
IL-6-572C/G、IL-6-174G/C基因多态性与梅毒感染的相关性分析
2024-01-08 06:41张英李海燕王西珍
中国卫生标准管理 2023年23期
张英 李海燕 王西珍
梅毒是由梅毒螺旋体(treponema pallidum,TP)感染引起,是通过性传播、母婴传播的一种慢性性病,梅毒感染可以增加人类免疫缺陷病毒(human immunodeficiency virus,HIV)的感染和传播风险。根据全国性病病例报告,1989—2019 年以来,梅毒感染病例的报告率呈逐年增长趋势,加重了全球公共卫生负担[1-2]。梅毒发病机制尚未完全明确,有研究发现重组梅毒螺旋体蛋白Tp0768 通过激活NF-κB 通路和JNK 通路诱导白细胞介素1β(interleukin-1β,IL-1β)、IL-6 和白细胞介素8(interleukin-8,IL-8)的表达,与健康对照组相比,梅毒患者IL-6、IL-2、IL-10 血清表达水平均明显增高,在梅毒感染的起病、进展及预后中机体免疫应答、基因、遗传易感性在发挥关键作用[3-4]。以往研究证实IL-6 基因启动子区域的核苷酸基因多态性,可以导致基因转录和表达发生变化,影响个体对炎症及多种免疫性疾病的易感性[5-6],目前有关TP 感染与IL-6-572C/G、IL-6-174G/C 启动子区单核苷酸多态性(single nucleotidepolymorphism,SNP)未见报道,本研究旨在探讨IL-6(-572C/G、-174G/C)SNP 与梅毒感染的相关性。
1 资料与方法
1.1 一般资料
本研究纳入2020 年5 月—2021 年5 月至连云港市第二人民医院就诊的门诊、住院梅毒确诊患者80 例作为试验组,男性35例,女性45例;年龄32~82岁,平均(48.59±11.74)岁。根据中华人民共和国国家卫生和计划生育委员会行业标准(WS273—2018)中的《梅毒诊断》[7]进行梅毒感染疾病分期,其中轻度感染组为Ⅰ期、Ⅱ期共60 例,年龄32 ~77 岁,平均(47.58±12.78)岁;重度感染组为Ⅲ期共20 例,年龄32 ~84 岁,平均(49.02±11.36)岁。同时期至连云港市第二人民医院体检中心体检的健康人作为对照组,共120 例,包括男性63 例,女性57 例,年龄27 ~88 岁,平均(50.98±17.46)岁。两组性别、年龄比较,差异无统计学意义(P>0.05),具有可比性。本研究获得江苏省连云港市第二人民医院伦理委员会批准(批件号:2019-029),研究对象均签署了知情同意书。
