
过敏性紫癜发病机制的研究进展
2023-12-28 04:36任瑞华罗沙沙钟小明
赣南医学院学报 2023年10期
任瑞华,罗沙沙,钟小明
(1. 赣南医学院第一临床医学院;2. 赣南医学院第一附属医院儿科;3. 赣州市儿童白血病肿瘤免疫治疗药物研发重点实验室,江西 赣州 341000)
过敏性紫癜(Henoch-Schönlein purpura, HSP)又称IgA 血管炎(Immunoglobin A vasculitis, IgAV),是以小血管炎为主要病变的系统性血管炎。病理变化主要以免疫球蛋白A(Immunoglobin A, IgA)沉积、补体因子和炎症浸润为特征,并伴有血管炎症。HSP 是儿童最常见的血管炎,临床特点主要为血小板不减少性皮肤紫癜,常伴关节肿痛、腹痛、便血、血尿和蛋白尿[1]。目前关于HSP的发病机制存在两种假说:基于IgA 肾病的四重打击学说和新多重打击学说[2]。但这两种假说均未阐明免疫细胞和免疫分子的相互作用。近年来基因突变、免疫细胞及免疫分子研究在HSP 发病机制中取得了一些成果,本文从外部因素、遗传易感性及免疫微环境的改变三个方面进行综述。
1 外部因素
1.1 感染目前研究证实大多数HSP 发病前存在感染。2020 年的一项研究表明,约50.92%的患儿存在感染,其中链球菌是最常见的感染源[3]。2022年LIU C等[4]研究发现,68.75%患儿有感染史,肺炎支原体是最常见的感染源(45.77%),其次是A 组溶血性链球菌(28.87%),除此之外,结核杆菌、EB 病毒、幽门螺杆菌、甲型流感病毒、乙型肝炎病毒也参与其中。这两项研究表明,不同时间和空间的HSP患儿感染存在不同的病原体疾病谱。另外,新近报道的2019-冠状病毒也能诱发HSP,以成人多见[5]。
1.2 非感染HSP的病因也与非感染因素相关,如食物过敏(蛋类、乳类、豆类等)、疫苗接种(流感疫苗、乙肝疫苗等)、药物(阿司匹林、抗生素等)、恶性病变等[1]。既往研究认为控制饮食对HSP患儿的治疗是非常重要的,通过禁止摄入所有动物蛋白和大部分植物蛋白来治疗和预防HSP。消除饮食中的过敏原和疑似食物配方是治疗食物过敏的主要目的,通常指消除食物中的蛋白质,这是治疗食物过敏症的基础。但尹东豪等[6]研究结果表明,动物蛋白饮食与HSP 复发或皮疹反复无明显相关性。WANG L等[7]将氨基酸的配方奶粉添加到HSP 患儿的饮食中,然后随机分为饮食限制组和饮食指导组,结果发现,饮食指导可以提高HSP 患儿的营养素和蛋白质摄入量,减少皮疹复发和并发症发生率。一些学者发现,20.5%的HSP 儿童在发病前食用特定食物,食物中90%的过敏原是蛋白质,这些蛋白质具有糖蛋白的抗原特异性,能够承受食物加工和加热而不受损害,不易在肠道内消化[8]。由于HSP 高发人群是处于生长发育阶段的儿童,长期禁止摄入蛋白可能会影响患儿的营养状态。因此,对HSP 患儿制定合理的饮食结构是非常重要的。
