石珊瑚共附生真菌次级代谢产物的抗炎活性及化学多样性研究
2024-01-12 12:41:22廖清楠周龙建杨志友冯昀铠黄谊君胡雪琼3张翼刘亚月
生物技术通报 2023年12期
廖清楠 周龙建,2,3 杨志友,2,3 冯昀铠 黄谊君 胡雪琼,3张翼,2,3 刘亚月,2,3
(1. 广东海洋大学食品科技学院 广东省水产品加工与安全重点实验室 广东海洋大学深圳研究院海洋医药研发中心 湛江市脑健康海洋药物与营养品重点实验室,湛江 524088;2. 南方海洋科学与工程广东省实验室(湛江),湛江 524006;3. 海洋食品精深加工关键技术省部共建协同创新中心 大连工业大学,大连 116034)
炎症是临床常见的一个病理过程,是机体组织受到刺激或损伤后所产生的一种防御性应答反应,主要以红、肿、热、痛甚至组织器官功能丧失为症状[1-2]。炎症是一把双刃剑,适度的炎症反应对人体是有益的;但过度、失控的炎症反应则会导致组织的病变和二次损伤,是许多疾病的发病基础。现代药理学研究表明,在系统性血管炎、肠道克罗恩病等一些自身免疫性疾病以及神经系统病变的疾病中,炎症反应发生时,白细胞介素‑1、白细胞介素‑6、PGE2 等炎性相关因子的表达都有所上升,进而伤害到人体内部细胞,并对人体产生危害。目前,这些疾病正在全球范围蔓延,并且已演变成严重危害人类健康和社会经济可持续发展的重要公共卫生问题,越来越受到各国政府和社会的高度重视。因此,开发治疗炎症的有效药物对多种疾病的治疗具有重要的意义。
珊瑚,隶属于腔肠动物门中的珊瑚虫纲,是一种常见的海洋低等无脊椎动物。作为珊瑚礁生态系统的重要组成部分,广泛分布于我国海南、西沙群岛及南沙群岛等热带海岸地区。珊瑚表面和组织内部聚集着丰富的微生物资源[3],这些微生物与珊瑚宿主在长期的共同进化过程中,通过合成如抗菌、抗肿瘤、抗病毒和抗污损等[4-5]具有生物活性的次级代谢产物,共同抵御外敌入侵和病害防御,一直是药物先导化合物的重要来源之一。从珊瑚共附生的真菌中分离的代谢产物化学结构多样,涉及生物碱、聚酮、萜类、内酯类、环肽类、聚醚类、甾醇类、多糖类及不饱和脂肪酸等非甾体类化合物类型。目前,人们已经对多种珊瑚来源的共附生真菌进行了研究,并从中获得了一系列具有抗肿瘤、抗炎、抗氧化、抗癌等多种生物活性物质。如李金凤等[6]从乳白肉芝软珊瑚(Sarcophyton glaucum)分离得到1 个新型西松烷型二萜,显现出良好的抗炎活性。李旺盛等[7]对软珊瑚Dendronephthyasp.的化学成分和抗炎活性进行研究发现,化合物11‑acetoxy‑3β,6α-dihydroxy‑9, 11‑seco‑5α-cholest‑7‑en‑9‑one 对LPS 诱导的BV‑2 细胞的炎症反应具有抑制作用。Liu 等[8]在珊瑚真菌土曲霉中发现10 种丁烯内酯类化合物,活性结果表明,versicolactone B 对LPS 诱导的RAW 264.7 小鼠巨噬细胞中 NO 的产生有很强的抑制作用,比阳性对照吲哚美辛的作用更显著,这表明它可能是一种潜在的新型抗炎药的先导化合物。中山大学的Liu 等[9]从柳珊瑚真菌Penicillium sclerotiorum中分离得到4 种新的聚酮化合物,均能显著抑制 LPS 诱导的巨噬细胞 RAW 264.7 中NO 的产生,且sclerketide B 和C,以及iso‑chromophilone IX 在mRNA 水平上表现出下调iNOS和COX‑2 表达。