
江苏省某监测点家畜来源肠致病性大肠埃希菌分离株分子特征分析
2024-02-23 01:57:02孔筱筱程晓庆
中国人兽共患病学报 2024年1期
孔筱筱,韩 悦,周 璐,程晓庆,董 晨
肠致病性大肠埃希菌(enteropathogenicEscherichiacoli,EPEC)是导致发展中国家婴儿腹泻的一个重要原因[1],已从动物和动物食品中分离出来。将携带编码紧密黏附素的eae基因,而不具有编码志贺毒素的stx基因的致泻性大肠埃希菌(diarrheagenicE.coli,DEC)定义为EPEC[2]。我国由DEC引起的5岁以下腹泻儿童病例中,EPEC是常见病原体[3]。
动物一直被认为是EPEC的天然宿主[4]。EPEC可被动物无症状携带,通过粪便污染周围环境,并在屠宰和加工过程中污染肉类,从而导致人类感染[1]。江苏省在畜牧养殖业方面有着传统优势,家畜粪便携带的EPEC对于环境污染所引发的公共卫生安全问题不容忽视。本研究对某监测点家畜来源的EPEC进行全基因组测序,了解其血清型分布、毒力基因携带情况、ST型别特征以及与同期分离的产志贺毒素大肠埃希菌(shiga toxin-producingEscherichiacoli,STEC)之间的遗传进化关系,进而评估其潜在的致病性,为感染性腹泻的监测与防控提供依据。
1 材料与方法
1.1 材 料
1.1.1 菌株来源 37株EPEC分离株、68株STEC分离株均分离自2019年江苏省产志贺毒素大肠埃希菌相关监测点牛羊家畜动物的粪便中,由江苏省疾病预防控制中心急性传染病防制所采用磁珠法-80 ℃保存。菌株的复苏、纯化及鉴定方法参照文献[4]。
1.1.2 主要试剂和仪器 营养琼脂培养基(北京陆桥技术有限公司)、ECC显色培养基(法国CHROMagar公司)、细菌基因组DNA提取试剂盒(美国Promega公司)、PCR Master Mix(美国Promega公司)、PCR仪(美国Applied Biosystems公司)、毛细管电泳仪(德国Qiagen公司)、PCR引物(上海生工生物)。