纳入标准:(1)经化学发光法检测梅毒螺旋体抗体(treponema pallidum antibody,TP-Ab)阳性,甲苯胺红不加热血清(tolulized red unheated serum test,TRUST)试验验证阳性。(2)在连云港市第二人民医院诊治前3 个月内没有任何免疫调节剂、激素类以及抗菌药物的使用史。排除标准:(1)伴有其他性传播疾病者。(2)妊娠期及哺乳期妇女。
1.2 主要仪器及试剂
雅培全自动免疫分析仪i2000 仪器(德国Abbott 公司);数显混匀器WZR-H6000(春妙上海医疗器械有限公司);PCR 仪器(ABI Verti);核酸电泳及凝胶成像系统(北京六一生物科技);高速离心机(Thermo Fisher);TRUST 诊断试剂盒(上海荣盛药业有限公司);天根DNA 提取试剂盒(天根生化北京有限公司);M5 HiPer plus Taq HiFi PCR mix(北京聚合美生物科技有限公司)。
1.3 方法
1.3.1 引物设计
在NCBI数据库中下载从loci 22726410至loci 22727184的基因序列,包含IL-6基因 rs1800795(loci 22727026)、rs1800796 (Loc22726627)位点,用Snapgene 5.0.5软件设计扩增目的基因的P C R 引物,引物序列如下:上游:5'- GCCAACCTCCTCTAAGTG-3',下游: 5'-TCAGACATCTCCAGTCCTAT-3' ,委托杭州有康生物技术有限公司合成。
1.3.2 标本收集
研究对象清晨空腹6 ~8 h, 静脉采血收集EDTA-K2抗凝全血和血清各5 mL。血常规检测正常者,血清采用雅培全自动免疫分析仪i2000 仪器检测TP-Ab 浓度,阴性的标本纳入本次试验的健康对照组。TP-Ab 检测阳性者同时采用TRUST 试验进行验证,按照诊断试剂盒说明书操作,阳性者作为试验组,将收集的标本置于-80 ℃储存备用。
1.3.3 扩增目的基因
天根DNA 提取试剂盒提取500 μL 外周全血的DNA,严格按照操作说明书进行。DNA 产物浓度≥100 ng/μL 符合要求,在PCR 仪上反应扩增,扩增条件:95 ℃、5 min,1 个循环;95 ℃、30 s,55 ℃、30 s,72 ℃、30 s,30 个循环;72 ℃、5 min,1 个循环,16 ℃ 持续。扩增产物取5 μL 进行聚丙烯酰胺凝胶电泳,凝胶成像系统观察结果,收集出现目的条带的扩增产物,放至-80 ℃储存备用。
1.3.4 研究产物测序