Shen 等[10]从软珊瑚Lobophytum sarcophytoides中分离到2 种cembrane 型二萜类化合物,其中cembratriene‑4,8,11‑triol 能显著抑制LPS 激活的RAW 264.7 细胞中NO 的产生。但是,现有活性先导化合物的研究只涉及珊瑚资源中的一小部分,且国内外对珊瑚共附生微生物的研究多集中在柳珊瑚或软珊瑚等,对其他珊瑚科,尤其是石珊瑚及其共附生真菌来源的抗炎活性先导化合物的发掘及其作用机制的研究却鲜有报道。
本研究中以徐闻石珊瑚来源共附生真菌为研究对象,探究不同培养条件对其次级代谢产物抗炎活性和化学多样性的影响。分别选用PDB 和糙米培养基,并设置 3 种盐度(0.3%、3% 和 10%),对 31株石珊瑚来源共附生真菌进行小规模发酵培养。以LPS 诱导的BV‑2 小胶质细胞为炎症模型,从中筛选出具有抗炎活性的菌株;并通过TLC 指纹图谱、UPLC‑QTOF‑MS 以及FBMN 等技术手段进一步分析抗炎活性菌株次级代谢产物的化学多样性,以期为后续深入挖掘石珊瑚共附生真菌中的抗炎活性次级代谢产物奠定基础。
1 材料和方法
1.1 材料
1.1.1 真菌菌株和细胞 本实验的31 株石珊瑚共附生真菌均采自广东徐闻珊瑚礁自然保护区,目前保藏在广东海洋大学海洋药物研究所,菌株种属及珊瑚来源如表1 所示;BV‑2 小胶质细胞购于武汉大学中国典型培养物保藏中心。
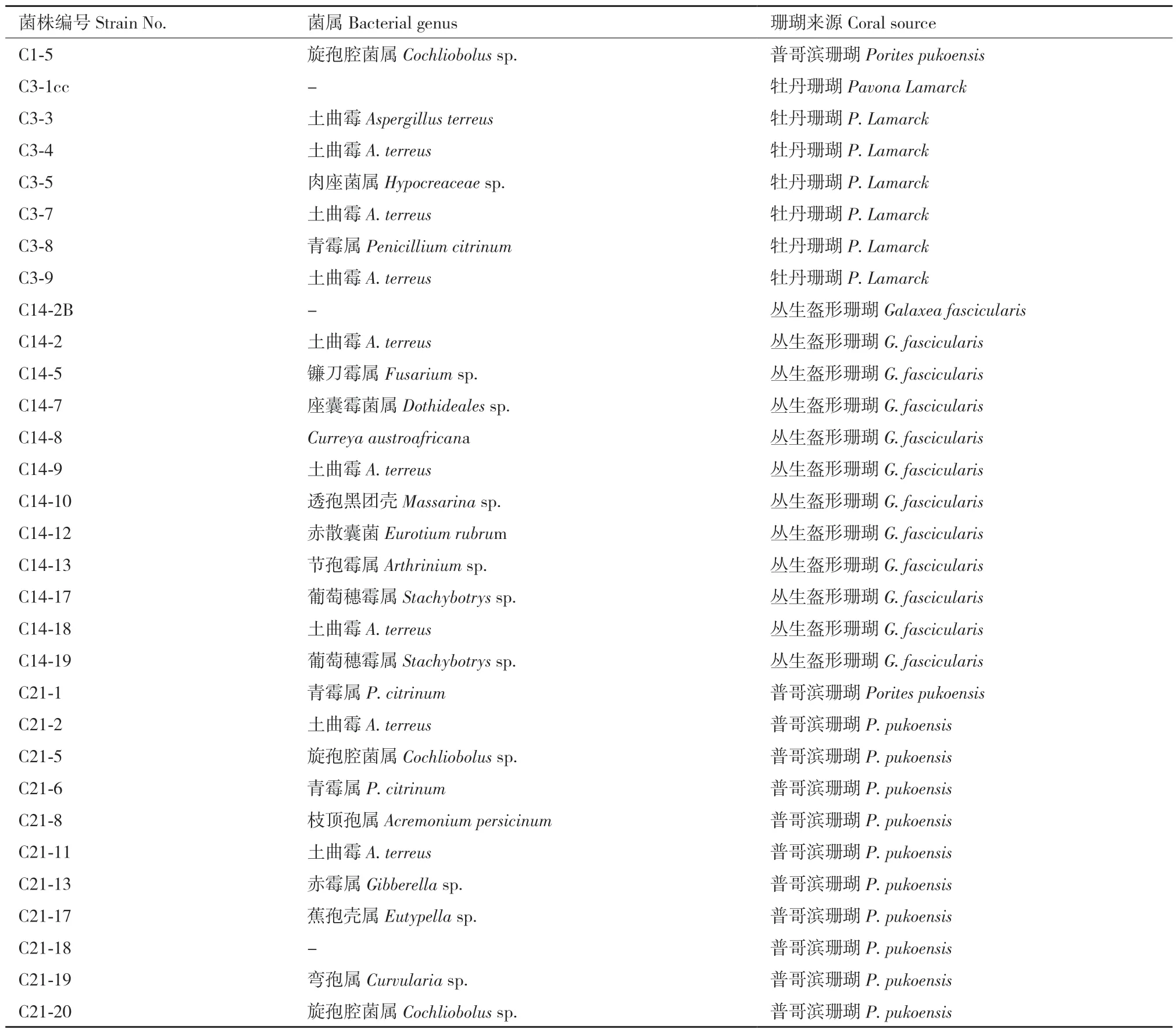
表1 31 株珊瑚共附生真菌菌属及珊瑚来源Table 1 31 coral symbiotic fungal genera and coral sources
1.1.2 培养基 PDB 培养基:配置好的马铃薯葡萄糖粉24.0 g,海盐适量(盐度为0.3%时添加量为3.0g,盐度3.0%时添加量为30.0 g,盐度10.0%时添加量为100.0 g),加水定容至1 L,pH =7.0,121℃灭菌30 min 备用。糙米培养基 :糙米50.0 g,海盐适量(盐度为0.3%时添加量为3.0 g,盐度3.0%时添加量为30.0 g,盐度10.0%时添加量为100.0 g),加水定容至1 L,pH =7.0,121℃灭菌30 min 备用。完全培养基:向500 mL DMEM(dulbecco’s modified eagle medium)高糖培养基中加入50 mL 过滤后的澳洲胎牛血清(fetal bovine serum, FBS)和5 mL 双抗(青霉素-链霉素混合液100×)。
1.1.3 主要试剂与仪器 DMEM 高糖培养基,Gibco公司;胰蛋白酶‑EDTA(0.25%),Gibco 公司;FBS,美国Zeta Life 公司;青霉素-链霉素混合液(100×),Gibco 公司;LPS,Sigma 公司;磷酸盐缓冲液(PBS),Gibco 公司;NO 检测试剂盒,碧云天公司;3‑(4,5‑二甲基‑2‑噻唑基)‑2,5‑二苯基溴化四唑(MTT),源叶生物。
