
遗传漂变与甜菜育种
2024-02-20 13:22胡华兵贺碧微孙琳琳刘建雄
新疆农垦科技 2024年1期
胡华兵,贺碧微,郇 町,孙琳琳,刘建雄,刘 珣
(石河子农业科学研究院,新疆 石河子 832011)
遗传漂变也叫基因漂变,是指物种种群基因库在代际之间发生随机变化的一种现象,即在一个群体内,从基因库抽样形成下一代个体的配子时,就会产生一定的误差,这种误差会引起群体内基因频率、基因型频率的随机偶然变化[1]。因为任何个体的基因来源于亲代具有一定的随机性,所以繁殖过程可视作一种抽样,即子代携带的等位基因是对亲代抽取的一种样本。这种抽样误差使子代中的等位基因频率与亲代并不相等[1]。遗传漂变是实际抽样与真实几率之间的差异,而非自然选择因素或环境因素所造成。
作物育种过程中,遗传漂变是亲本材料性状不断变化的原生动力之一,直到达到育种目标,并达到群体遗传平衡。遗传漂变会改变群体基因频率和基因型频率,打破遗传平衡[2]。在甜菜自育品种生产实践中,常常出现复配组合产量、质量不及原配组合的情况,主要原因是复配亲本基因频率和基因型频率发生了改变,即发生了遗传漂变,导致组合种性发生了变化。甜菜不育系、保持系和授粉系性状稳定是品种种性保持稳定一致的关键所在。
本文根据群体遗传漂变规律和遗传平衡定律由浅及深揭示自育甜菜品种种性变化原因,并从理论上推导出授粉系不能是S型胞质材料,只能是N型胞质材料,为甜菜亲本选育、新品种选育繁育提供理论支撑。
1 作物的遗传漂变
1.1 不同授粉类型作物的影响
不同作物有不同的授粉方式,如大麦属于自花授粉作物,自然异交率为0.04%~0.15%[3];小麦也属于自花授粉作物,自然异交率为1%左右[4];棉花属于常异花授粉作物,陆地棉的天然异交率为1%~18%[5];甜菜、玉米等属于典型的异花授粉作物,一般异交率为50%以上,高的可达95%以上[6]。在育种工作中,不同作物极小群体在相同正常条件下,自交后代基因频率和基因型频率变化幅度是不一样的,即发生遗传漂变的效应是不一样的;异花授粉作物遗传漂变效应强、速度也快,常异花授粉作物遗传漂变效应和速度皆为中等,自花授粉作物遗传漂变效应弱、速度也慢,遗传最稳定(见图1)。

图1 小群体遗传漂变规律
1.2 群体规模的影响
群体规模是影响群体遗传漂变的重要因素,一般情况下,对于同一种作物不同数量群体发生遗传漂变的效应和速度是不一样的。群体越小发生遗传漂变速度越快,有的群体1~2代就造成某个基因的固定或另一个基因的消失,从而改变其遗传结构;群体数量越大,发生遗传漂变的效应越弱(见图2),漂变速度也越慢。遗传漂变是生物进化的关键机制之一。

图2 不同群体数量遗传漂变规律律
1.3 初始基因频率的影响
雌雄比例和基因的初始频率是影响群体遗传漂变的重要因素[7]。作物的选择育种,其实就是人为选择极小群体或单株,利用奠基者效应定向干预群体基因频率和基因型频率,使之发生遗传漂变,发生基因的固定或者丢失,会造成群体的遗传基础越来越窄,物种(群体)的遗传多样性降低。从理论上讲,群体规模越大,越有利于基因不被固定或丢失。尽量扩大保种群体规模,对于保持群体的遗传多样性、维持群体遗传结构的相对稳定以及提高保种效果都具有积极意义[8]。
2 遗传漂变与遗传平衡定律
无限大的群体在没有突变、选择、迁移、遗传漂变的条件下,随机交配,则群体内一个位点上的基因频率和基因型频率将代代保持不变,处于遗传平衡状态[9]。这就是遗传平衡定律,亦即哈迪-温伯格定律。遗传平衡是一种理想情况,在自然状态下是无法达到的,但在一个足够大的种群中,如果个体间是随机交配且没有明显的自然选择,那么这个群体可以近似地看作符合遗传平衡[10]。
如果无限大的群体内发生遗传漂变,群体的基因频率和基因型频率就会发生波动。如果群体内配子不能随机交换,子代取样误差增加,后代群体的基因频率和基因型频率会发生波动,就会发生遗传漂变,无法达到遗传平衡。在小群体里,遗传漂变现象能够表现出来,而在大群体里,遗传平衡定律对基因频率和基因型频率的影响就会表现出来。
如对于一个地区总人口(没有迁移和突变)来讲,白化病的基因频率和基因型频率是波动的。白化病是隐性基因控制,现实中白化病患者很少能结婚,而表型正常、白化病基因携带者往往能够成婚。自由婚配后,白化病患者的配子通常无法传递于子代,而婚配携带者可以正常传递配子于子代,配子交换具有选择性就发生了遗传漂变,群体中白化病基因频率和基因型频率会发生变动。可以推论:若干年后,该地区人口白化病的基因频率和基因型频率会逐渐降低,无法达到遗传平衡。相较于白化病,色盲患者和携带者往往能够正常结婚,自由婚配后,患者和携带者均可将配子随机传递于子代,可以推论:该地区人口色盲基因频率和基因型频率基本维持稳定,近似达到遗传平衡。
3 遗传漂变对甜菜育种的影响
3.1 遗传漂变与甜菜育种
在甜菜育种实践中,甜菜不育系、保持系选育需要保持株和不育株杂交后再循环回交,这种株对株极小群体杂交、循环回交就会发生明显的遗传漂变,最后选育出的姊妹不育系、保持系在表型、产质量、配合力等方面会出现明显的不同或相似之处;低代分离会出现显著性差异,高代分离就会有相似之处。自交系的选育类似于保持系的选育。不育系、保持系的提纯扩繁过程亦会导致群体基因频率和基因型频率改变,发生遗传漂变,这是复配组合种性变化的原因之一[11]。
在成系群体繁殖过程中,甜菜种子更新繁育需增加繁殖群体数量,减少繁育代数,做好田间管理[11]。图3中a和b表示甜菜群体两种繁殖利用方法,其中N表示繁育的种子总数量,X表示生产或组合配置所消耗的种子数量。从图中可以看出,b的繁育方法能很好控制群体遗传漂变的发生,保持不同年份间群体种性基本稳定一致,继而保持复配组合的种性稳定。