样本的扩增产物委托杭州有康生物技术有限公司,样本双向测序,采用Sanger 双脱氧终止测序法,在ABI3730xl全自动测序仪上进行。3730 Data Collection v3.0 软件收集实验数据,FinchTV(1.4.0 版)软件分析IL-6-572C/G和IL-6-174G/C 位点基因多态性,分别采用rs1800796 和rs1800795 位点后10 个碱基,定位变异碱基的位置。
1.4 观察指标
试验组和对照组年龄、性别基础资料分布,目的基因测序结果,rs1800796(IL-6-572C/G)、rs1800795(IL-6-174G/C)等位基因频率、基因型以及不同遗传模式下试验组和对照组的基因型分布。
1.5 统计学方法
采用SPSS22.0 软件对数据进行统计学分析。计量资料采用(±s)表示,两样本均数的比较采用t检验;计数资料采用n(%)表示,基因型和等位基因频率采用直接计数法,两组间比较采用χ2检验;Logistic 回归模型分析IL-6 基因多态性和梅毒感染的相关性。P< 0.05 为差异有统计学意义
2 结果
2.1 对照组与试验组患者一般资料比较
对照组与试验组在性别、年龄等方面比较,差异无统计学意义(P>0.05);轻度感染组与重度感染组在性别、年龄等方面比较,差异无统计学意义(P>0.05),具有可比性,见表1、表2。
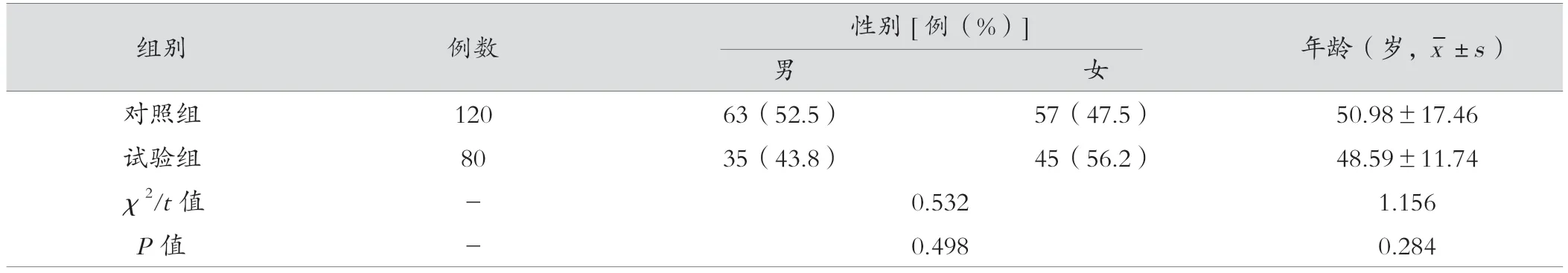
表1 对照组与试验组临床一般资料比较
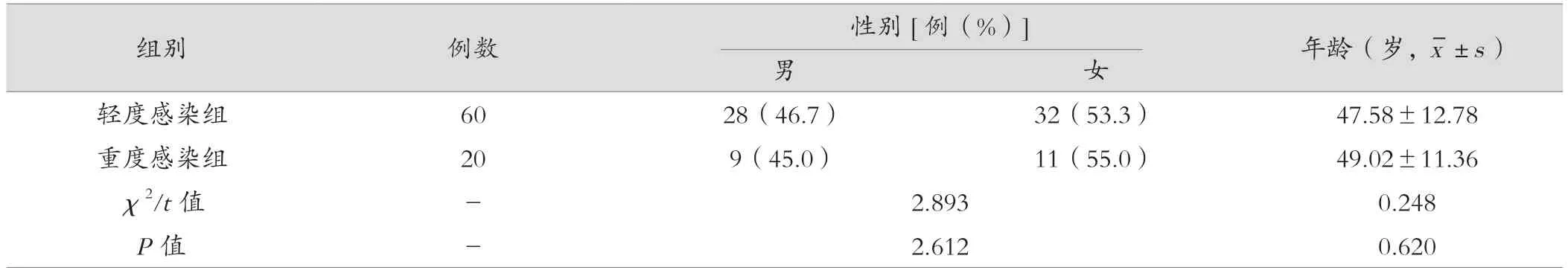
表2 重度感染组与轻度感染组临床一般资料比较
2.2 目的基因扩增及测序
IL-6 基因目的片段扩增产物有771 bp,在 750 bp DNA标记物的附近。连云港地区汉族人群IL-6-572C/G 等位基因有C 和G,基因型可见CC、CG 和GG 3 种;IL-6-174G/C 等位基因包括C、G,基因型可见GG 和CG 2 种,见图1。

图1 IL-6 启动子区-572 和-174 位点基因多态性测序结果
2.3 Logistic 回归分析IL-6 基因多态性和梅毒感染的相关性。
IL-6-174G/C 和-572C/G 多态性在试验和对照组中的分布频率符合Hardy-Weinberg 遗传平衡(P>0.05)。比较健康对照组和试验组IL-6-572C/G 基因型和等位基因频率,与CC 基因型相比,CG 和GG 两组与梅毒感染组差异均无统计学意义(P=O.428,OR=1.75,95%CI0.44 ~6.97;P=0.351,OR=1.94,95%CI0.48 ~7.86),等位基因C和G 频率在对照组和试验组无明显变化(P=0.817,OR=1.05,95%CI0.67 ~1.67),在显性遗传、隐性遗传、共显性遗传等不同的遗传模式下,基因型 CG+GG 和 CC,CC+CG 和GG,CC、CG 和 GG 都与梅毒感染不相关。