多功能紫外透射仪,上海精科实业有限公司;霉菌培养箱及生物安全柜,上海博讯实业有限公司医疗器械厂;生物安全柜,上海博讯实业有限公司医疗器械厂;UPLC/Xevo G2‑XS QTOF 超高效液相/串联四极杆飞行时间质谱仪,沃特世科技(上海)有限公司;旋转蒸发仪,上海爱朗仪器有限公司;超净工作台,上海博讯实业有限公司;CO2细胞培养箱,美国精骐有限公司;倒置显微镜,日本岛津仪器有限公司;细胞计数仪,Countstar BioMed;Epoch2 酶标仪,购自美国BioTek 公司;离心机,湖南湘仪实验室仪器开发有限公司;高效薄层层析板(GF254),购自青岛凯邦公司。
1.2 方法
1.2.1 种子液的制备 将31 株石珊瑚来源共附生真菌冻存管置于温度 28℃、湿度 80% 的霉菌培养箱活化过夜后,接种到已灭菌的海水马铃薯琼脂(potato dextrose agar,PDA)培养基的平板上,将平板置于霉菌培养箱中培养,待菌落孢子生长茂盛,接种到预先装有200 mL 灭菌PDB 培养基的锥形培养瓶中培养 3-4 d,待菌落孢子生长茂盛备用。
1.2.2 OSMAC[11]策略下菌株的小规模培养 将平板长势良好的31 株菌株在无菌条件下制备成种子液并分别接种到6 种不同培养基(盐度0.3%、3%、10%的PDB 培养基,盐度0.3%、3%、10%的糙米培养基),每组设3 个平行组,于室温条件下静置培养28 d。
1.2.3 菌株次级代谢产物的提取 将培养后的菌株分别用不同方法提取次级代谢产物,PDB 培养基的菌体和菌液分开,菌体用等体积甲醇进行超声提取3 次,菌液用等体积的乙酸乙酯萃取3 次,减压浓缩得粗提物;糙米培养基用等体积甲醇浸泡,超声提取30 min,重复3 次,甲醇提取液减压浓缩得粗提物。
1.2.4 细胞培养及给药 将冻存的BV‑2 细胞迅速解冻后,将细胞冻存液转移到离心管中并加入5 mL 完全培养基混合,1 000 r/min 离心5 min,弃去培养基,重新加入5 mL 完全培养基将贴壁的细胞吹打下来转移到培养瓶继续培养,备用。把处于对数生长期的细胞制成细胞悬液,将细胞以2×104个/孔的密度100 μL/孔的体积接种到96 孔板中,分为多组培养(control 组,LPS 组(1 μg/mL),试验组(1、10、20、50、100 μg/mL),每组设5 个平行,放入培养箱培养24 h。
1.2.5 NO 测定 本实验通过碧云天NO 试剂盒测定细胞中的NO 含量。其原理是:在有机酸条件下,对氨基苯磺酸和NO2-发生重氮化反应,然后再与盐酸-萘胺偶合生成紫红色物质,测定OD540nm值易知NO2-含量,进而推算出NO 含量。待测样品所用溶液将1 mol/L 标准品稀释为不同浓度(0、1.56、3.13、6.25、12.5、25、50、100 μmol/L),吸取30 μL 稀释好的标准品于96 孔板中,加入磺胺溶液30 μL,室温下避光反应5-10 min,再加入盐酸-萘胺溶液30μL,继续在室温避光孵育5-10 min,用酶标仪检测540 nm 处的吸光度。绘制标准曲线,得到回归方程:y= 0.008 7x+ 0.049 3(R2= 0. 997)。细胞接种到96孔板分组培养24 h 后,分别吸取control 组、LPS 组和试验组中培养基30 μL 于新的96 孔板,每孔加入30 μL 磺胺溶液反应5 min 后再加入30 μL 盐酸-萘胺溶液继续反应5 min,使用酶标仪检测540 nm 处的吸光度。