2 遗传易感性
遗传因素在HSP 发病机制中的影响越来越受到关注。有研究[9]认为,不同种族的人群发病率不同,其中亚洲人发病率最高(24.0/10 万),黑种人发病率最低(6.2/10万)。但由于流行病学调查研究方法不一致,该结论仍存在争议,需进一步采用多中心、统一的方法进行研究,以确定种族差异是否为HSP的危险因素。
近年来发现多个基因位点与HSP 相关。研究证实,HLA-B35和HLA-DRB1*01等位基因与HSP易感性有关,特别是HLA-DRB1*0103与HSP易感性升高密切相关[9]。ZHANG X 等[10]研究表明,血管紧张素转换酶D等位基因可能是HSP的危险因素。酶编码基因的多态性也可能影响Gd-IgA1 合成。NAGPA基因内含子的遗传变异与HSP 有很强的相关性,是迄今为止最强的非HLA 信号,其作用机制是NAGPA基因参与甘露糖6-磷酸的形成,并编码甘露糖6-磷酸形成的催化酶,酶活性可能与自身抗原的生成有关,进而诱发自身免疫疾病[11]。凝血酶因子V 基因突变也可能与临床症状有关[12]。也有研究[13]发现,地中海热基因(MEFV)突变与地中海人群中的HSP患儿存在关联,MEFV突变导致先天免疫系统炎症改变,从而增加血管炎的易感性。LEE M 等[14]研究发现,在小儿IgA 肾病和HSP 的1 号染色体上有一个新的易感基因——单核苷酸(rs9428555),该基因变异可能会影响维生素D相关通路和肾功能。
3 免疫微环境的改变
人体免疫系统通过免疫器官、免疫组织、免疫细胞和免疫分子执行免疫功能,免疫细胞和免疫分子在HSP患儿的免疫微环境中发挥重要作用。
3.1 免疫细胞
3.1.1 T 淋巴细胞T 淋巴细胞依据CD 分子可分为CD4+T 淋巴细胞和CD8+T 淋巴细胞,CD4+T 淋巴细胞与HSP 发病相关得到普遍认可,CD4+T 淋巴细胞可分化为不同细胞亚型,其中辅助性T 淋巴细胞(Helper T cell,Th)和调节性T 淋巴细胞(Regulatory T cell,Treg)具有重要作用。既往研究证实,HSP 患儿体内存在Th1细胞和Th2细胞活性失衡:Th1细胞功能降低,Th2 细胞功能优势活化,Th1 细胞的分化在白细胞介素12(Interleukin-12,IL-12)和干扰素γ(Interferon-gamma,IFN-γ)的作用下,使Th1 细胞特异性转录调节因子T-bet上调,增强IL-12受体表达,同时抑制GATA-3 的表达;抗原信号和IL-4 促进GATA-3 的表达,并阻止IL-12 受体表达,从而诱导Th2 细胞分化,同时抑制Th1 细胞分化。一项Meta分析表明,Th17细胞和Treg 细胞失衡与HSP 疾病活动密切相关,当机体受到抗原刺激后,TGF-β、IL-21诱导Th17 细胞分化并分泌IL-17、IL-6 等细胞因子,IL-6 和IL-21 在异常的免疫微环境中可能导致Treg细胞向Th17 细胞转化,两者失衡可诱导B 细胞产生过量IgA抗体沉积于小血管壁而促使HSP发病[15]。