IL-6-174C/G 基因型可见GG 与GC,等位基因G 和C,试验组和对照组可见分布差异无统计学意义(P=0.773,OR=0.66,95%CI0.04 ~10.77,P=0.874,OR=0.66,95%CI0.04 ~10.71),见表3。
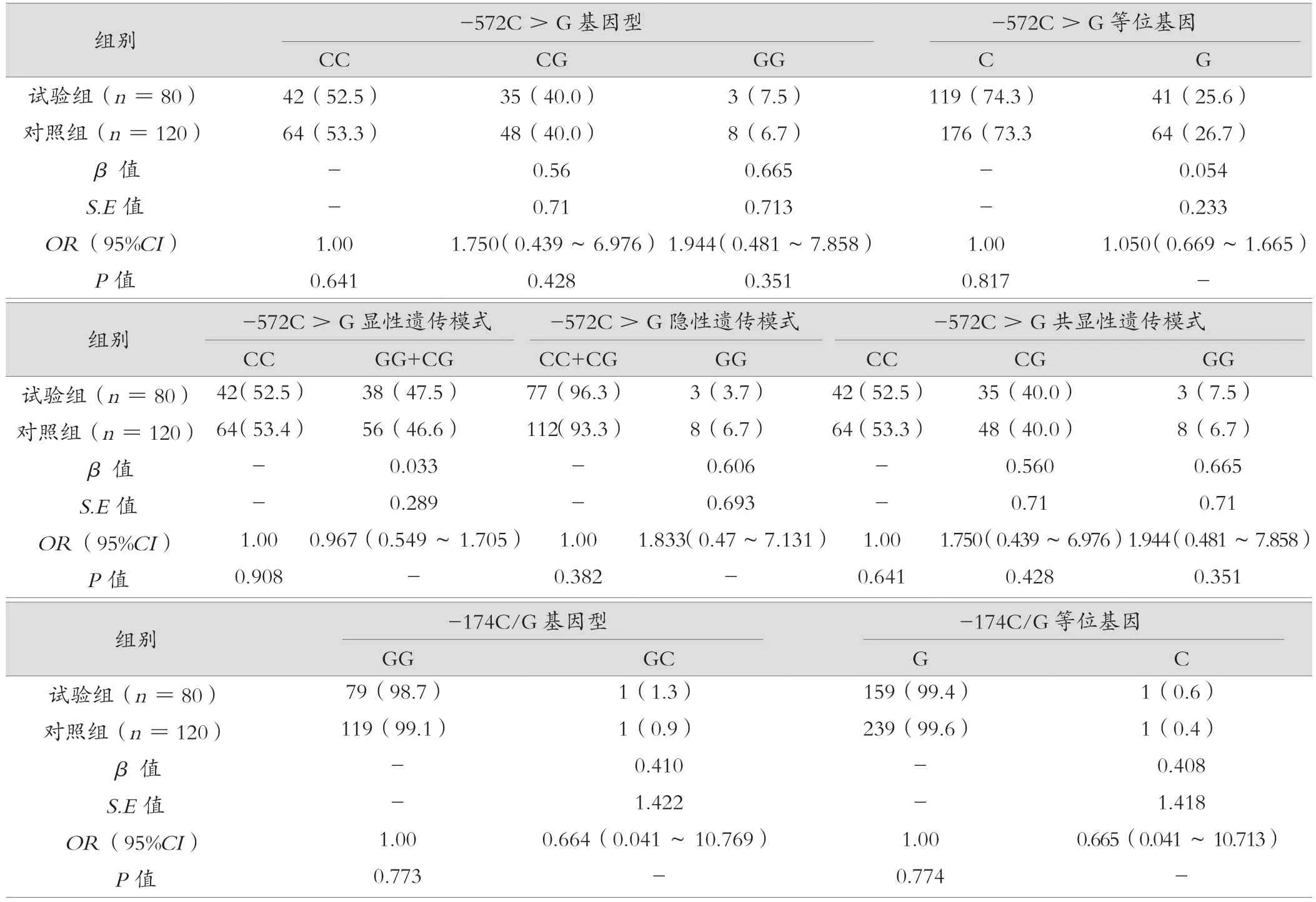
表3 IL-6-572C >G、-174C/G 等位基因频率、基因型以及不同遗传模式下的基因型分布 [ 例(%)]
3 讨论
梅毒感染病变可累及皮肤、眼、骨头、神经系统、心血管、肾脏、肝脏等多器官和脏器,引发严重的并发症和死亡率。逃避免疫监视是梅毒感染的重要发病机制,梅毒螺旋体入侵机体早期,首先T 淋巴细胞介导的细胞免疫被激活,并在清除病原体中发挥重要作用。随着病情进展到晚期,患者细胞免疫防御能力下降,Th1/Th2 细胞等免疫细胞及其分泌的细胞因子(TNF-α、IL-6 等)出现动态变化。梅毒螺旋体感染后增殖期,引发迟发性超敏反应,致敏CD4+T 细胞被激活,并在局部增殖并释放、吸引和激活巨噬细胞的细胞因子,而迟发性超敏反应无效时则转变为慢性病变。而免疫细胞分泌炎症因子能够调节机体的免疫应答,IL-6 为重要的免疫细胞因子。研究发现神经梅毒患者血清和脑脊液中 IL-6、IL-10 的含量明显高于对照组,但是血清中的 IL-2、IL-12和 IFN-γ 含量低于对照组[8]。课题组前期研究发现江苏省连云港地区梅毒患者IL-6 浓度比对照组显著升高,而且Ⅲ期患者IL-6 血清浓度明显高于Ⅱ期和I 期患者,说明IL-6 在梅毒感染的发生、发展过程中发挥了重要的作用。另有研究显示 IL-6 基因启动子区域中 SNP 对该基因的表达具有明显的影响[9]。近年来IL-6 多态性研究广泛,已经证实其SNP 和多种免疫性及感染性疾病相关性显著。笔者推测梅毒患者IL-6 启动子区域位点基因多态性,在梅毒感染过程中有一定的作用。