1.2 方 法
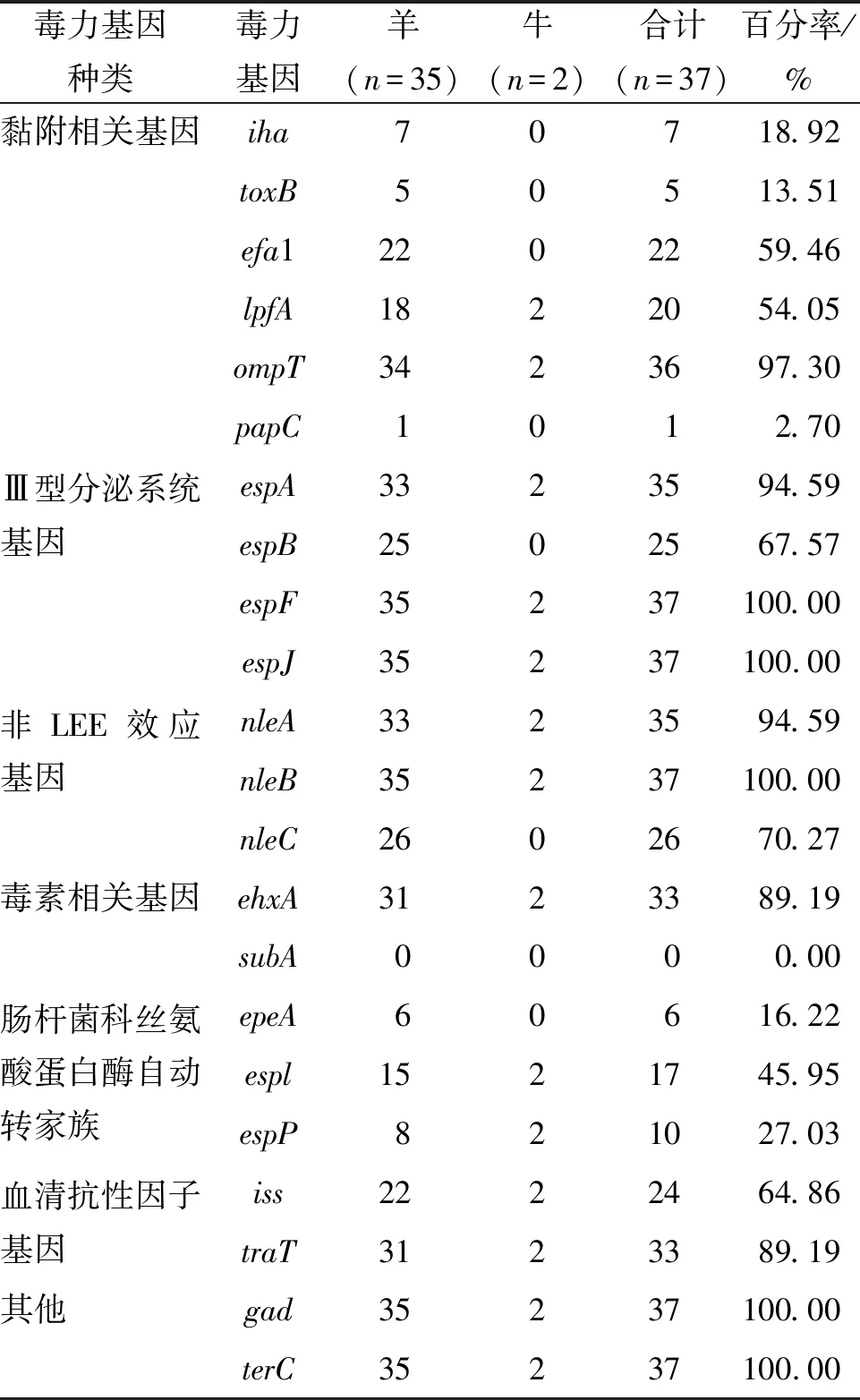
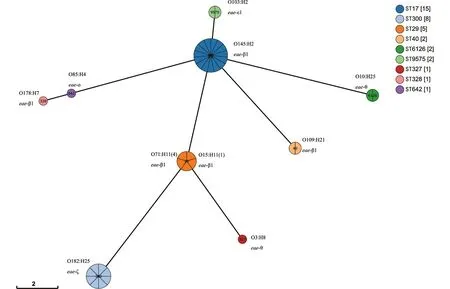
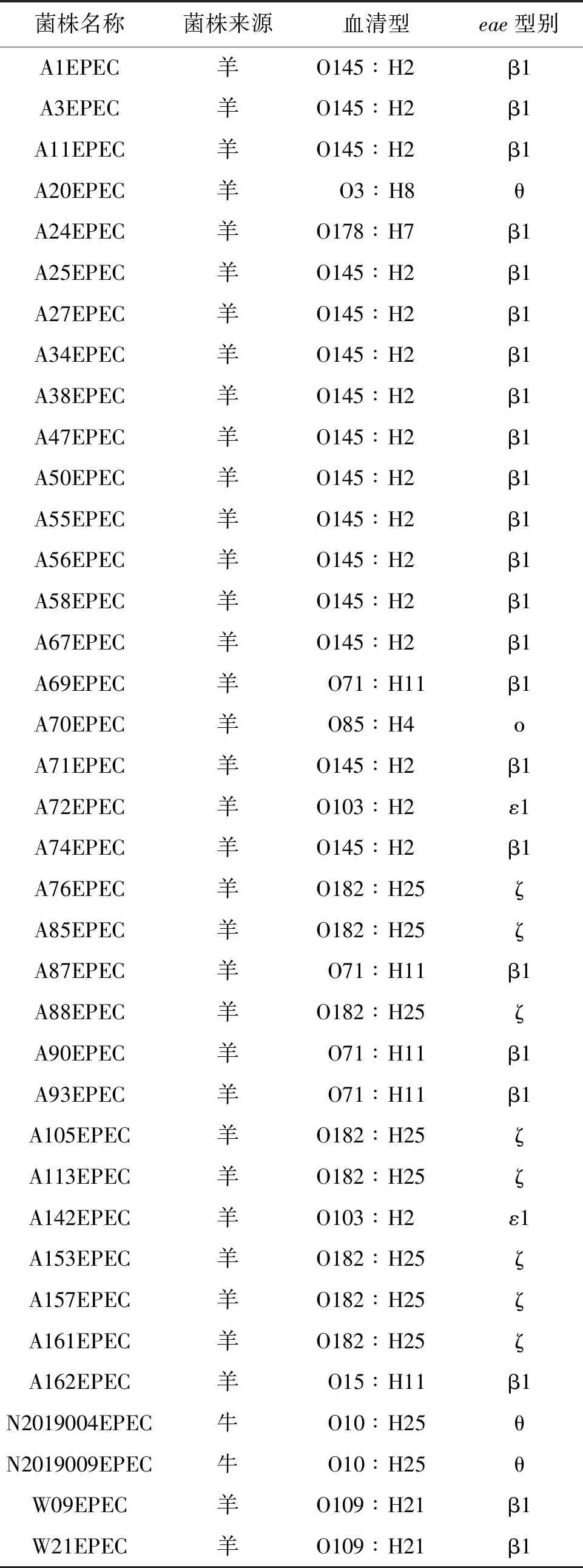
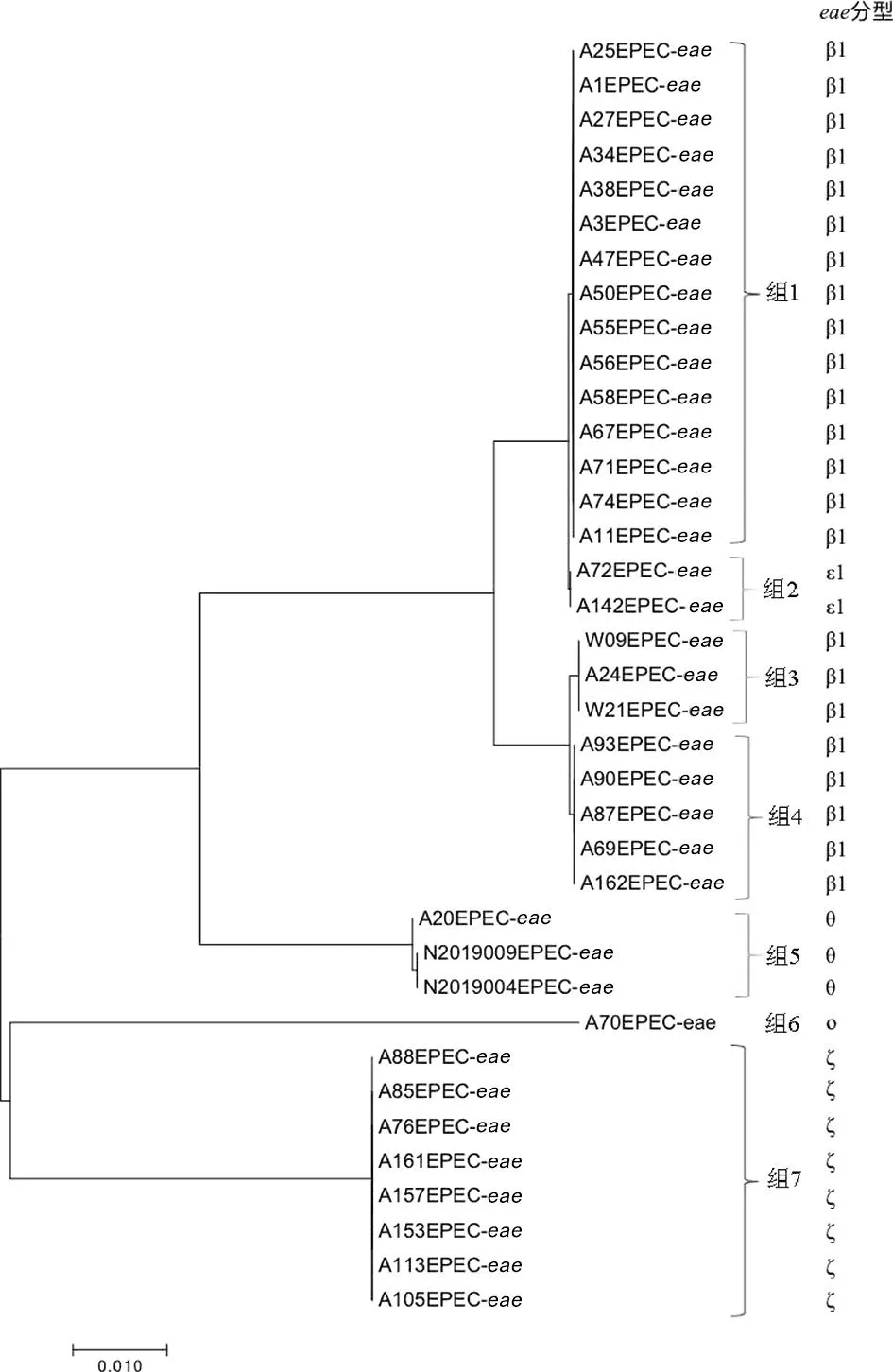
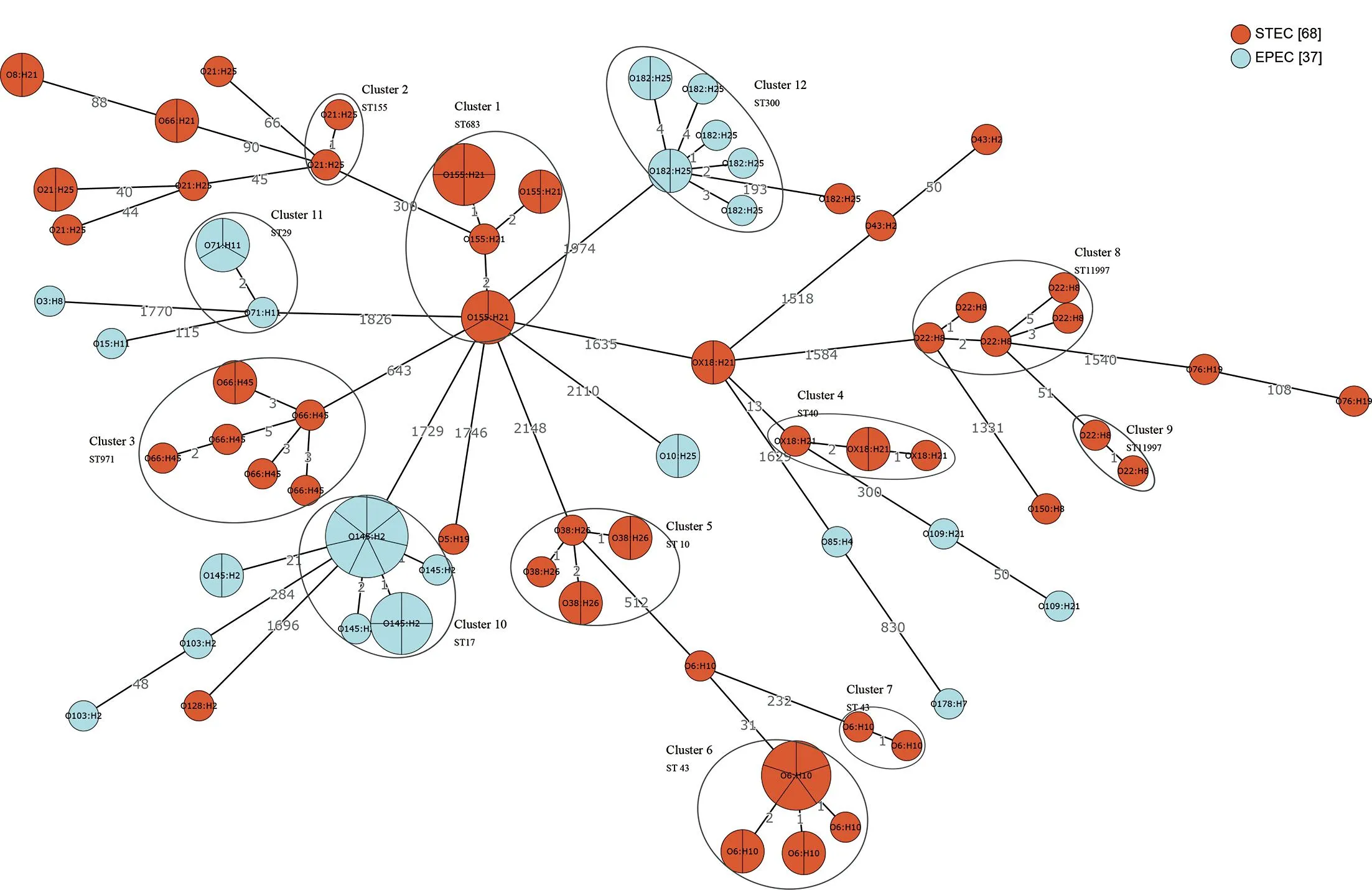
1.2.1 全基因组测序、组装 采用细菌基因组DNA提取试剂盒对测试菌株基因组进行提取。使用Illumina HiSeq PE150测序平台构建成对双末端测序文库,全基因组测序工作由上海生工生物工程有限公司完成。通过fastp 0.21.0剔除reads首尾质量 1.2.2 EPEC菌株分子特征分析 1.2.2.1 血清型 将组装完整的基因组序列与Center for Genomic Epidemiology(CGE)SerotypeFinder v2.0数据库[5](https://cge.cbs.dtu.dk/services/SerotypeFinder/)进行比对,设置比对参数为:序列相似性≥90%,覆盖度≥95%,获得菌株O∶H血清型,如果基因组血清型未预测成功,再使用EnteroBase数据库进行血清型预测。 1.2.2.2bfpA基因、毒力基因分析 将组装完整的基因组序列与CGE VirulenceFinder v2.0数据库[6](https://cge.cbs.dtu.dk/services/Virulence-Finder/)进行比对,识别bfpA及其他毒力基因,设置比对参数为:序列相似性>90%,覆盖度≥90%。 1.2.2.3 MLST分型 使用EnteroBase数据库对菌株基因组识别基于7个管家基因(adk、icd、fumC、rgyrB、purA、mdh和recA)的MLST分型,采用MEGA v7.0.26软件构建最小生成树。 