1.2.6 细胞存活率 将细胞以5×103个/孔的密度和100 μL/孔的体积接种到96 孔板中培养,24h 后,加入待测样品,使加入后样品的终浓度分别为1、10、20、50、100 μg/mL;24 h 后,每孔加入10 μL 的MTT 溶液(用PBS 溶解成5 mg/mL),置于37℃、5% CO2的培养箱中继续培养4 h,弃去培养基,每孔加入DMSO 溶液100 μL 溶解甲瓒,振荡混匀后检测波长540 nm 处的吸光度(λ)值,计算细胞存活率:细胞存活率(%)=样品组OD540/空白组OD540×100%。
1.2.7 菌株次级代谢产物的TLC 分析 将上述粗提物用甲醇配置成10 mg/mL 的浓缩液,采用高效硅胶薄层层析板对各粗提物的化学多样性进行分析。选择乙酸乙酯∶正己烷=2∶1 二元溶剂体系作为TLC分析展开剂,并在波长254 和365 nm 下观察代谢产物的分布情况。酚类化合物在自然界中广泛存在,在治疗炎症方面也起到了良好的效果[12],用铁氰化钾-三氯化铁试剂进行显色,对次级代谢产物中的酚类化合物进行追踪分析[13]。
1.2.8 LC‑MS/MS 分析 将各粗提物用色谱级甲醇配成等浓度样品溶液进行液质分析。液相色谱条件:进样量为1.0 μL,洗脱条件为:0.00-0.50 min,30%的乙腈水溶液(含0.1%甲酸);0.50-4.00 min,30%-80%的乙腈水溶液(含0.1%甲酸);4.00-7.00min,80%的乙腈水溶液(含0.1%甲酸);7.00-7.20 min,80%-30%的乙腈水溶液(含0.1%甲酸);7.20-8.50 min,30% 的乙腈水溶液(含0.1%甲酸),流速:0.3 mL/min。DAD 检测器信号采集波长为190-600 nm,监测波长为 254 和 360 nm。色谱柱为:Waters ACQUITY UPLC BEH RP18(2.1 mm×50 mm,1.7 μm),质谱的质量扫描范围设置为m/z50-2 000,电喷雾离子化,正离子模式。
1.2.9 FBMN[14]可视化分析 将原始液质数据文件的格式转换为.abf 格式后,将其上传至MS‑DIAL 基于化合物的母离子精确质量和二级质谱的相似度与数据库中的化合物进行匹配。然后通过MSFINDER Ver 3.52 将特征点列表里的化合物与不同的数据库如PubChem、NPASS、COCONUT 和ChEBI 等进行比较,以注释每个特征点可能代表的化合物。然后上传至全球天然产物分子网络集群数据库(global natural products social molecular networking,GNPS)数据库计算分析,将分析结果导入Cytoscape 软件绘制分子网络图,利用FBMN 基于GNPS 的二级质谱相似度匹配功能可注释样品所含的已知化合物,并根据分子网络图中不同节点的关系,结合质谱数据,以推断化合物种类及结构。
1.2.10 数据统计分析 采用GraphPad Prism 8 软件,采用单因素方差分析(ANOVA)进行数据统计分析,P<0.05 被认为差异具有统计学意义,P<0.001 被认为差异有高度统计学意义。
2 结果
2.1 菌株次级代谢产物的抗炎活性分析
