
广东主要森林类型林分生物量和碳储量模型研建*
2024-01-20 10:02郭泽鑫胡中岳
林业科学 2023年12期
郭泽鑫 胡中岳 曹 聪 刘 萍
(1.华南农业大学林学与风景园林学院 广州 510642;2.国家林业和草原局中南调查规划院 长沙 410014)
森林作为陆地生态系统的主体,在维持陆地碳循环和稳定全球气候方面发挥着主导作用。2018 年,《美国国家科学院院刊》(Proceedings of the National Academy of Sciences of the United States of America,通常简称为PNAS)以专辑形式刊发中国科学院“碳专项”系列成果,在中国陆地生态系统中,森林生态系统碳储量占比38.9%,2001—2010 年年均固碳1.63亿t,贡献了超过80%的陆上固碳量(Tanget al.,2018;Fanget al.,2018)。且由于过去大规模造林行动,现阶段仍存有大量幼中龄林,森林平均碳密度较低(Huanget al.,2012;Huet al.,2015),意味着未来仍有很大碳封存空间,持续的森林碳汇监测尤为重要。森林碳储量是碳汇和碳汇潜力研究的基础,碳储量可通过含碳系数由森林生物量导算。近几十年来,国内外开展了大范围、多尺度、多种类的生物量建模工作,以单木模型为主(Sileshi,2014;Luoet al.,2020),林分模型较少。然而,由于并非所有森林资源调查均提供每木检尺数据,如二类调查只能获取小班的林分因子,导致单木模型应用受到限制,因此有必要建立林分模型作为数据缺失下森林生物量估算的补充。近年来,欧洲和部分亚洲国家的相关研究逐渐增多(Khanet al.,2018;Jagodzińskiet al.,2019;Aguirreet al.,2021),国内也几乎同步推进,但研究对象仍主要集中在北部和东北部地区(Donget al.,2019;曾伟生等,2021;何潇等,2021),针对南方树种建立林分生物量或碳储量模型的研究较少。
目前,广东地区已有相关研究开始填补这方面的空白,建立了林分水平的理论生长方程(曹磊等,2020;黄金金等,2022a;2022b)和异速生长模型(薛春泉等,2022),为区域尺度森林碳存量和固碳潜力研究奠定了基础。但尚有一些问题未被考虑:一是理论生长方程依赖于林龄的确定,可获取时间序列内任一节点的预测值,但在分析有调查数据的某一节点的生物量时,相较直接反映林分生长的变量,林龄对生物量的影响是间接的,且天然林年龄的确定较为复杂,易进一步造成误差传递,估算结果存在较大不确定性;二是异速生长模型主要基于林分胸高断面积和平均树高构建,这2 个因子无法在年度更新的森林资源档案数据库或森林资源管理“一张图”中获取,导致应用受限;三是上述研究所建模型均为总量模型,无法单独估算地上或地下生物量,不利于生物量分配规律研究,且在遥感生物量建模中,因提取到的影像因子主要反映森林冠层和高度等地面以上信息,故建模的目标变量一般为地上生物量,需要相应统计模型获取地上生物量的观测值,也正是由于地上生物量与常见调查因子或遥感参数关联性较强、相对易测且占比较高,其在实践中往往更受关注,是全球气候观测系统54 个基本气候变量之一,因此,单独建立地上生物量模型有其实践性和必要性。