随着HSP 发病机制的深入研究,其他T 淋巴细胞亚群也被关注,如滤泡辅助性T(Follicular T helper cells, Tfh)淋巴细胞和γδ T 淋巴细胞。研究发现,Tfh 淋巴细胞是抗体产生的真正辅助者,Bcl-6 可能调控初始T 淋巴细胞分化CXCR5+Tfh 淋巴细胞分泌IL-21,IL-21 可诱导B 淋巴细胞分化为浆细胞,进而分泌大量的IgM、IgG、IgA[16]。近年来,γδ T 淋巴细胞的作用也逐渐显现。郭佳琦等[17]研究表明,HSP患儿存在γδ T 淋巴细胞免疫失调和凋亡失调,γδ T淋巴细胞通过Fas/FasL、TNF 相关凋亡诱导配体及穿孔素/颗粒酶等途径发挥细胞毒性作用,还可以分泌IL-17、IFN-γ 激活JAK-STAT 或Ras 途径等通路,调节淋巴细胞比例而改变免疫微环境。
3.1.2 B 淋巴细胞B 淋巴细胞不但可以分化为浆细胞,通过浆细胞产生抗体介导体液免疫应答,而且也具有抗原提呈及免疫调节作用。目前,浆细胞产生的IgA 和B 淋巴细胞分泌的具有协同刺激作用的细胞因子参与HSP 患儿的发病得到普遍认可。研究发现,具有免疫调节作用的调节性B 细胞经抗原刺激后产生IL-10 和TGF-β,进一步影响Th17 细胞和Treg 细胞的平衡,参与HSP 和紫癜性肾炎的发生发展[18]。然而,ZHANG N 等[19]研究发现,HSP 患儿总B 细胞的百分比和绝对数量明显增高,而浆细胞的百分比显著降低,未发现紫癜性肾炎的发病机制与B10细胞产生的IL-10的减少有关。
3.1.3 其他免疫细胞NK 细胞在对抗肿瘤细胞和病原体感染细胞中起重要作用。NK 细胞调节紊乱可能导致清除外来抗原障碍,产生免疫损伤。HSP 患儿NK 细胞降低,异常增高的IgA 与NK 细胞表面的Fc受体结合,抑制NK细胞介导细胞毒作用,并且异常的糖酵解途径抑制NK 细胞杀伤作用,这些免疫微环境的变化提示NK 细胞免疫监测受损[20]。此外,既往研究发现,在HSP 患儿外周血单个核细胞中表达的细胞表面信号转导跨膜受体(Toll 样受体)异常活化,异常活化的Toll 样受体可能是通过上调Tfh、Th2 细胞的免疫应答从而诱导IL-21、IL-4、IL-6等炎症细胞因子释放来参与HSP的发病。
3.2 免疫分子
3.2.1 IgA 及Gd-IgA1 的产生IgA 是一种Y 型的免疫球蛋白,可分为IgA1 和IgA2,IgA1 约占90%。IgA1的铰链区通常包含3~6个O-聚糖位点,是糖基化的核心结构,当这些位点结合异常或过早酸化时,将导致铰链区中半乳糖糖基化或酸化的糖基减少,异常的糖基化可形成Gg-IgA1。目前,Gd-IgA1及其循环免疫复合物在肾小球系膜区沉积是公认的IgA 肾病可能发病机制之一。有研究[21]表明,紫癜性肾炎和IgA 肾病患儿血清Gd-IgA1 水平高于HSP 患儿和健康对照组,而HSP 患儿血清Gd-IgA1水平与健康对照组无差异,结果提示Gd-IgA1 水平与肾损害相关。