小胶质细胞(BV‑2)是中枢神经系统的固有免疫细胞,响应中枢神经系统疾病,特别是神经炎症相关疾病的病理刺激。本文以LPS 诱导的BV‑2 小胶质细胞为炎症模型,对31 株石珊瑚来源共附生真菌次级代谢产物的抗炎活性进行筛选,结果如表2和表3 所示。PDB 培养基条件下,共有17 株菌株的次级代谢产物在不同浓度下显示出一定的抗炎活性,其余14 株菌株均不显示或者显示弱(P≤ 0.05)的抗炎活性。且随着盐度的增加,具有抗炎活性菌株明显增加。而糙米培养基条件下,27 株菌株的次级代谢产物呈现出显著的抗炎活性,但随着盐度的增加,菌株抗炎活性无明显差异。上述结果表明,不同培养条件对菌株抗炎活性的影响很大,如C1‑5、C3‑3、C3‑8、C3‑9 和C14‑17 在PDB 培养条件下菌株次级代谢产物均不显示抗炎活性,但在糙米培养基的3 个盐度条件下均表现出良好的抗炎活性。同时上述5 株真菌的粗提物对NO 的抑制能力均表现出显著的浓度依赖性,即随着浓度的增大抑制NO产生的能力也随之加强(图1),尤其是菌株C3‑9在低浓度条件下(10 和20 μg/mL)也表现出了显著的NO 抑制活性。
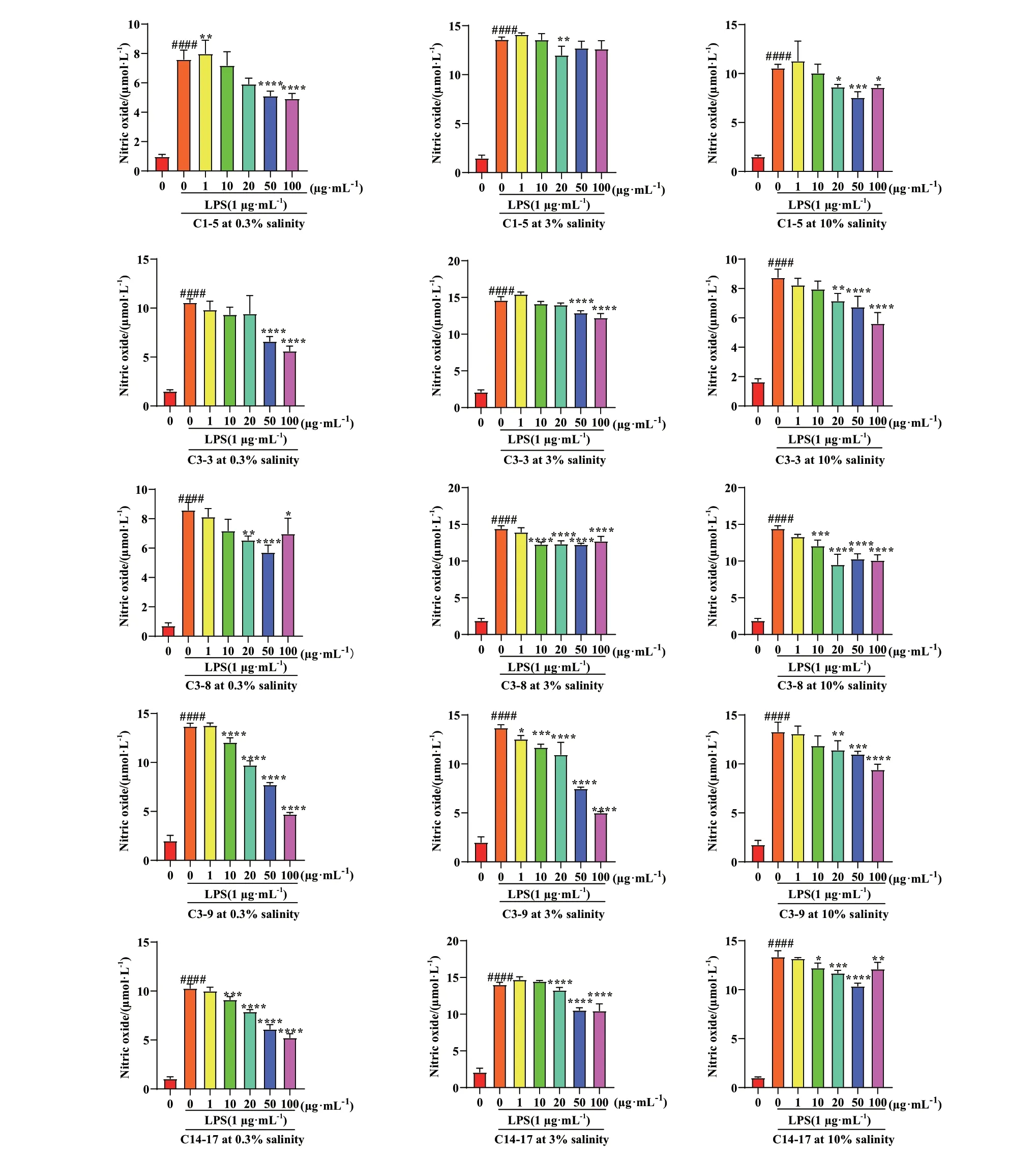
图1 糙米培养基条件下5 株共附生真菌粗提物对BV-2 细胞NO 产生的影响Fig. 1 Effects of crude extracts from five strains of symbiotic fungi on NO production in BV-2 cells under brown rice medium conditions
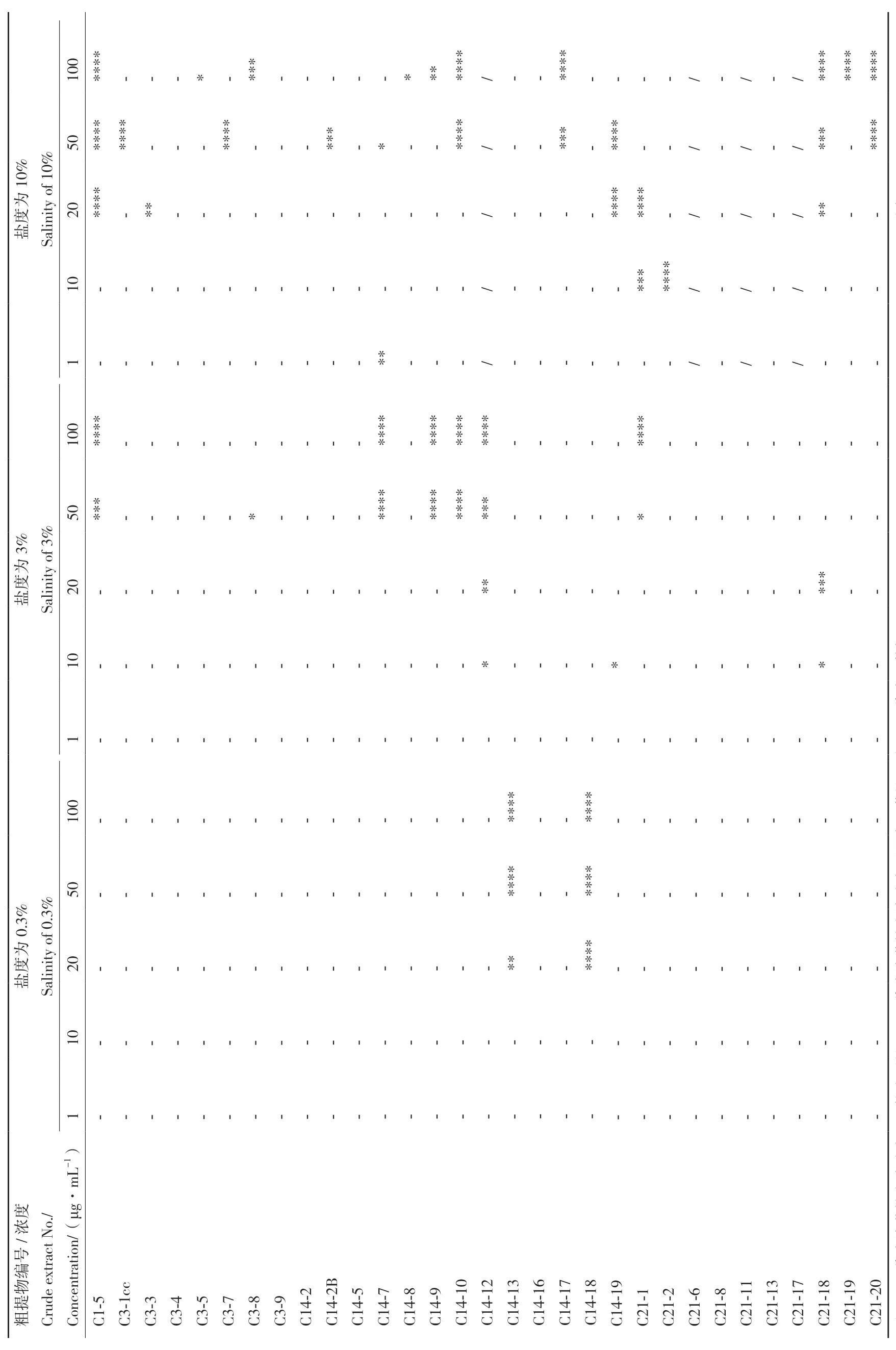
表2 PDB 培养基条件下各菌株粗提物的抗炎活性Table 2 Anti-inflammatory activities of crude extracts extracted from different strains under PDB medium conditions
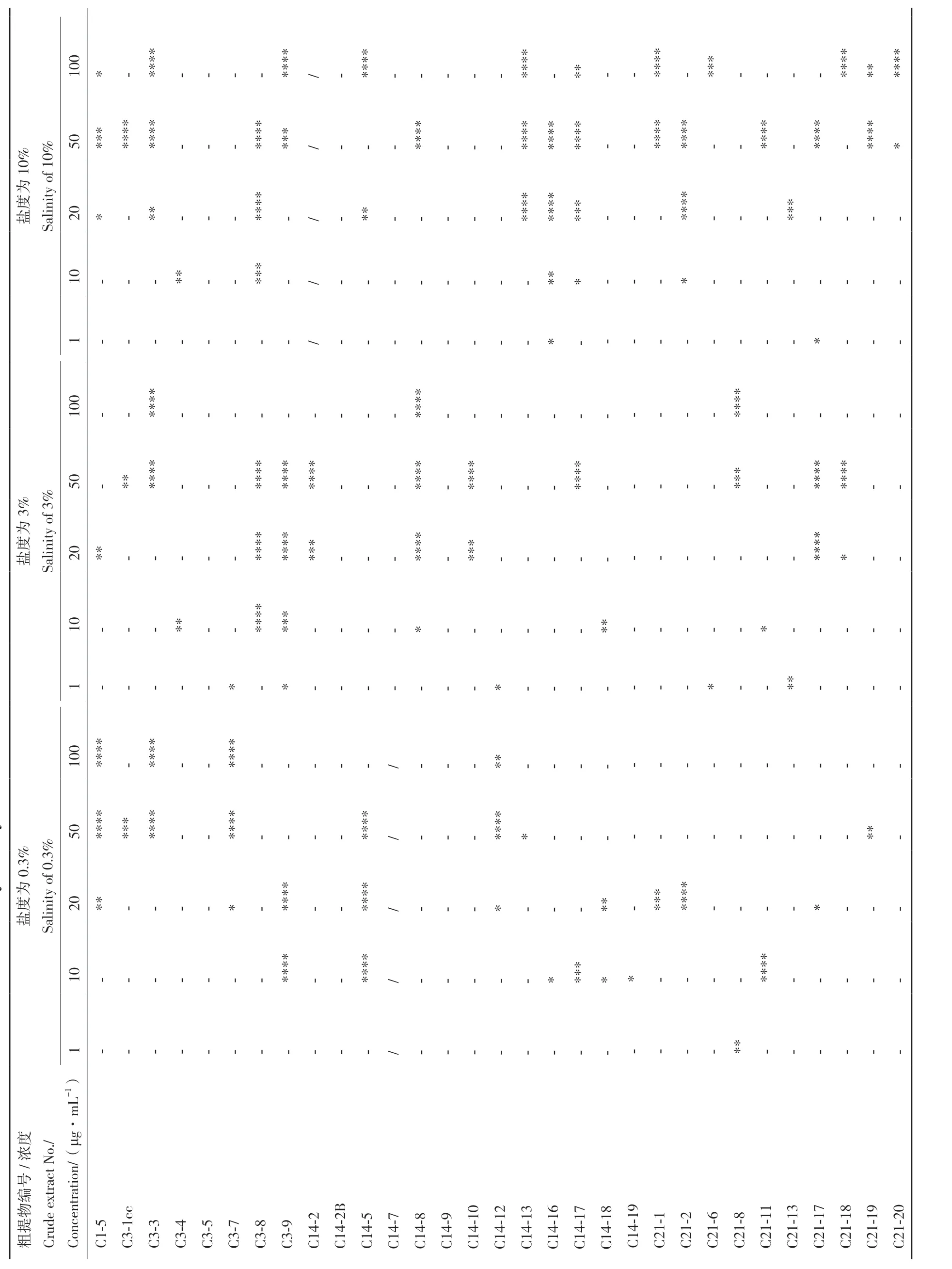
表3 糙米培养基条件下各菌株粗提物的抗炎活性Table 3 Anti-inflammatory activity of crude extracts obtained from different strains under brown rice medium conditions
通过MTT 实验检测样品对BV‑2 小胶质细胞存活率的影响,进而确定实验的样品浓度水平。如图2 所示,大部分实验组细胞在用粗提物(浓度依次为1、10、20、50、100 μg/mL)处理24 h 后,其OD540nm值和空白组OD540nm值的比值约为100%,说明在这个浓度范围里,粗提物对BV‑2 小胶质细胞的存活率无显著性影响。因此,在此浓度梯度下,大部分粗提物没有细胞毒性,可用此浓度梯度继续实验。
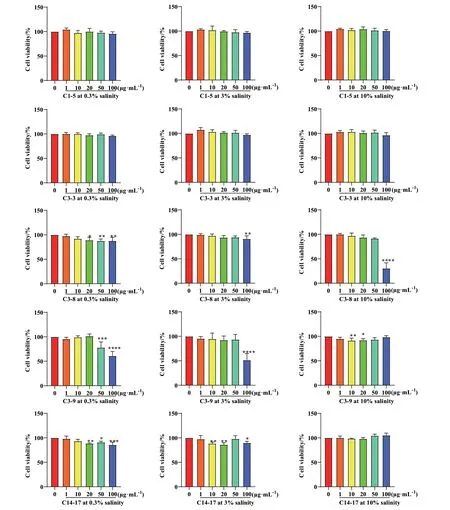
图2 糙米培养基条件下5 株共附生真菌粗提物对BV-2 细胞存活率的影响Fig. 2 Effect of crude extracts of five strains of symbiotic fungi on the viability and survival of BV-2 cells under brown rice medium conditions