此外,在气候持续变化背景下,许多研究者开始关注气候因子对生物量的影响,建立气候敏感的单木或林分生物量模型(Fuet al.,2017a;Heet al.,2021;Zenget al.,2021),但少有系统针对同一区域多个树种开展研究。鉴于此,本研究基于2007、2012 和2017 年3 期广东省森林资源连续清查数据,利用非线性误差变量联立方程组构建省内12 种主要森林类型与蓄积量兼容的地上和地下生物量、地上和地下碳储量模型;以哑变量形式区分树种结构,以再参数化方法建立气候敏感的林分生物量和碳储量模型,采用从简单到复杂的渐进性设计方案,为省内储量数据的本底摸查、省级与县市级储量数据的有效衔接提供模型支撑;研究树种结构和气候条件对模型的影响和作用机制,为更精细的碳汇监测及森林质量提升提供理论指导。
1 数据来源与研究方法
1.1 数据来源
本研究基础数据包含固定样地调查数据和气候数据2 部分。固定样地调查数据源自2007、2012 和2017 年广东省森林资源连续清查数据,样地面积0.066 7 hm2,地类为乔木林地,仅统计活立木中的林木部分,排除散生木和四旁树。森林类型划分主要依据样地内各树种蓄积量占比,除针阔混交林外,其余森林类型优势树种蓄积占比不低于65%。根据2017 年清查数据确定面积和蓄积占比较多的12 种森林类型为建模对象,分别为马尾松(Pinus massoniana)林、湿地松(Pinus elliottii)林、杉木(Cunninghamia lanceolata)林、桉树(Eucalyptusspp.)林、栎树(Quercusspp.)林、木荷(Schima superba)林、相思(Acaciaspp.)林、其他软阔林、其他硬阔林、针叶混交林、阔叶混交林和针阔混交林。剔除异常数据和离群点后,获得建模样地3 040 块,除相思林外,其余森林类型均达到大样本(样本量达50 个以上)要求。各森林类型建模样地的林分特征统计量见表1,其中平均胸径为主林层优势树种胸径的平方平均值,平均树高为3~5 株平均木树高的算术均值。
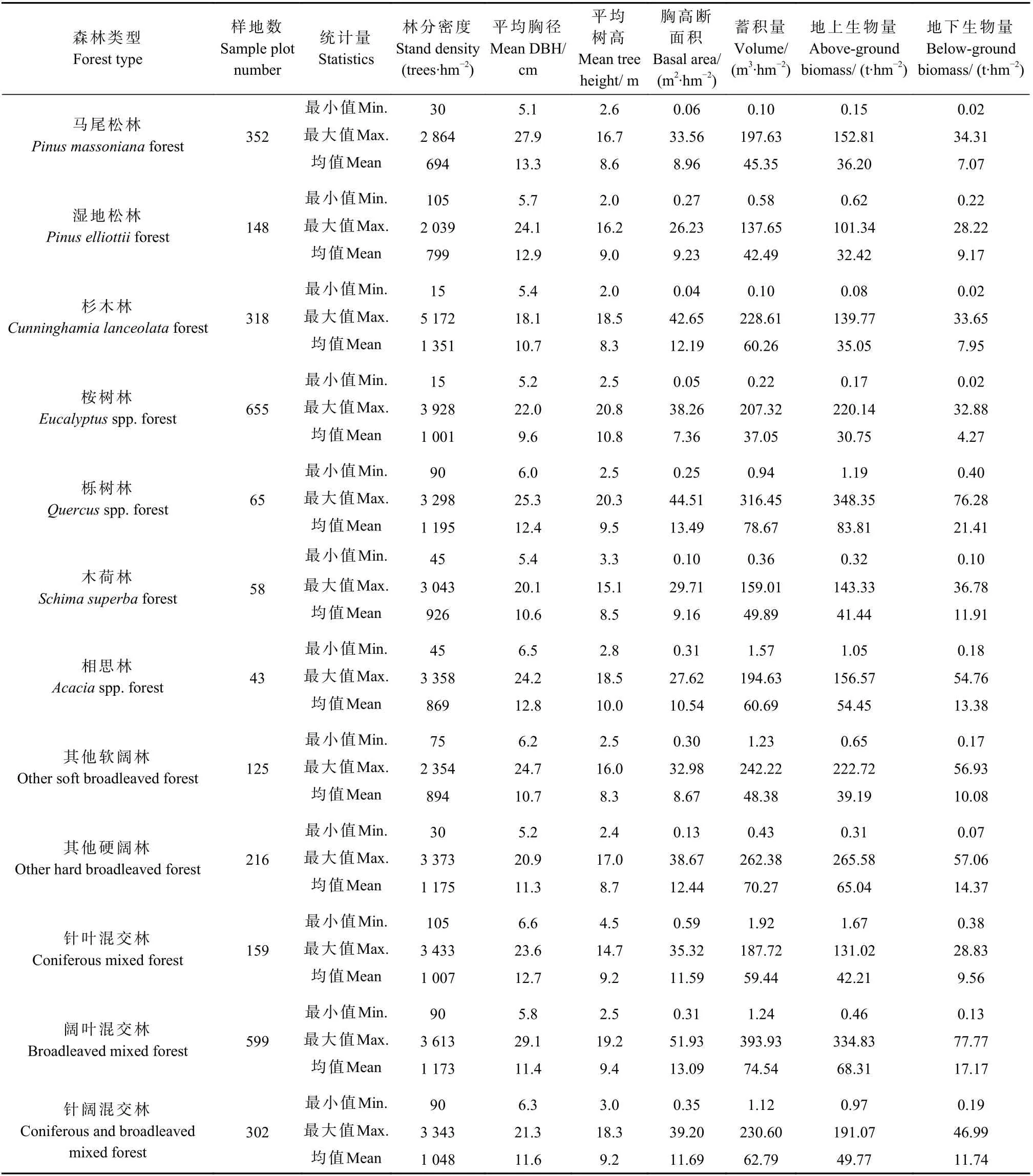
表1 建模样地林分特征统计量Tab.1 Summary statistics of stand characteristics for modeling sample plots
气候数据提取自ClimateAP(Wanget al.,2017),将固定样地的地理坐标和海拔作为输入数据,生成各样地的逐年气候变量,每个变量取5 年调查间隔期的均值。考虑到广东省的气候特征,选用与植物生长密切相关的8 个变量参与后续研究,包括年平均温度(mean annual temperature,MAT)、最热月平均温度(mean warmest month temperature,MWMT)、最冷月平均温度(mean coldest month temperature,MCMT)、最热月与最冷月均温差(temperature difference between MWMT and MCMT,TD)、年平均降水量(mean annual precipitation,MAP) 、 年热湿比指数( annual heat:moisture index,AHM)、一年中5 ℃以上积温(degreedays above 5 ℃,DD5)、哈格里夫斯气候水分亏缺(hargreaves climatic moisture deficit,CMD)。