IL-6 主要由活化的单核细胞分泌,生理状态下其水平较低,在病原体刺激下,表达量显著升高。IL-6 表达受转录严格调控,IL-6 基因位于7p21 染色体上,全长5 kb,含有5 个外含子。IL-6 基因多态性主要表现在5'端启动子区域-572、-174 等位点的G/C,G/A 单核苷酸变异。余波等[10]发现急性脑梗合并肺部感染患者IL-6 基因-572G 基因频率较高,且基因型GG 频率较高,G 基因可能是易感基因。另有研究发现,脓毒血症患者IL-6 基因启动子区-572C/G 基因型CC 及等位基因C 频率明显低于对照组,基因型GG 及等位基因G 频率明显高于对照组,脓毒血症患者血清IL-6水平显著高于健康组[11]。本研究发现IL-6 启动子区-572 等位基因有C 和G,基因型有CC、CG 和GG3 种,IL-6 启动子区-174 等位基因有C、G,基因型有GG 和CG 两种,与上述研究发现一致。但是课题组进一步研究发现梅毒患者IL-6-174 基因型GG、CG 以及等位基因C 和G 频率与健康人群中的分布并差异无统计学意义(P> 0.05),IL-6-572 3 种基因型和等位基因频率和健康者同样差异无统计学意义(P> 0.05),同时在显性遗传模式、隐性遗传模式、共显性遗传模式下,试验组和健康对照组之间均没有关联,这提示IL-6 启动子区域-572 和-174 位点多态性与梅毒感染易感性无关。既往研究表明,梅毒感染与多个遗传位点的SNP 联合作用有关,即可能存在梅毒感染患病相关的基因间存在SNP 位点间的交互调控作用,且相关基因位点间的单体型也可能增大梅毒感染的患病风险[12-14]。这也进一步提示梅毒感染是由先天性免疫、遗传易感性等共同作用的结果。
综上所述,本研究首次证实IL-6-572C/G 和IL-6-174G/C 基因多态性与梅毒易感性没有关联,为后续IL-6基因多态性与梅毒感染的位点研究提供选择依据。同时也存在一定的局限性:是小样本单中心研究,仅分析了IL-6启动子区2 个位点多态性,需要更大样本多中心以及涉及IL-6 5'端和3'端多个位点的基因多态性的综合研究,明确IL-6 基因SNP 与梅毒感染的相关性。文章中体现了中华人民共和国国家卫生和计划生育委员会行业标准(WS273—2018)中的梅毒诊断执行标准[7]。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
智慧健康(2021年17期)2021-07-30
现代检验医学杂志(2016年5期)2016-08-20
西南农业学报(2016年6期)2016-04-16
西南军医(2016年4期)2016-01-23
法医学杂志(2015年4期)2016-01-06
法医学杂志(2015年4期)2016-01-06
西南医科大学学报(2015年1期)2015-08-22
河南医学研究(2014年7期)2014-02-27
现代检验医学杂志(2014年1期)2014-02-06