2.2 菌株次级代谢产物的TLC图像分析
2.2.1 5 株具有抗炎活性的菌株表观形态对比 根据上述抗炎活性结果,选择C1‑5、C3‑3、C3‑8、C3‑9 和C14‑17 等5 株具有显著抗炎活性的菌株作为目标菌株。通过对比菌株的表观形态(图3)和DNA 测序可知,5 株菌株分别属旋孢腔菌属Cochliobolussp.(C1‑5),土曲霉A. terreus(C3‑3 和C3‑9),青霉属P. citrinum(C3‑8)和葡萄穗霉属Stachybotryssp.(C14‑17),其中3 株来源于牡丹珊瑚,1 株来源于普哥滨珊瑚,1 株来源于丛生盔形珊瑚。
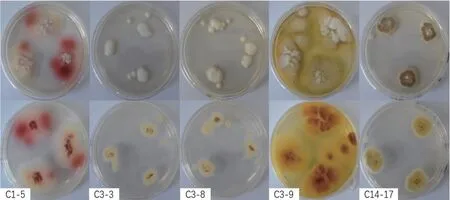
图3 5 株具有抗炎活性的菌株生长形态Fig. 3 Growth morphology of five strains with anti-inflammatory activity
2.2.2 5 株菌株次级代谢产物的TLC 图像分析 利用 TLC 对上述5 株具有抗炎活性菌株的粗提物进行了不同波长下的紫外吸收、荧光斑点、三氯化铁-铁氰化钾显色斑点的 TLC 指纹谱图综合比较,结果如图4 所示,不同培养条件下各菌株次级代谢产物差异较大。相比其他菌株,C3‑9 在波长 254 和 365nm 下吸收斑点明显更多(图4‑C1、C2),这意味着其含有的次级代谢产物种类更为丰富。从图4‑C3 可看出菌株C3‑9 在三氯化铁-铁氰化钾显色剂作用下显现出丰富的蓝色斑点,这表明其具有丰富的酚类产物。进一步分析菌株C3‑9 的TLC 图像可知,PDB和糙米培养基下菌株代谢产物具有较大的差异。且随着盐度的增加,同一培养基下菌株的次级代谢产物也发生了一定的差异,部分次级代谢产物的斑点强度明显降低,这表明盐度的增加,会导致部分代谢产物的产量降低。
2.3 菌株C3‑9次级代谢产物化学多样性分析及FBMN可视化分析
为了进一步分析不同培养条件对菌株C3‑9 代谢情况的影响,对6 个不同培养条件下的提取物进行了 LC‑MS/MS 和FBMN 分析。从提取物的色谱基峰图(图5)可知,菌株C3‑9 在PDB 和糙米培养基中的代谢产物具有一定的差异性,且与上述TLC 分析结果一致。PDB 培养基条件下,菌株次级代谢产物无明显差异,但随着盐度的增加,部分产物的峰面积明显减少,这表明其产量的降低,说明在PDB培养基中盐度对次级代谢产物的产量影响较大。而糙米培养基中,0.3%和3%盐度的条件下的粗提物的种类没有表现出明显的差异性。但是随着盐度的增加,10%盐度下菌株的次级代谢产物丰富度明显降低,且与3%盐度的产物表现出了明显的差异性,这与2.1 部分观察到的3%盐度产物抗炎活性显著高于10%盐度产物的现象具有很好的相关性。而MS‑DIAL 特征离子表积分面积的对应峰在3%和10%两种盐度条件下的积分面积比进一步表明不同盐度条件下的产物产量具有明显的差异(表4),且在3%盐度条件下产生的主要次级代谢产物的产量显著高于10%盐度条件下。从中选择8 个质谱可检测的产量发生显著变化的主峰(峰1-8,差异性倍数>2),将其质谱数据提交到FBMN,并结合COCONUT、PubChem、LOTUS 等多数据库做注释分析(图6),最终确定了这8 个主峰依次为cordycepin、visnagin、thymol、6‑methoxyflavone、periplogenin、spathulenol、ectoine、gallocatechin。查阅文献发现,这8 个化合物均具有抗炎活性。这对于3%盐度产物明显优于10%盐度产物提供了合理的解释。