3.2.2 补体的激活HSP 患儿皮肤活检中C3沉积提示补体的激活。研究表明,HSP 患儿血清中C4d、MBL、MASP1/3 沉积、P 因子与非HSP 患儿相比明显增多,MBL 可被各种病原体(包括细菌、酵母、真菌和病毒)上的末端甘露糖和N-乙酰氨基葡萄糖激活,高度糖基化的IgA1 可能是MBL 更有效的激活剂,凝集素和旁路途径的激活参与HSP 患者的组织损伤,并诱导细胞因子表达上调及招募炎症细胞[22]。但也不能排除广泛的血管壁损伤引起继发性经典途径激活。
3.2.3 细胞因子细胞因子是在细胞间发挥相互调控作用的一类小分子可溶性蛋白质,通过结合相应受体调节细胞生长、分化和免疫效应。除IL-6、IL-12、IL-17、IL-21 及IFN-γ 等细胞因子外,还值得关注的是HSP 患儿血管损伤中涉及到的其他细胞因子。IL-8 是中性粒细胞重要的趋化因子,中性粒细胞是HSP 患儿皮肤和胃肠道活检中炎症浸润的主要细胞。HSP 患儿中的IgA 沉积物可能通过Fcα受体Ⅰ激活中性粒细胞,从而导致中性粒细胞胞外陷阱的释放及TNF-α 的产生,Fcα 受体Ⅰ激活后会启动IL-8 等细胞因子和趋化因子级联反应而导致肾脏损伤[23]。此外,在2007年才被正式命名主要由调节细胞(如调节性T 淋巴细胞,调节性B 淋巴细胞等)分泌的IL-35,通过抑制TNF-α及NF-κβ 通路,从而抑制趋化因子和中性粒细胞迁移[24]。
4 总结与展望
虽然HSP 病因及发病机制的研究在基因学及免疫学方面取得一些进展,可能的发病机制如图1所示,但仍需进一步探索易感基因对相关通路的影响及免疫细胞与免疫分子的相互作用。由于免疫学的理论发展和技术提高,免疫学知识已广泛用于疾病的预防、诊断和治疗。特别值得一提的是近年来发现维生素D 具有免疫调节的作用,其在HSP 发病中的作用和治疗价值也越来越受到关注。
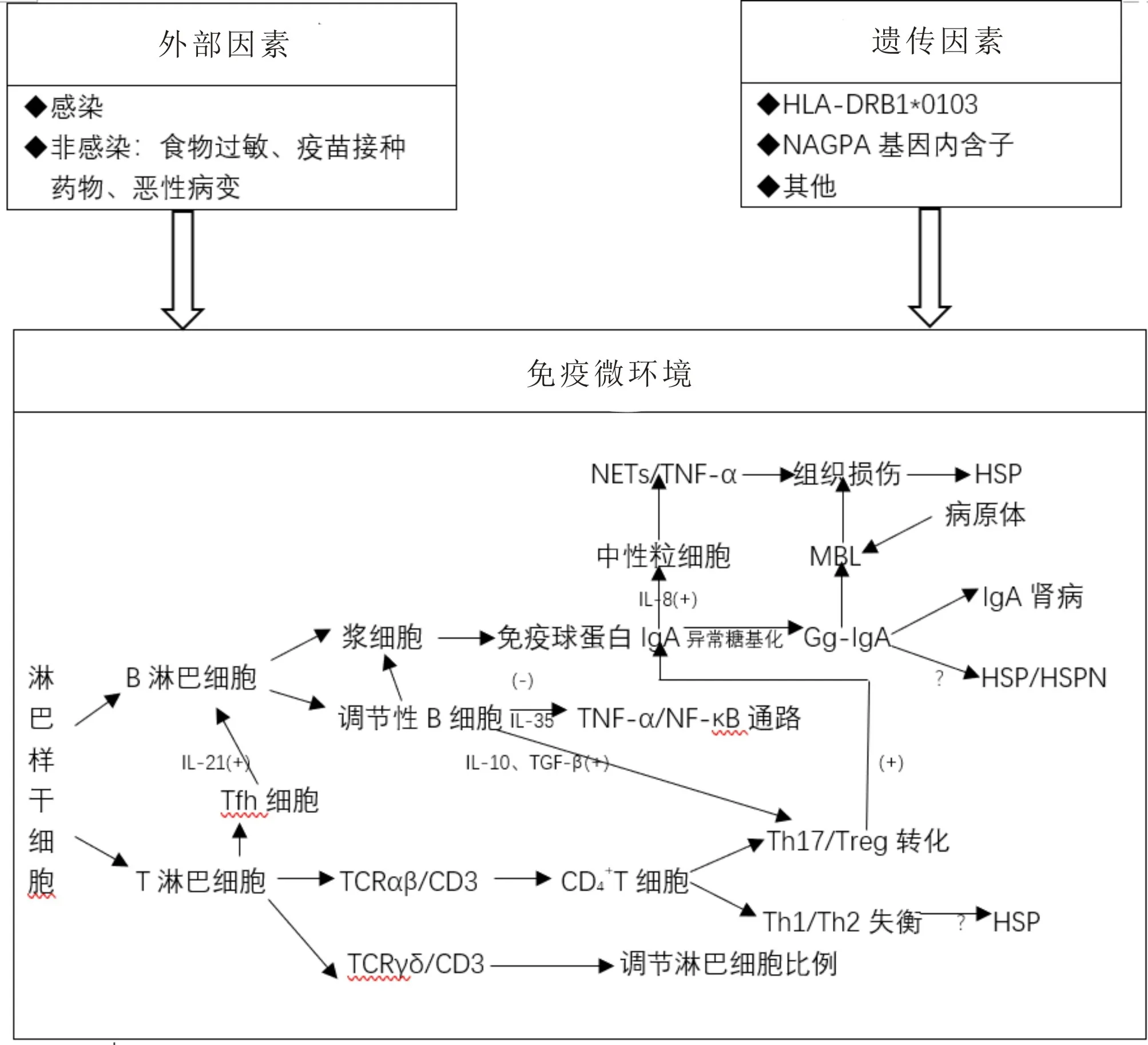
图1 HSP可能的发病机制作用图
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
现代临床医学(2021年4期)2021-07-31
哈尔滨医药(2016年3期)2016-12-01
中西医结合心脑血管病杂志(2016年20期)2016-03-01
癌变·畸变·突变(2016年3期)2016-02-27
医学研究杂志(2015年12期)2015-06-10
现代检验医学杂志(2015年6期)2015-02-06
现代检验医学杂志(2014年1期)2014-02-06
西南军医(2014年1期)2014-02-03
中国医学科学院学报(2013年2期)2013-03-11