1.2.2.4eae基因分型及进化关系分析 从组装完整的基因组序列中提取eae基因的全长,从NCBI GenBank数据库中获得常见26种eae基因型别序列。使用NCBI中BLASTn套件[7](https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行eae型别比对,当序列同源性大于98%时,则认为该分离株eae与参考eae型别一致。 使用MEGA v7.0.26软件中Clustal W进行多序列比对并对齐,采用最大似然法构建进化树,分析37株EPEC分离株间eae基因关系。 1.2.2.5 基于cgMLST构建EPEC与同期分离的STEC系统发育树 本研究利用Enterobase数据库进行cgMLST分型来评估37株EPEC与当地同期分离的68株STEC菌株之间的进化关系。cgMLST核心基因组方案采用Enterobase数据库中大肠杆菌2 513个核心基因组等位基因位点方案[8]。通过GrapeTree v. 2.2[9]程序使用MSTree v2算法构建最小生成树(minimum spanning tree,MST),同时将簇定义为小于10个等位基因差异的菌株群。 2.1 血清型、毒力基因及ST型 37株EPEC分离株共分为10种不同O血清群和7种不同H型,组成10种不同O∶H血清型。从羊粪中分离的EPEC的优势血清型为O145∶H2 (40.5%, 15/37),O182∶H25 (21.6%, 8/37),和O71∶H11 (10.8%, 4/37)(表2)。2株来源于牛粪的EPEC血清型同属于O10∶H25。 37株EPEC中均为bfpA基因阴性,判定为非典型EPEC(atypical EPEC, aEPEC)。其他毒力基因分布情况详见表1。 表1 EPEC分离株其他毒力基因分布Tab.1 Distribution of other virulence genes in EPEC isolates 根据MLST分型方法,37株分离株被分成9种ST型(图1),其中ST17和ST300比例较高,分别为40.5%(15/37)和21.6%(8/37)。