1.2 林分生物量和碳储量计算
利用已发布的单木生物量模型计算各树种每木器官(干、皮、枝、叶、根)生物量,乘以相应器官含碳系数得到每木器官碳储量,各器官汇总得到单木地上、地下生物量和碳储量,样地内单木数据累加得到林分地上、地下生物量和碳储量。马尾松、湿地松、杉木、柳杉(Cryptomeria fortunei)、 栎树、 木荷、 枫香(Liquidambar formosana)采用全国立木生物量模型和碳计量参数行业标准计算生物量(国家林业和草原局,2014a;2014b;2014c;2016a;2016b;2016c;2016d),桉树、其他软阔类和其他硬阔类估算模型源自《中国森林植被生物量和碳储量评估》(李海奎等,2010),相思估算模型源自《中国主要林木生物量模型手册》(罗云建等,2015),其中相思、其他软阔类、其他硬阔类为二元模型,基于叶金盛(2006)建立的广东树种相对树高曲线模型估算树高。其余伴生树种则采用曾伟生等(2012)和曾伟生(2017)提出的通用性地上生物量模型及相应根茎比、木材密度估算。
前期报道已实测了广东省内十余种树种的含碳系数(张红爱,2018;徐期瑚等,2018a;2018b;2019),几乎覆盖本研究主要建模树种,且多数显示出与全国平均水平的显著差异,因此本研究优先采用已有报道含碳系数计算各树种碳储量,其次采用上述全国行业标准的参考值。以上有独立器官含碳系数的树种占全部建模样本总蓄积比例超98%,其余伴生树种视地上、地下含碳系数相同,采用《中国森林植被生物量和碳储量评估》(李海奎等,2010)数值计算。
1.3 基础林分生物量和碳储量模型
林分生物量建模一般有2 种途径:一是直接构建基于林分变量的生物量模型;二是基于林分生物量与蓄积量的转化关系构建生物量转扩因子(biomass conversion and expansion factor,BCEF)模型(Donget al.,2019;Jagodzińskiet al.,2019)。为同时提供2 种方法且保证估算结果的一致性,本研究建立与蓄积量兼容的地上和地下生物量、地上和地下碳储量模型,利用非线性误差变量联立方程组(唐守正等,2008),通过BCEF、根茎比和含碳系数将五者关联起来,其中地上生物量基于BCEF 由蓄积量转化,地下生物量通过根茎比由地上生物量导出,地上和地下碳储量则分别通过相应林分平均含碳系数由地上和地下生物量得到。单木尺度上,材积由树干形数、断面积和树高决定,因而在林分尺度上,本研究以林分胸高断面积和平均树高作为蓄积量的预测因子。此外,许多研究显示林分生物量与胸高断面积、平均胸径、平均树高等存在显著相关性(González-Garcíaet al.,2013;Jagodzińskiet al.,2018;2019;2020),其中胸高断面积和平均胸径均反映树木的径向生长,但林分模型一般应优先选择胸高断面积,因为林分是单木的聚集体,其生物量不仅与树木生长相关,同时受林分密度影响,而胸高断面积兼顾林分这两大特征,因此,生物量和碳储量模型也采用胸高断面积和平均树高作为预测变量,得到第一个模型系统(M-1),见式(1)。由于平均树高获取难度相对大、准确性难以保证,本研究也提供仅包含胸高断面积的一元模型(M-2),限于篇幅,不单独列出。
