者T 细胞免疫识别的潜力靶抗原,是构建更强保护力的新型TB 亚单位疫苗的保证[19]。目前多数进入临床试验的TB蛋白疫苗保护性有限。MTB感染者中含有大量LTBI人群,对这类人群进行预防保护,阻止发展成为ATB患者,是2035年消灭结核病的关键一环。目前认为结核抗原诱导Th1 型免疫应答水平(尤其是抗原特异性IFN-γ水平)与抗TB 保护性密切相关[20]。诱导效应性和记忆性CD4+和CD8+T细胞产生高水平 IFN-γ和 IFN-γ/IL-2 多功能应答效应,成为评价候选疫苗靶抗原免疫原性的重要依据[21]。Arroyo等[22]比较Dos R、重组融合蛋白ESAT-6-CFP10(E6-C10)和结核菌素(PPD)抗原刺激潜伏感染人群和活动性结核患者的外周血单核淋巴细胞,检测细胞上清液 IFN-γ 水平,认为 pfkB 是区分LTBI和活动性感染组的最优单一抗原,作为Dos R调控蛋白,Rv2628c、Rv1737c均属于潜伏感染相关抗原,本实验鉴于IFN-γ释放试验和潜伏感染抗原的免疫特点,通过原核系统重组、表达和纯化获得Rv2628c-Rv1737c融合蛋白,去刺激ATB和LTBI人群均检测到IFN-γ mRNA表达水平在两者具有差异和较强特异性,在鉴别ATB和LTBI人群上具有潜在应用价值。另外实验小鼠模型中,Rv2628c-Rv1737c/DMT和BCG+Rv2628c-Rv1737c/ DMT免疫小鼠均可诱导出IFN-γ、TNF-α和IL-2因子的高水平分泌,以及产生特异性 Ig G。TNF-α 参与了肉芽肿的形成,其协同 IFN-γ共同诱导单核细胞和粒细胞向感染病灶的迁移,控制MTB感染[23],因此重组Rv2628c-Rv1737c/DMT 诱导小鼠产生以Th1型细胞反应为主的免疫应答。实验数据证实,重组Rv2628c-Rv1737c/DMT的免疫效果低于BCG组,这也佐证BCG作为减毒活菌疫苗,能在较长时间内持续刺激机体免疫反应,引起的免疫反应比单一抗原或组合抗原引起的免疫反应要强,而BCG+Rv2628c-Rv1737c/DMT组比Rv2628c-Rv1737c/DMT组及BCG的免疫效果都要强,说明Rv2628c-Rv1737c组合蛋白能增强BCG的免疫效果,可以用于构建重组卡介苗,或者用于卡介苗接种后的加强接种。
本研究获得的Rv2628c-Rv1737c蛋白能刺激TB患者尤其是 LTBI 者外周血 T 细胞特异性识别,诱导小鼠产生以Th1 型细胞反应为主的免疫应答,而且可以增强卡介苗的免疫应答,因此可作为靶抗原用于构建融合多肽抗原的TB 增强保护型亚单位疫苗及重组卡介苗。
利益冲突:无
猜你喜欢
保健医苑(2022年5期)2022-06-10
医学概论(2022年4期)2022-04-24
昆明医科大学学报(2021年3期)2021-07-22
中成药(2018年10期)2018-10-26
特别健康(2018年4期)2018-07-03
中国医学装备(2016年6期)2016-12-01
湖南中医药大学学报(2016年1期)2016-12-01
现代畜牧兽医(2016年11期)2016-02-21
中国卫生(2015年1期)2015-11-16
中国当代医药(2015年36期)2015-03-11