除ST29 包含有2种血清型(O71∶H11,O151∶H11)外,其余ST型对应单一的血清型。 图1 EPEC菌株ST型别构建的最小生成树Fig.1 Minimum spanning tree constructed on the basis of EPEC strain ST type 2.2eae基因分型及进化关系分析 37株EPEC分离株经比对后,共产生5种eae型别,分别为β1、ζ、θ、ε1和ο。其中最主要的型别为β1,占比62.2%(23/37),其次为ζ,占比21.6%(8/37),而θ、ε1和ο型分别为3株、2株和1株。见表2。 表2 37株EPEC分离株eae基因分型结果Tab.2 Genotyping results of eae in 37 EPEC isolates 35株羊来源的EPEC分离株存在5种eae型别(β1、ζ、θ、ε1和ο),而2株牛来源的EPEC分离株eae型别均为θ。见表2。 eae基因的聚类结果表明: 37株菌的eae基因共分为7个组,其中组1、组3、组4分别对应不同的β1型基因序列,组5包含2种不同的θ型eae基因序列,其余3组各对应ε1、ο、ζ型的1种基因序列。从和血清型的对应关系来看,组1为β1型,包含有15株菌,血清型为O145∶H2;组2为ε1型,包含有2株菌,血清型为O103∶H2;组3 为β1型,包含有3株菌,血清型为O178∶H7、O109∶H21;组4为β1型,包含有5株菌,血清型为O71∶H11、O15∶H11;组5为θ型,包含有3株菌,血清型为O3∶H8、O10∶H25;组6为ο型,包含1株菌,血清型为O85∶H4,组7为ζ型,包含有8株菌,血清型为O182∶H25。见表2和图2。 图2 采用最大似然法构建eae基因的系统进化树Fig.2 Phylogenetic tree of the eae gene with the maximum likelihood method 2.3 基于cgMLST构建EPEC与同期分离的STEC系统发育树 37株EPEC和68株同期分离的STEC进化关系采用Enterobase数据库中的cgMLST方案,如图3所示。