
三峡库区消落带草本植物根际细菌群落季节变化特征及功能预测
2023-12-26 02:41张丽苗袁中勋李昌晓
生态学报 2023年23期
谭 雪,董 智,张丽苗,袁中勋,李昌晓,*
1 西南大学生命科学学院,三峡库区生态环境教育部重点实验室,三峡库区植物生态与资源重点实验室,重庆 400715
2 西南山地生态循环农业国家级培育基地,重庆 400715
三峡大坝正式竣工后,采用了冬蓄夏排的水位调度方式,形成了水位落差达30 m,面积近400 km2的消落带[1]。在此特殊的水文条件下,消落带原有的陆生植被大量减少,环境污染、土地退化等问题日益严重,生物多样性明显降低[2]。为了改变这一现状,研究者们筛选出了一些物种用于消落带的植被重构,其中两种C4植物狗牙根(Cynodondactylon)[3]和牛鞭草(Hemarthriaaltissima)[4]被确定为消落区植被修复的优良适生草本植物种类。狗牙根属禾本科多年生草本植物,在消落区广泛分布,其根系发达、繁殖能力强,对水淹和干旱有较强的适应能力[5];牛鞭草是一种在湿润和干旱环境都能生长的植物[6—7],在河岸带植被修复中得到广泛的应用[8]。目前有关狗牙根、牛鞭草的研究主要集中在水淹胁迫下植物土壤养分变化、根系酶活[9]、光合特性[10]等方面,但对两种适生草本的土壤微生物,特别是三峡库区消落带水文变化条件下根际微生物的季节动态及功能特征方面报道较少,这使得难以准确深入地理解和认识两种适生草本植物在三峡消落带特定环境下的根际微生态特性。
“根际”这一概念系1901年德国微生物学家 Hiltner首次提出,根际为根系周围、受根系生长影响的土体[11]。根际微生物是受植物影响最大的土壤微生物群体[12],一方面植物根系分泌物提供能源供微生物所需,另一方面根际微生物还能够推动土壤物质和能量的流动,促进植株生长[13]。因此,为了从根际微生态的视角探究适生草本如何适应三峡库区消落带这一特殊生境,本研究选取位于三峡库区的重庆忠县汝溪河流域消落带修复示范基地为研究区域,选取经8 a淹水周期,仍然保持良好生长特性的牛鞭草、狗牙根草本植被根际土壤作为实验材料,采用illumina hiseq测序技术,对两种适生草本植物根际土壤细菌的季节动态分布格局及其驱动因子进行研究,同时结合基于未观测状态重建的群落系统发育研究(PICRUSt2)功能预测[14]方法,对两种适生草本植物根际土壤细菌功能类群进行了预测分析,为深入探究消落带根际土壤细菌群落的变化规律,了解三峡库区消落带植被修复与重建对土壤微生物菌群的影响提供参考依据。
1 研究地点与方法
1.1 研究地点概况
研究区域位于重庆市忠县汝溪河流域(107°32′—108°14′E,30°03′—30°35′N),处于重庆市中部,远离主城区,水位变化与三峡大坝水位调度方式一致,区域属亚热带东南季风气候,季节分明,降雨充沛,全年温度≥10℃,年积温为5787℃,年均气温18.2℃,无霜期341 d,日照时数1327.5 h,日照率29%,太阳总辐射能3.5×105J/cm2,年降雨量1200 mm,相对湿度80%。该区域土壤主要为石灰性紫色土,土壤熟化程度较低,水土流失、土层侵蚀现象严重。
课题组于2012年3—4月在重庆忠县石宝镇汝溪河流域的消落带构建植被修复示范基地,基地面积13.3 hm2,土壤质地、肥力状况基本一致。研究区域内,栽植有大量的牛鞭草、狗牙根多年生草本植物,所选岸坡样地平均坡度为26°,岸坡光照辐射强度大致相同,土壤雨热条件基本一致,植被覆盖率为100%。经过连续8 a的淹水周期,植物生长状况良好。
1.2 样品采集与土壤化学性质测定
根据水位在忠县石宝镇汝溪河流域的变化及植被生长情况,本实验分别于2020年5月(春季)、7月(夏季)、9月(秋季)进行野外取样调查,其采样时间分别对应消落带退水初期、草本植被生长旺盛期及消落带淹水前期,而冬季适生草本植被处于水淹状态,无法完成土壤取样。