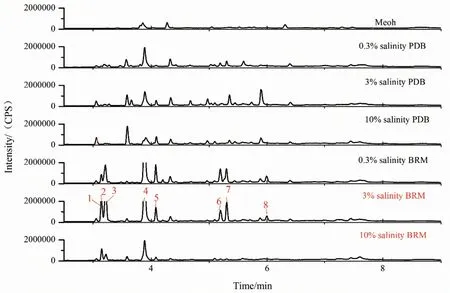
图5 不同培养条件下菌株C3-9 发酵产物的LC-MS/MS色谱基峰图Fig. 5 LC-MS/MS base-peak chromatograms of the fermented products of strain C3-9 under different culture conditions
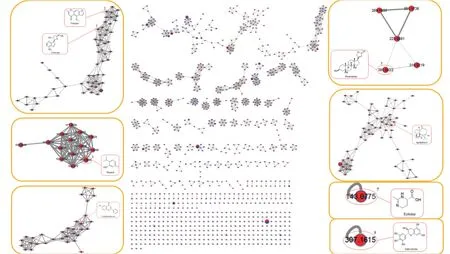
图6 菌株C3-9 在0.3%和10%盐度糙米培养基的代谢产物基于二级质谱联系的分子网络局部放大图Fig. 6 Local magnification of metabolites of strain C3-9 in 0.3% and 10% salinity brown rice medium based on secondary mass spectrometric correlation of molecular network
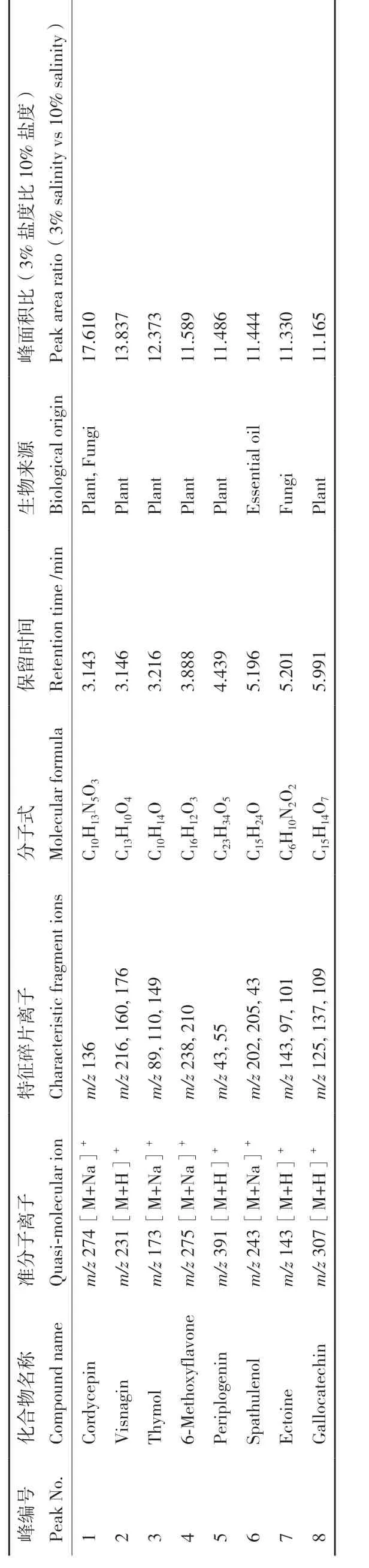
表4 3% 和10% 盐度下菌株C3-9 基峰色谱图中产量具有差异主峰的数据库挖掘及其峰面积的变化倍数Table 4 Database mining of strain C3-9 base peak chromatograms with differential primary peaks in yield at 3% and 10% salinity and the multiplicity of changes in their peak areas
3 讨论
OSMAC 策略[15-17],亦称单菌株多次级代谢产物策略,是通过改变培养基营养成分、状态,添加酶抑制剂,调控表观遗传,混合培养等方法,充分挖掘微生物合成次级代谢产物的能力,从而获得更多类型的次级代谢产物。彭艾等[18]通过OSMAC策略从深海真菌Stachybotryssp.3A0009 中得到7 个单体化合物,其中包括3 个对人口腔上皮癌KB细胞有较强增殖抑制作用的化合物。Si 等[19]用OSMAC 策略从黄曲霉菌株中分离到8 个化合物,其中新化合物flavichalasines N 和O 及已知化合物aspochalasins I 和D 对3 种人癌细胞株有明显的毒作用。通过OSMAC 策略,da Silva 等[20]从A. sydowii中激活沉默的代谢产物,得到具有较好AChE 抑制活性的5 个化合物:cyclo‑(L‑Leu‑L‑Pro)、cyclo‑(L‑Val‑L‑Pro)、cyclo‑(L‑Val‑L‑Leu)、cyclo‑(L‑Phe‑L‑Val)和ergosterol peroxide,且cyclo‑(L‑Val‑L‑Pro)的抑制活性最强。