所有分离株间等位基因差异为0~2 148个。当采用10个等位基因差异的聚类阈值时,EPEC 中形成了3个簇,而STEC中形成了9个簇。每个簇包含有相同的血清型和ST型,菌株间的等位基因差异为1~5个等位基因。簇间的等位基因差异数至少为51个等位基因。EPEC中的优势簇为簇10,由13株 O145∶H2 EPEC组成。在簇12中观察到eae阳性O182∶H25 STEC菌株和O182∶H25 EPEC菌株之间有193个等位基因差异。 注:圆圈内的编号代表菌株血清型,圆圈间连线上的数字代表等位基因差异数。图3 基于37株EPEC和同期分离的68株STEC核心等位基因差异构建最小生成树Fig.3 Minimum spanning tree based on core allele differences between 37 EPEC strains and 68 STEC strains isolated in the same period 过去,血清型一直做为EPEC的鉴定依据。随着高通量测序技术的发展,越来越多的EPEC的血清型被发现[10-11]。迄今为止,世界范围内已经发现200多种血清型的EPEC菌株[12]。本研究的结果显示,监测点分离的EPEC菌株的血清型呈现多样性。建议在EPEC日常检测中采用分子生物学方法。 bfpA基因是区分典型EPEC(tEPEC)和非典型EPEC(aEPEC)的主要依据[1]。本研究中,所有的EPEC均为aEPEC。这与其他文献报道的相一致:即tEPEC的宿主只有人,而aEPEC在人和动物中广泛存在[11]。近年来越来越多的研究发现,目前aEPEC在发展中国家和发达国家的存在的比例高于tEPEC[13],甚至有报道表明其存在率达100%[14]。我国的数据也表明,中国EPEC菌株中,aEPEC菌株所占比例也显著高于tEPEC[12]。