尽管胸高断面积优于平均胸径,但其属于小班调查中的间接计算因子,无法在森林资源档案数据库或森林资源管理“一张图”中获取,因此本研究以平均胸径和林分密度的组合变量近似替代胸高断面积,构建不依赖胸高断面积和平均树高的生物量和碳储量模型,即第三个模型系统(M-3),见式(2)。

1.4 区分树种结构的林分生物量和碳储量模型
除上述林分生长指标外,林分生物量和碳储量还与树种结构、龄组、起源等定性因子存在一定关系。由于广东省森林以幼中龄林为主,且各森林类型的起源也较为单一,故不单独考虑龄组和起源的影响。但绝大多数森林并非是由单一树种组成的绝对纯林,而不同树种采用不同的单木生物量估算模型和含碳系数,因此有必要研究树种结构对林分生物量和碳储量的影响。本研究参照《国家森林资源连续清查技术规定(2014)》,将除混交林以外的其他森林类型按优势树种蓄积占比划分为纯林(≥90%)和相对纯林(65%~90%)。设置变量S为反映不同树种结构的哑变量(唐守正等,2008),S=1 时表示纯林,S=0 时表示相对纯林,以M-1 为基础构建第四个模型系统(M-4),见式(3)。M-2 和M-3 为数据缺失情况下的补充模型,不作拓展。
式中:mi1、ai1、bi1、ci1、di1(i=0,1,2)为哑变量S的参数。
1.5 气候敏感的林分生物量和碳储量模型
由于同一树种采用固定的含碳系数计算碳储量,本研究视区域内林分平均含碳系数仅与树种结构相关,气候对碳储量的影响主要通过联立方程由蓄积量和生物量体现。对气候变量作初步筛选后保留数量仍有8 个,为简化模型和降低变量间共线性,需作进一步筛选。以各森林类型V、AGB 、BGB为因变量,初筛8 个气候因子为自变量,进行逐步回归,进入和剔出模型的临界显著性水平分别为0.05 和0.10。计算剩余变量的方差膨胀因子(VIF),剔除VIF 最大变量后重新逐步回归,直至所有变量VIF 均小于5。将逐步回归筛选得到的气候变量以再参数化方式引入到M-1 各基础参数中,得到第五个模型系统(M-5),见式(4)。但由于各气候变量并非对每一参数都有显著影响,因此对V、AGB 、BGB模型进行独立非线性回归,并对气候变量作参数t检验,剔除P值最大的变量后再次拟合,重复该过程直至所有变量P值均小于0.05,所得模型回代至M-5 中进行联合估计。
式中: Clj为气候变量,mi j、aij、bij(i=0,1,2;j=1,2,3···)为 Clj的参数。
1.6 异方差与模型评价
蓄积量、生物量和碳储量的异速生长模型普遍存在异方差现象,本研究采用通用权函数处理异方差,权函数形式为W=1/f(x)λ(曾伟生,2013),f(x)为原模型本身, λ取1~2,每个模型反复测试直至找到合适的λ值。确定权函数后,利用ForStat 的非线性误差变量联立方程组模块进行模型系统的参数估计。
采用曾伟生等(2011)提出的6 项基本评价指标进行模型评价,分别为决定系数(R2)、估计值的标准差(standard error of estimate,SEE)、总相对误差(total relative errors,TRE)、平均系统误差(mean systematic errors,MSE)、平均预估误差(mean prediction errors,MPE)和平均百分标准误差(mean percent standard errors,MPSE)。由于本研究各模型系统参数个数差异较大,故将R2修改为调整决定系数(R2a),见式(5)。同时通过F检验分析模型系统M-4 和M-5 与M-1 是否存在显著差异,见式(6)。曾伟生等(2011)研究认为适用性检验否决模型只是小概率事件,建议利用全部样本进行建模,故本研究不划分检验样本。
