
杂交稻三系配套的原理及功能研究
2015-05-24 16:14陆作楣张莉
种子科技 2015年8期
陆作楣,张莉
南京神州种业有限公司,江苏 南京 210095
杂交稻三系配套的原理及功能研究
陆作楣,张莉
南京神州种业有限公司,江苏 南京 210095
介绍了杂交稻三系配套的基本原理、三系在配套中的功能定位以及对保持系功能的一些错误认识,供杂交稻育种工作者学习参考。
杂交稻;不育系;保持系;恢复系;三系配套;原理;功能
中国杂交稻的成功,始于1970年发现崖县普通野生稻花粉败育株(野败),1973年实现三系配套,1975年开始大面积示范推广杂交稻优势组合。从此,三系配套一直被认为是杂交稻成功的关键。
1 杂交稻三系配套的基本原理
三系是指水稻雄性不育系、水稻雄性不育保持系和水稻雄性不育恢复系,简称为不育系、保持系和恢复系。三系最早配套的过程是将浙江选育的几个早籼稻品种作为父本,与野败不育株杂交,杂交F1表现雄性不育的植株继续与原来的父本回交,如此连续进行4~5次,育成与原父本品种相似的不育系。例如:珍汕97A、威20A等,原来的父本品种成为该不育系的保持系,如珍汕97和威20记作珍汕97B、威20B。随后用不育系测交筛选恢复系,得到多个恢复力好的品种,包括泰引1号、IR24、IR26、古154等。其中珍汕97A×IR24定名为汕优2号,珍汕97A×IR26定名为汕优6号,威20A×IR26定名为威优6号,它们均表现出很强的杂种优势和较好的综合性状,成为今后大面积推广的杂交稻优良组合。
研究表明,不育系和保持系的细胞核内都有两对隐性的雄性不育基因(rf3rf3rf4rf4),不育系的细胞质线粒体有一对不育基因,它与两对细胞核不育基因相互作用,形成了雄性不育的特性,故常被称为质核互作型不育系。但保持系的细胞质内线粒体没有相应的不育基因,所以,虽然细胞核内也存在两对不育基因,保持系仍然表现为正常可育。恢复系的细胞核内有两对显性的恢复基因(Rf3Rf3Rf4Rf4),它与不育系杂交的后代(F1)具有不育细胞质的背景,而核基因呈杂合状态(Rf3rf3Rf4rf4),两个恢复基因抑制了两个等位的不育基因与不育细胞质基因的互作,使F1的雄性恢复正常,从而使杂交稻能够正常结实。
2 三系在配套中的功能定位
从三系配套中不育系、保持系、恢复系之间的关系看,定型的不育系接受保持系的花粉,异交结实的种子仍为不育系,保持系自交产生下一代保持系种子,两者共同构成了不育系和保持系的繁殖过程,可以用以下简图表示:
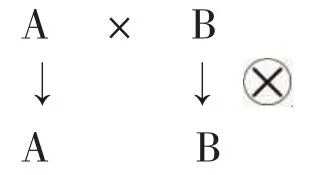
另外,不育系接受恢复系的花粉,异交结实产生杂交稻种子(F1),同时恢复系自交产生下一代恢复系种子,完成杂交稻制种过程,也可以用以下简图表示:

在以上不育系繁殖和杂交制种的过程中,三系分工协作、各司其职,分别以异交和自交的方式,实现了三系的自身繁殖和杂交种的生产过程。由此可见,三系配套的直接功能是保证杂交稻安全供种及在通常条件下能够正常结实。但是,这种三系间恢复系、保持系关系的配套,并不能保证杂交稻具备生产上要求的杂种优势。而要解决这个问题,需要将三系配套的概念引入到一个新的层次,即三系间配合力的优良匹配。
众所周知,影响农作物产量的基因主要是细胞核基因,杂种优势的强弱受细胞质的影响并不明显。杂交稻的杂种优势主要取决于父本恢复系和母本不育系的配合力,而配合力的高低取决于优良的加性基因和非加性的部分显性基因的集合。与一、二个不育基因或恢复基因之间的协调配套相比,三系间配合力的协调与配套要复杂和困难得多。不育系的细胞核基因都是由保持系多代回交置换的结果,所以不育系的配合力几乎完全由保持系决定。换句话说,杂交稻的杂种优势是由恢复系和保持系共同决定的,而不育系只是起了一个媒介的角色。我国最早育成的几个杂交稻优良组合,都是在完成三系恢复系、保持系关系配套的同时,又完成了双亲配合力的协调,其原因就在于应用了原有的优良品种作为保持系和恢复系,前者如珍汕97和威20,后者如IR24和IR26,它们都是中外水稻育种家长期选育的成果,成为选育杂交稻强优势组合的物质基础。只有具备了这样的理性认识,才能对三系的功能进行正确的定位。