尽管aEPEC较tEPEC引起的腹泻症状轻,不会造成脱水,但是其腹泻时间延长,容易引起儿童慢性腹泻、营养不良等[13]。本文中aEPEC毒力基因携带种类多样。值得关注的是,Ⅲ型分泌系统基因espB(67.57%)、espF(100%)、espJ(100%)、非LEE效应基因nleB(100%)、毒素相关基因ehxA(89.19%)都是与EPEC致病相关的重要毒力基因[13]。MLST分型结果表明,本地监测点EPEC菌株ST型丰富,且与其他地区报道的家畜来源的大肠埃希菌ST型不同[14],说明此监测点家畜来源EPEC的传播具有地区局限性的特点。 eae基因位于EPEC染色体的LEE(locus of enterocyte effacement)毒力岛中,会对肠黏膜造成黏附/抹平(attaching and effacing,A/E)损伤[15],全长约2 800 bp[4]。由于eae基因5′端相对保守,而3′端差异较大,根据这种差异,将eae分为不同亚型,目前已发现超过30种不同型别,不同eae型别具有不同的宿主特异性和组织嗜性[16]。本研究中,β1型是eae基因主要的亚型。有报道表明,含有β1 型eae基因的EPEC 菌株在腹泻病人中最为常见[4]。可见此监测点动物来源的EPEC具有一定的致病潜力,需要加以关注。eae基因的聚类结果显示,同种血清型的EPEC菌株,具有同种的eae亚型,如图2组1中,15株O145∶H2 EPEC菌株均为β1型;同时不同血清型的EPEC菌株也可能具有相同的eae亚型,如图3,虽然其中包含的菌株eae亚型序列相同,均为β1型,但是他们的血清型不同。这个结果可以推测,相同的eae亚型可以在不同血清型的菌株中转移[17]。 有研究表明,aEPEC在遗传学上和携带有eae基因的STEC更为接近。本研究采用基于全基因组测序技术的cgMLST方法来构建EPEC与同期分离STEC的系统发育树,以了解EPEC和STEC之间的遗传进化关系。结果表明,基于不同的血清型和ST型,EPEC和STEC各自成簇。其原因可能是此监测点分离到的大多数STEC不含有eae基因。