本文采用OSMAC 策略,研究了不同培养条件和盐度对31 株石珊瑚共附生真菌次级代谢产物抗炎活性和化学多样性的影响。抗炎活性结果表明,多数菌株粗提物在10%的PDB 和糙米培养基条件下显示出了显著的抗炎活性,且糙米培养基的粗提物的抗炎活性明显大于PDB。化学多样性分析结果表明,菌株在PDB 和糙米培养基条件下次级代谢产物具有一定的差异性。推测可能的原因是糙米培养基与PDB 培养基营养源的不同,使得产生的次级代谢产物有所区别。糙米培养基以淀粉作为碳源,而PDB 培养基则均以葡萄糖为碳源,碳源为菌体生长与代谢提供了物质基础与能量,是培养基中重要的营养物质之一。碳源不同,菌株对碳源种类的利用程度不同,从而造成菌体产生的代谢产物种类和产量不同[21]。而在高盐条件下表现出更好的抗炎活性推测可能的原因是高盐条件下部分菌株细胞内编码抗炎活性组分的沉默基因被激活了,从而导致更多的抗炎活性的代谢产物能够被合成,使得粗提物的抗炎活性增加。
GNPS 是一个开放的串联质谱数据的数据库,能够实现分子网络的公共基础平台[22],能快速对LC‑MS/MS 生成的数据集进行分析[23-24]。马小翔等[25]对一株高产丁内酯I 的海洋来源土曲霉C23‑3进行化学调控,运用液相色谱-串联质谱法以及基于质谱的分子网络分析其次生代谢产物的产量和多样性发现,菌株C23‑3 经化学调控后的次生代谢产物产生了不同程度的变化。Wang 等[26]基于 MS/MS的分子网络来研究A. caelatus的化学成分,从而发现了新的二酮哌嗪二聚体和曲霉素。Freire 等[27]通过GNPS 对海绵Agelas dispar的甲醇部分进行研究,从中发现了新的溴吡咯衍生物。
FBMN 是一种基于高分辨质谱的改进GNPS 方法[28]。在本研究中,利用FBMN 进行了活性菌株次生代谢产物的注释,从数据库中匹配到了8 个抗炎化合物[29-36],且基于二级质谱相似性可发现多样化的衍生物,从而获得更多的抗炎活性物质。但从匹配结果可看出化合物的生物来源主要来自于植物和细菌,在珊瑚共附生真菌中尚未被发现,或尚未被GNPS 数据库所收录,说明珊瑚共附生真菌中的次级代谢产物值得进行深入研究。
4 结论
不同培养条件下石珊瑚共附生真菌次级代谢产物的抗炎活性和化学多样性表现出明显差异。其中5 株菌株在糙米培养基条件下表现出显著的抗炎活性,而菌株C3‑9 粗提物在低浓度条件下也表现出了显著的抑制NO 产生的作用。进一步的TLC 和FBMN 分析显示,菌株C3‑9 的次级代谢产物中含有丰富的抗炎活性化合物,这为后续抗炎活性化合物的深入研究提供了基础,对抗炎药物的研发具有一定的科学意义。
猜你喜欢
保健与生活(2023年9期)2023-05-19 21:07:36
江苏农业学报(2022年4期)2022-09-07 06:21:22
云南化工(2021年5期)2021-12-21 07:41:20
中国果业信息(2021年5期)2021-12-05 22:10:28
广东海洋大学学报(2015年3期)2015-12-22 10:05:30
华南农业大学学报(2015年5期)2015-12-04 03:04:38
中国洗涤用品工业(2015年5期)2015-02-28 19:02:26
食品工业科技(2014年13期)2014-03-11 18:16:55
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:20:57
食品研究与开发(2012年12期)2012-09-05 14:21:44