图3 种子繁育示意图
国外优良甜菜品种在推广应用中,往往一个品种推广十余年,期间品种的种性保持基本稳定一致,这与国外完善的良种繁育体系有密切关系。甜菜育种流程繁杂,品种作为三系三交种,品种的种性稳定依赖于两对不育系、相应保持系以及授粉系的稳定。因此,甜菜育种制种难度高于其他作物。
3.2 甜菜N型胞质授粉系利用理论推导
按照欧文假说和勃里斯假说,甜菜的育性受X和Z基因共同支配,其中,雄不育的恢复受X显性基因所支配,半不育受X基因的下位基因Z所支配[12]。S(xxzz)为全不育,S(xxzZ)、S(xxZZ)为半不育,S(Xxzz)、S(XXzz)为半可育,S(X_Z_)为可育。在一个S型胞质的群体中,各种育性的基因都可能存在,即使人为拔除全不育型株型,后代群体也还是会分离出全不育型。在一个表型相对整齐一致的S型胞质群体中,各种配子都会产生,但全不育型、半不育型、半可育性三种类型都是接受配子多,而自身不产生配子或产生少量配子。群体中配子未能自然随机交换配子,会增加子代群体等位基因取样误差,造成基因频率和基因型频率波动,即发生遗传漂变,导致种性发生改变。多代遗传漂变作用积累,势必会造成群体的种性发生较大变化。以此群体配置的组合亦会发生种性改变。一直以来,只要自交系综合性状优良,整齐度好,就可以作为父本配置组合。因此,父本的育性胞质类型有S型,或N型,或混合型,只要有花粉即可,但没有考虑未来复配组合的性状是否稳定一致。甜菜育种实践中,成系群体必须达到性状一致且遗传平衡,这样基因频率和基因型频率可以基本保持稳定,所以S型胞质的群体不适合做授粉系(理论上讲S(XXZZ)可以,但实际中几乎不可能获得纯基因型群体),只有N型胞质的材料才能选育授粉系。
欧美国家甜菜育种机构多年以前选育的品种就是三系三交杂交种,即A1/B2//B3(图4),该杂交种亦为三元不育系,B3为多粒保持系;如果A1/B2为单粒,则杂交种为单粒,如果A1/B2为多粒,则杂交种为多粒。长久以来,我国科研工作者认为该类杂交种之所以用多粒保持系B3作为父本是为了使杂交种保持不育性状,以便限制品种基因被他人利用。但其实还有另一方面,多粒保持系B3的细胞质类型为N型,该群体在代际扩繁中能保持性状稳定,达到遗传平衡,年度之间配置的同一组合能保持种性稳定一致。
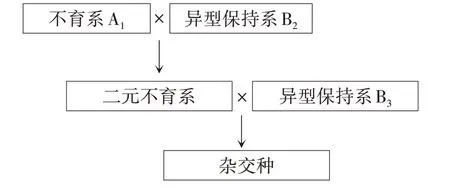
图4 国外甜菜品种配置方式
4 结论
作物群体达到遗传平衡,基因频率和基因型频率会保持稳定,这是一种理想状态。实际上,群体数量、地理环境、自然选择以及初始基因等因素皆会造成群体基因频率和基因型频率波动。因此,甜菜群体代际繁殖中种性会逐渐发生变化。
在作物育种实践中,群体数量越小遗传漂变速度越快,反之亦然。对同样数量的小群体,不同授粉类型的作物发生遗传漂变速度亦不一样,异花授粉作物>常异花授粉作物>自花授粉作物。甜菜作为异花授粉作物,不育系、保持系、自交系的选育以及极小群体的繁育都会发生严重的遗传漂变,导致种性发生变化,发生群体基因频率和基因型频率波动,打破群体遗传平衡。
对于甜菜育种来说,理论上S型胞质材料不适合做授粉系,只有N型胞质的材料才适合选育成授粉系。
猜你喜欢
——白化病儿童肖像作品
中外文摘(2021年24期)2021-12-30
园艺与种苗(2021年10期)2021-11-28
生物学通报(2021年11期)2021-09-28
中国种业(2018年8期)2018-08-28
中国产前诊断杂志(电子版)(2018年1期)2018-07-21
中学生物学(2017年1期)2017-03-23
红领巾·探索(2015年7期)2015-09-02
种子科技(2015年8期)2015-05-24
中国医药导报(2015年26期)2015-02-28
安徽农业科学(2015年13期)2015-01-13