式中:n为样本量;p为模型参数个数;SSEbase、dfbase分别为M-1 的残差平方和、自由度, SSE 、 df分别为M-4或M-5 的残差平方和、自由度。
2 结果与分析
2.1 基础模型拟合与评价结果
模型系统M-1 拟合与评价结果见表2。以马尾松林分模型为例,c0和d0的估计值实际就是地上和地下部分的林分平均含碳系数,由于采用联合估计,式中AGC 和BGC模型可直接由AGB 和BGB模型乘以相应含碳系数得到。从马尾松林评价指标可见,碳储量模型的SEE 约为相应生物量模型的一半,这实际是含碳系数作用的结果,而其余指标值则非常接近,这种规律在其他森林类型和模型系统中也均有体现,因此下文将不再单独列出碳储量模型的评价指标,仅提供相应的含碳系数。
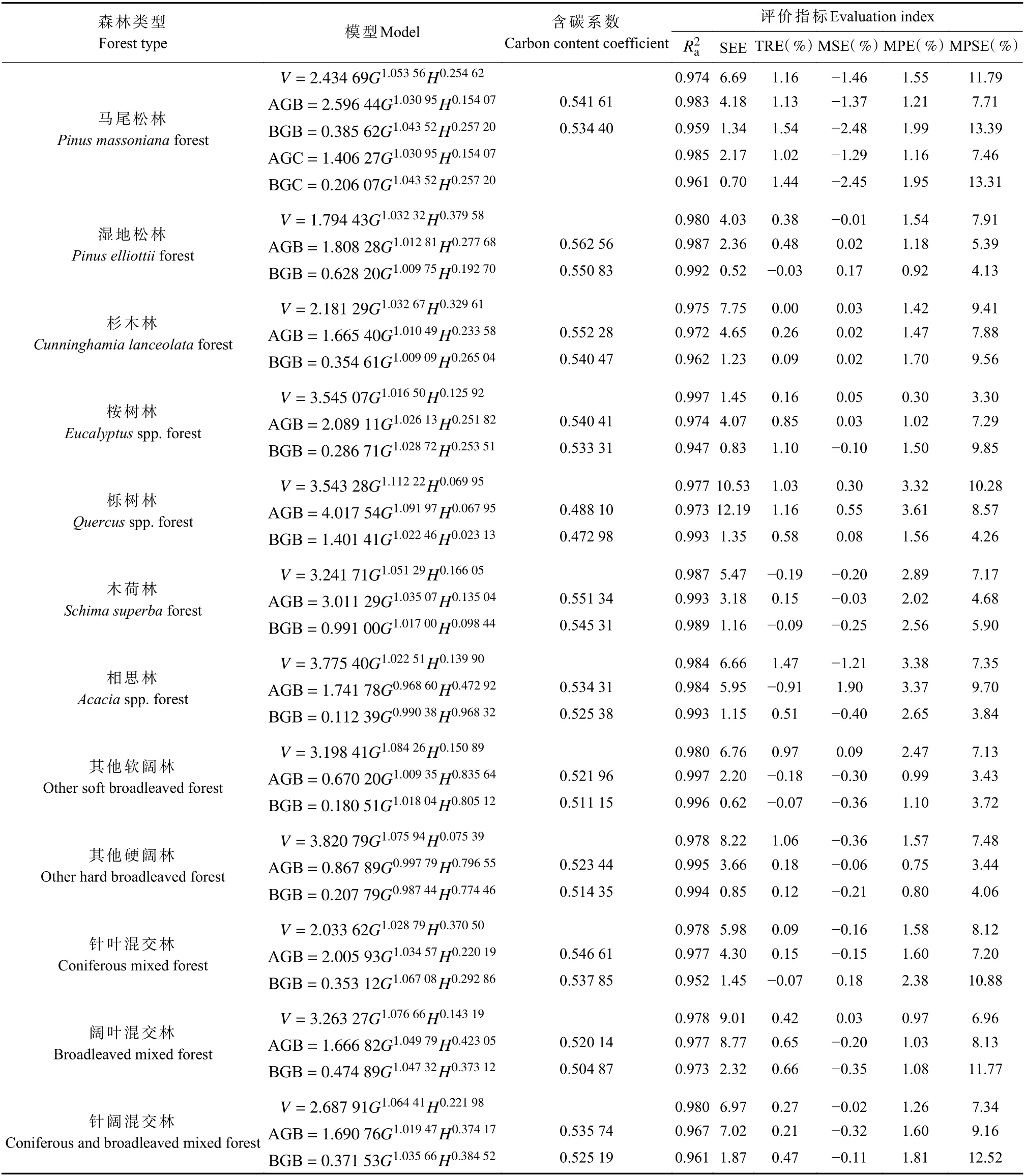
表2 模型系统M-1 拟合与评价结果①Tab.2 Fitting and evaluation results of model system M-1
分析12 种森林类型拟合与评价结果可看出,无论蓄积量、生物量还是碳储量模型,胸高断面积的指数值最稳定,且均维持在1.0 左右,其他2 个参数值则因森林类型、目标变量不同存在较大变异。模型为0.947~0.997,蓄积量模型SEE 为1.45~10.53 m3·hm-2,生物量模型SEE 为0.52~12.19 t·hm-2,地下生物量模型该值相对较小,主要与其基础数值低于地上生物量有关。TRE 在±1.54% 范围,MSE 在±2.48% 范围,MPE为0.30%~3.61%,MPSE 为3.30%~13.39% 。TRE 和MSE 反映拟合效果,理想情况下应控制在±3%或±5%范围;MPE 和MPSE 反映模型精度,前者一般要求保持在3%以内,后者要求保持在15%以内。模型系统M-1 中,仅栎树林、相思林部分模型MPE 略超3%,其余指标值均达到上述标准。以马尾松林M-1 模型残差为例(图1),加权回归(右)较未加权回归(左)异方差得到有效处理。

图1 马尾松林M-1 模型未加权(左)和加权(右)回归残差分布Fig.1 Distribution of unweighted(left) and weighted(right) regression residuals of model system M-1 of Pinus massoniana forest
模型系统M-2 和M-3 拟合与评价结果见表3,限于篇幅,部分指标不再列出,但对指标范围进行描述。从参数估计结果来看,无论M-2 还是M-3,模型指数值均稳定维持在1.0 左右。对于M-2,为0.876~0.996,仅相思林地下生物量模型低于0.9。TRE 除相思林地下生物量模型为7.64%外,其余均在±3.19%范围,MSE 则均在±2.74% 范围。MPE 和MPSE 也表现为相思林地下生物量模型较大,分别为11.30%和20.84%,其余模型MPE 为0.36%~4.70%,大多不超过3%,MPSE 则为4.18%~15.61%。