而1株eae阳性O182∶H25 STEC菌株与其他STEC菌株遗传距离较远,与同种血清型的 EPEC菌株之间有193个等位基因差异。有研究假设,在感染、分离或继代培养过程中,携带eae基因的STEC菌株可以和EPEC菌株通过丢失或溶原化编码Stx的噬菌体进行转化[18]。因此,我们推测这株eae阳性O182∶H25 STEC菌株的编码Stx的噬菌体在同种血清型的EPEC中可能发生了转移。 综上所述,本研究采用全基因组测序技术对江苏省某监测点分离的家畜来源的37株EPEC菌株进行特征性分析,发现此监测点家畜来源的EPEC均为aEPEC,其血清型多样、携带多种与腹泻相关的毒力基因,ST型丰富且具有区域性特征,eae基因包含有5种亚型,有1种优势亚型在腹泻病人中最为常见,可见此监测点aEPEC对当地公众健康构成潜在威胁。此外,我们推测此监测点的aEPEC与STEC存在通过噬菌体的转移发生转化的可能性,由于STEC是一类可引起暴发和严重疾病的最常见致泻性大肠杆菌,所以我们认为不可忽视家畜传播aEPEC的潜在风险,应在感染性腹泻的监测工作中,加强对家畜粪便及养殖环境的监测。 利益冲突:无2 结 果
3 讨 论
猜你喜欢
云南化工(2021年6期)2021-12-21 07:31:04
农药科学与管理(2019年6期)2019-11-23 08:17:12
天然产物研究与开发(2018年9期)2018-10-08 03:25:52
大连海洋大学学报(2018年3期)2018-07-24 13:04:48
China Communications(2017年12期)2017-04-10 02:40:10
食品界(2016年4期)2016-02-27 07:36:44
华南农业大学学报(2015年5期)2015-12-04 03:04:38
中国兽医杂志(2015年8期)2015-08-08 10:16:06
医学综述(2014年14期)2014-03-27 10:31:04
现代检验医学杂志(2014年5期)2014-02-02 02:51:51