3 对保持系功能的误判需要纠正
从三系间恢复系、保持系关系配套的表面现象看,保持系主要是为不育系传宗接代服务的,因而只是杂交稻种子生产中的一个配角,特别是在杂交稻起步阶段,转育不育系所用的保持系都是国内现成的常规品种,得来容易,转育快速,因而淡化了选育保持系的重要性。另外,我国三系杂交稻的父母本分属两个杂种优势群,父强母弱是普遍现象,杂交稻的超亲优势都是和恢复系作比较,不少育种工作者和种业界人士顺理成章地认为,杂交稻的优势主要取决于恢复系,这种片面的认识进一步使保持系的地位边缘化。
最典型的事例莫过于著名杂交稻组合汕优63的选育过程。母本珍汕97A是20世纪70年代育成的,父本明恢63则是70年代末到80年代初育成并配组的,两位育种家因此获得科技界的最高荣誉。可是早在20世纪60年代育成珍汕97(B)的育种家却被遗忘了将近30年,浙江温州地区农科所林恭松团队选育的珍汕97(B)、威20(B)、协青早(B)等优良品种,成为中国三系杂交稻应用最广和最久的保持系,一直到了21世纪初,浙江省有关部门才组织了该项成果的鉴定会,后来该项成果获得了浙江省科技进步一等奖,但至今与任何国家级奖项无缘,林恭松这个名字也从未在关于杂交稻历史的记载和报道中出现过。
以上这种重不育系轻保持系的思想意识,还长期充斥在所谓不育细胞质资源的挖掘过程中。在20世纪的最后30年里,杂交稻研究人员先后育成多种不育细胞质类型的不育系,其中应用面积较广的有矮败型、冈型、D型、K型、印尼水田谷型、红莲型等。在当时的认识水平上,这些不同来源不育细胞质的发现和应用,有一定的科学价值。
长期以来,人们习惯将不育系配组的杂交稻组合称为某某型杂交稻,同时对其不育系或杂交稻作出评价,诸如矮败型分蘖力强、米质好(长粒型),冈型、D型稻瘟病抗性好,印尼水田谷型、红莲型和K型的异交习性好,制种产量高,组合的杂种优势强、米质好。不过在冷静思考之后,以上这些类型的种种优点,似乎无法和不育细胞质联系起来,无论是农艺性状还是配合力,基本上都是细胞核基因控制的。随后的研究发现,除红莲型不育系外,其他类型的不育系都有相同的恢复系、保持系关系,同一个保持系可以转育成多种同核异质的不育系,而且表现几乎完全一致。华南农业大学的研究进一步指出,所谓不同类型细胞质的线粒体不育基因,并没有发现差异,因而除了红莲型外,其他类型统统可以归入野败型这个大类。实际上不育系的改良都是从改良保持系做起的,不育细胞质的影响几乎可以忽略不计。所以至今对不同细胞质来源的不育系的评价,可以说是张冠李戴。
如今,越来越多的有识之士都已逐步认识到保持系在增强不育系选育的预见性、保障不育系育性稳定和制种安全以及保护知识产权等方面都具有不可替代的作用。三系杂交稻虽然在错误导向下承受了重重压力,但并未失去其顽强的竞争力。丢掉包袱,开动机器,把不育系选育提高到新的水平,是杂交稻育种的当务之急。
1005-2690(2015)08-0032-02
S511
A
2015-04-30
猜你喜欢
园艺与种苗(2021年10期)2021-11-28
蔬菜(2021年7期)2021-11-27
粮食科技与经济(2020年7期)2020-10-20
中国种业(2018年11期)2018-11-16
中国种业(2018年8期)2018-08-28
麦类作物学报(2018年6期)2018-07-02
园艺与种苗(2015年8期)2015-12-28
江苏农业科学(2015年1期)2015-04-17
安徽农业科学(2015年13期)2015-01-13
发明与创新·大科技(2009年5期)2009-05-31