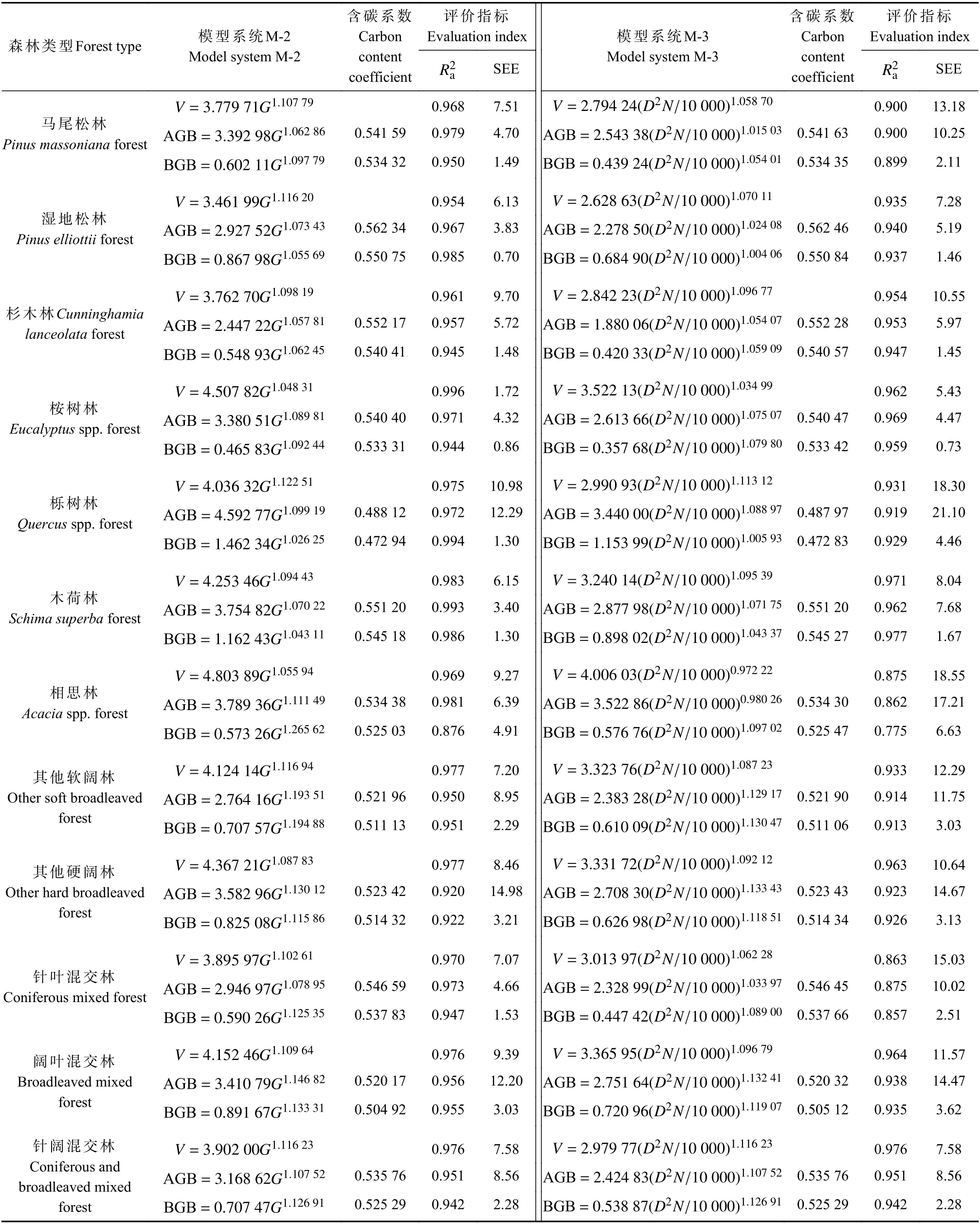
表3 模型系统M-2 和M-3 拟合与评价结果Tab.3 Fitting and evaluation results of model systems M-2 and M-3
图2 所示为基础模型预测值与观测值之比的分布情况,可看出,各模型的中位数和平均值基本均在1.0 附近,M-1 数据分布更为集中,M-2 和M-3 离散程度增大,尤其M-3,远离1.0 的数据增加,异常数据点也更多,其中相思林最为明显。总的来说,M-1 表现最优, M-3 最差,但基本均能达到较高的预测精度。
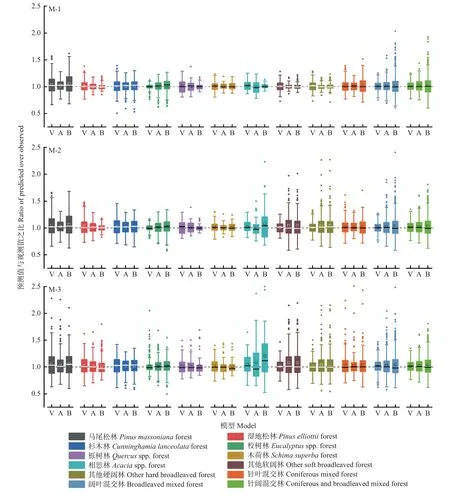
图2 基础模型预测值与观测值之比的分布Fig.2 Distribution of the ratio of predicted over observed of the basic models
2.2 区分树种结构模型拟合与评价结果
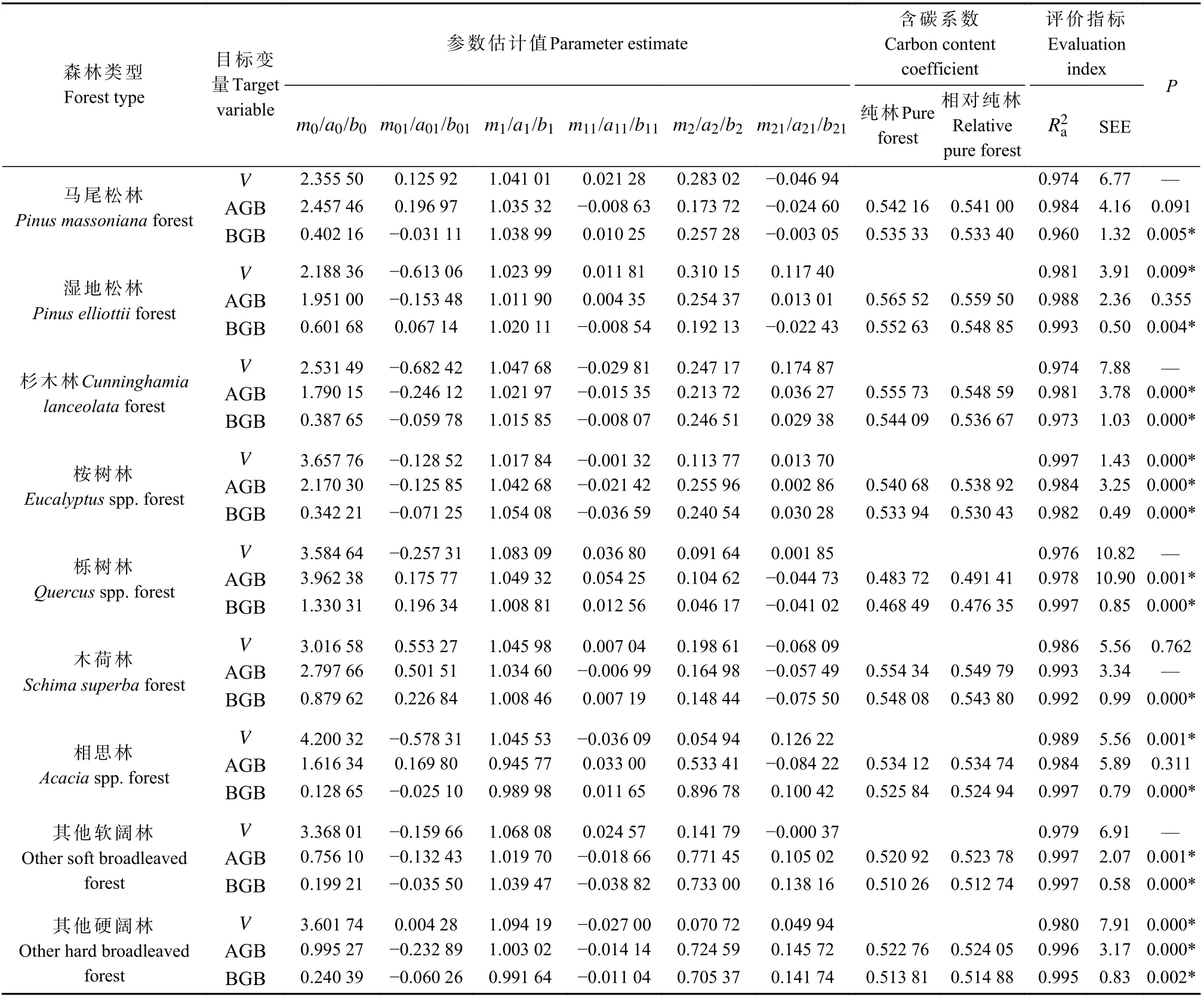
表4 模型系统M-4 拟合与评价结果①Tab.4 Fitting and evaluation results of model system M-4
2.3 气候敏感模型拟合与评价结果
模型系统M-5 拟合与评价结果见表5,相思林在逐步回归过程中所有气候变量均未能达到进入模型的显著性水平,杉木林、木荷林和其他硬阔林筛选得到的气候变量在再参数化过程中均未能通过参数t检验。最终构建马尾松林等8 种森林类型气候敏感的林分生物量模型,所得模型为0.947~0.998。TRE 在±1.86%范围内,MSE 在±1.96%范围内,MPE 为0.29%~2.65%,MPSE 为3.18%~13.29%,四个指标均达到既定标准。从M-5 与M-1 的F检验结果来看,栎树林、其他软阔林、阔叶混交林和针阔混交林所有模型均较M-1显著改进,马尾松林、桉树林部分生物量模型较M-1显著改进,针叶混交林仅蓄积量模型有显著改进,湿地松林所有模型均不受气候变量显著影响。
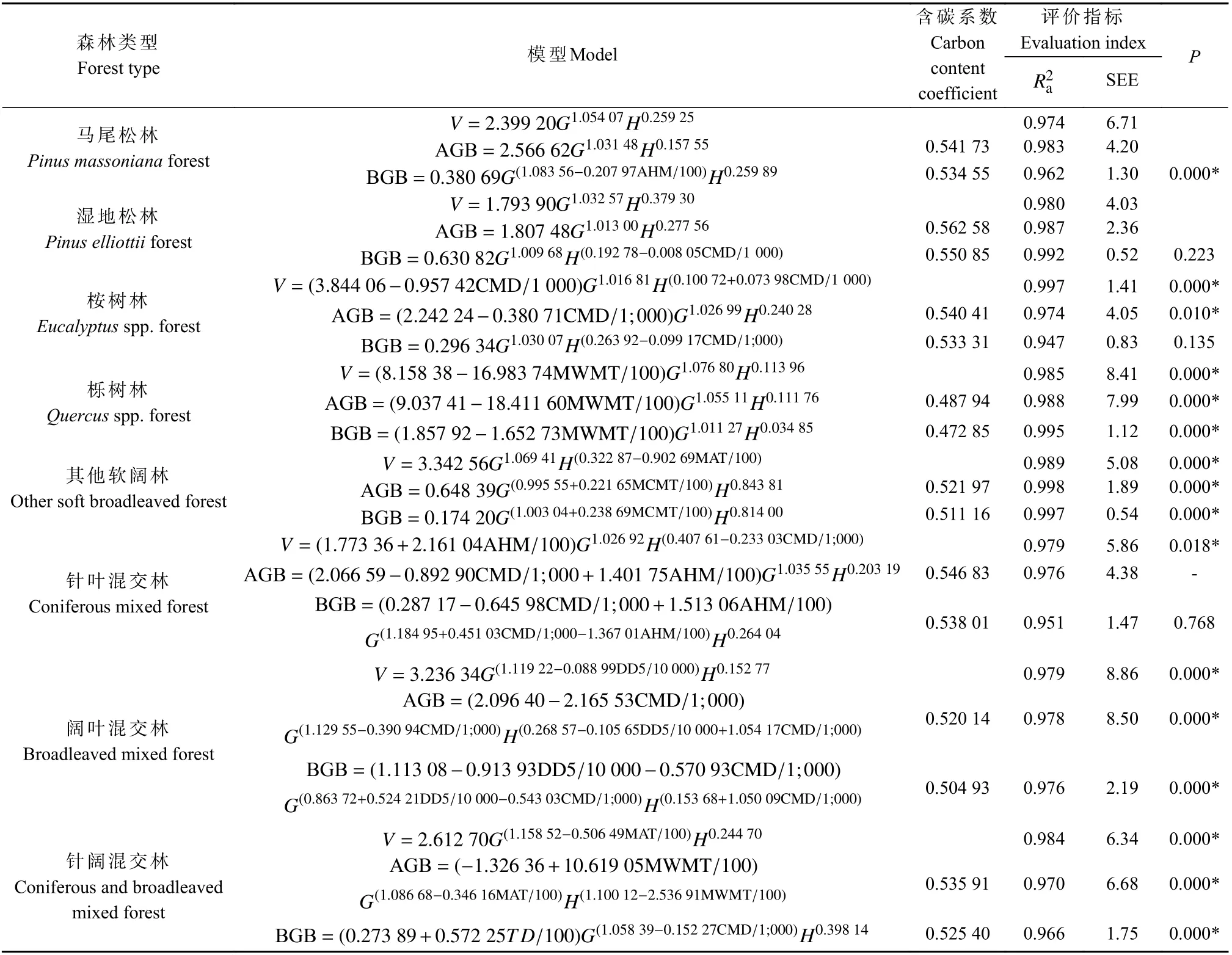
表5 模型系统M-5 拟合与评价结果①Tab.5 Fitting and evaluation results of model system M-5
观察模型参数,由于胸高断面积可能存在小于1的情况,为便于分析相关关系,将位于胸高断面积指数上的气候变量转移至其他参数上,结果显示参数显著的前提下符号未改变。马尾松林地下生物量与AHM(样本范围为10.9~27.2 ℃·mm-1)呈负相关,而AHM=(MAT+10)/(MAP/1 000),表明马尾松林地下生物量与MAT 呈负相关或与MAP 呈正相关,又或二者皆有。湿地松林地下生物量与CMD(118.8~425.6 mm)呈负相关,CMD 是参考蒸散量与降水量的差值,表明湿地松林地下生物量与蒸散量呈负相关或与降水量呈正相关,又或二者皆有。桉树林生物量也与CMD(106.2~540.0 mm)呈负相关。栎树林蓄积量和生物量均与MWMT(23.8~29.1 ℃)呈负相关。其他软阔林蓄积量与MAT(14.9~23.0 ℃)呈负相关,生物量与MCMT(3.5~15.1 ℃)呈正相关。针叶混交林蓄积量和地上生物量与AHM(12.6~22.6 ℃·mm-1)呈正相关,与CMD(102.2~413.0 mm)呈负相关。阔叶混交林蓄积量和地上生物量均与DD5(3 465.0~6 824.4 ℃)呈负相关,表明随着一年中5 ℃以上日均温总和的增加,阔叶混交林蓄积量和地上生物量反而降低。针阔混交林蓄积量和地上生物量与MAT(14.1~23.6 ℃)呈负相关,地下生物量与TD(13.2~23.0 ℃)呈正相关,与CMD(89.0~430.2 mm)呈负相关。此外,还有部分模型同一气候变量在不同参数中符号相反,本研究尝试删除部分重复变量以简化模型,但多数情况下容易造成剩余气候变量参数不显著,这可能是因为该气候变量对蓄积量或生物量的作用不是简单的正负相关。
3 讨论
本研究所建林分模型中,无论何种森林类型、何种模型系统,胸高断面积的指数值均稳定维持在1.0左右,从其他研究建立的模型中也观察到类似规律(Donget al.,2019;曾伟生等,2021),这可能与单木蓄积量和生物量模型中胸径指数值也较为稳定有关。实际上,早有研究尝试从理论层面推导单木地上生物量M与胸径D0的异速生长关系,影响最大的是基于生态代谢理论(metabolic theory of ecology)推导的M=a(a为参数)(Westet al.,1999),此外,还有几何相似模型M=a、恒应力相似模型M=a(Sileshi,2014),国内曾伟生等(2012)通过干形的维数分析,从几何学角度推导得到M=a。可见,对于
不同树种单木生物量模型而言,胸径的异速生长指数在理论层面上是恒定的,在经验层面上是稳定的,这使得作为单木聚集体的林分,其生物量等储量模型也表现出胸高断面积指数值的稳定性。M-3 中替代胸高断面积的组合变量D2N也表现出类似规律,该模型假设样地内所有样木胸径均为平均胸径D,则G∝D2,这也解释了胸高断面积的指数约为单木模型中胸径指数的一半。
本研究所建基础模型大多能达到较好的拟合效果和预测精度,整体上M-1 表现最优,M-3 最差,因此,在胸高断面积可获取的情况下,应优先选择M-1 和M-2,一类调查虽无法直接从样地表获取胸高断面积,但可利用样木胸径计算得到,二类调查虽没有样木表,但提供小班的角规断面积,对于胸高断面积确实无法获取的数据库,如森林资源管理“一张图”,可用M-3替代计算。此外,本研究提供了区分树种结构的林分模型,但仅按纯林和相对纯林2 种类型进行建模。实际上,树种结构或树种组成是一个复杂多变的因素,即便在一定区域范围内,潜在的树种组合及比例也非常丰富,这种可变性难以用单一林分变量进行量化。因此,本研究的出发点是在提供大范围森林生物量和碳储量估算模型的前提下,确保纯林的估算精度,以满足小尺度或单一纯林估算的需要,尤其是杉木林、桉树林等人工纯林较多的森林类型。
气候尤其温度和降水直接或间接影响森林生物量或碳储量的积累,一些研究甚至显示气候比土壤在预测森林生物量上贡献更大(Bennettet al.,2020)。本研究12 种森林类型中有8 种全部或部分生物量模型对气候变量敏感,且多数在提高生物量预估精度上具有统计学意义,这表明即便在相对较小的区域范围内,也有必要考虑气候对生物量估算的影响。一方面,随着时间推移,气候环境可能发生持续变化;另一方面,海拔等地理要素也会导致垂直向的地带性气候。一般在特定范围内,充足的水热条件能够推动树木生理过程、促进树木生长发育,进而增加森林生物量和碳储量,但气候具有时空差异性,树木适生条件和抗逆能力也有种间特异性,因而相关研究往往不能在气候对森林生物量和碳储量的影响上得出一致结论。Zeng 等(2017 ;2021) 研究表明中国油松(Pinus tabuliformis)、杨树(Populusspp.)生物量与年平均温度正相关。Fu 等(2017a;2017b)研究表明中国马尾松、落叶松(Larixspp.)地上生物量与最湿季平均温度负相关,Sullivan 等(2020)研究表明热带森林碳储量与最高温度负相关。本研究多数森林类型生物量与温度负相关,仅在一年中的低温时段才显示出与温度正相关,如其他软阔林生物量与最冷月平均温度呈正相关,针叶混交林地上生物量虽与AHM 正相关,但对模型精度并没有显著影响。Wang 等(2019)研究显示珠三角地区森林碳储量与年平均降水量和年平均最低温度呈正相关,与年平均最高温度呈负相关,与本研究结论一致。此外,本研究显示多数森林类型生物量与蒸散量呈负相关或与降水量呈正相关,值得注意的是,直接反映降水的指标MAP 未能进入到任一模型中,取而代之的是综合反映蒸散和降水的CMD 以及综合反映温度和降水的AHM,结合广东省的气候特点及本研究多数森林类型生物量与温度的负相关性,推测区域内限制森林生物量增长的水分因素可能不是降水不足,而是由于温度过高导致蒸发过多。总的来说,对于广东省主要森林类型,温度过高可能是限制生物量和碳储量增长的最主要因素。
4 结论
本研究基于2007、2012 和2017 年3 期广东省森林资源连续清查数据,利用非线性误差变量联立方程组构建省内12 种主要森林类型蓄积量、地上和地下生物量、地上和地下碳储量的基础林分模型,并分别以哑变量和再参数化方法建立区分树种结构和气候敏感的林分生物量和碳储量模型,得出以下结论:
1) 基于胸高断面积和平均树高的林分蓄积量、生物量和碳储量模型具有较好的拟合效果和较高的预测精度,仅基于胸高断面积的简化模型和基于平均胸径、林分密度的补充模型也仍有较好表现,可用于数据缺失下森林生物量和碳储量估算。林分模型中,胸高断面积指数较为稳定,基本维持在1.0 左右。
2) 区分树种结构对各森林类型全部或部分生物量和碳储量模型具统计学意义,具有更详细森林调查数据时应优先采用此模型,尤其是估算小范围或单一纯林的生物量和碳储量时。
3) 所建气候敏感的林分生物量和碳储量模型多数显著优于相应基础模型。温度过高、蒸散过多或降水不足是限制广东各森林类型生物量和碳储量增长的主要因素。
猜你喜欢
矿产勘查(2020年4期)2020-12-28
矿产勘查(2020年2期)2020-12-28
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
山东林业科技(2018年6期)2019-01-08
山东林业科技(2017年1期)2017-06-29
林业与生态(2016年2期)2016-02-27
亚热带资源与环境学报(2015年3期)2015-01-22
断块油气田(2014年5期)2014-03-11