实验材料为经过8 a反复水淹,生长状况良好的多年生草本植物狗牙根、牛鞭草根际土壤。为了确保样地不受到消落带上部可能的人为干扰以及下部的水文干扰带来的影响,采样时选择在海拔160—170 m中间地带划定S形取样带。在此海拔区间,两种草本生长状况良好,植被纯度及覆盖率高,在S形样带内划定3个1 m×1 m取样样方,每个样方间距十米以上。在每个样方内按梅花形设置5个(15 cm×15 cm×20 cm)重复取样点,去除表层地被物后,采集0—20 cm土层的原状土壤样品,然后将一个样方内的根际土壤混合。根际土壤的采样方法参考Riley和Barberd的抖落法,除去根系附近的石子及动植物残体,抖落根系表面的粒土和杂质,采集附着于根际表面4 mm以内的土壤作为根际土样[15]。每个季节采集土壤混合样本6个,3个季节共采集根际土壤混合样本18个。
采集的土壤样品分为两部分,一部分储存于-80℃用于细菌群落测定,一部分自然风干,研磨过筛(1 mm和0.25 mm),用于其他土壤化学性质的测定。土壤含水量采用烘干法[16]测定,土壤pH值采用电极电位法测定(1∶2.5土水比浸提液),土壤全碳(TC)、全氮(TN)含量测定采用元素分析仪(Elementar Vario EL,德国),土壤有机质(OM)测定采用水合热重铬酸钾-硫酸-比色法[17];土壤全磷(TP)、速效磷(AP)含量测定采用钼锑抗比色法;全钾(TK)、速效钾(AK)含量测定采用电感耦合等离子体发射光谱法[18](ICP-OES, Thermo Fisher iCAP 6300,英国),土壤硝态氮、铵态氮采用全自动间断化学分析仪(CleverChem,德国)测定。
1.3 基于高通量测序的细菌群落结构分析
本研究中根际土壤总DNA的提取采用DNeasy® PowerSoil® Kit(QIAGEN)试剂盒,操作步骤按内置使用说明书进行,提取的总DNA经过1.2%琼脂糖凝胶电泳鉴定,然后置于-20℃保存备用。采用通用引物细菌16S V4—V5 区进行聚合酶链式反应(PCR)扩增,引物序列为515F(5-GTGCCAGCMGCCGCGG-3)和907R(5-CCGTCAATTCMTTTRAGTTT-3),每个样品进行三次重复扩增,然后混合同一样品扩增产物;PCR产物切胶纯化,Qubit荧光定量仪定量之后,等摩尔混合,建立测序文库,采用 illumina hiseq 进行测序。
测序获得的数据采用微生物生态学定量研究平台研究平台(QIIME)进行进一步的生物信息学分析[19],采用FLASH算法拼接双端序列,同时,为获得高质量序列,根据质量分数Q20对序列进行质控,使用UPARSE算法[20]将得到的高质量序列进行聚类OTU,在聚类过程中严格去除嵌合体序列,使用核糖体数据库项目(RDP)数据库对可操作分类单元(OTU)代表序列进行物种注释。
土壤细菌α多样性由Chao1指数、Simpson指数、Shannon指数、Observed-Otus和PD-whole-tree指数进行表征,Chao1指数反映了种群的丰富度,Shannon和Simpson指数反映了物种多样性,Observed-Otus表示注释的OTU数目,通过对相似度高于97%的序列进行Cluster聚类得到。PD-whole-tree指数是基于系统发育树进行计算的,其过程是将样品的代表序列发育树枝长加和,得到多样性指数,其值越大,群落多样性越高。
相关计算公式为:
其中,S1为观察到的OTU目;ni=含有i条序列的OTU数目;N=所有序列数。
1.4 统计分析
采用单因素方差分析(ANOVA)对不同季节根际土壤理化特性及细菌群落功能丰度进行分析,采用冗余分析(RDA)分析微生物群落与土壤理化性质之间的关系,用PICRUSt2[21]对细菌群落进行功能预测分析,下游数据分析在QIIME、R软件、SPSS 18.0中进行,采用R 4.0.3、Canoco 5、Excel 2018作图。
2 结果与分析
2.1 植物根际土壤理化性质
研究区域土壤温度在不同季节间存在显著差异(P<0.05),其中夏季温度最高(表1)。如表1所示,根际土壤含水量范围为16.84%—18.86%,季节间差异不显著。狗牙根和牛鞭草两种适生草本根际土壤pH值范围为6.89—7.37,且夏秋两季pH值显著高于春季(P<0.05)。与此同时,根际土壤全碳、全氮含量夏季显著高于秋季(P<0.05);土壤有机质含量秋季显著低于春夏两季(P<0.05);土壤铵态氮含量夏季显著大于春季(P<0.05);土壤速效钾随季节逐渐升高,且秋季显著大于春季(P<0.05)。

表1 植物根际土壤理化性质的季节动态变化
2.2 根际土壤微生物群落α多样性指数
经过对3个季节两种适生草本根际土壤样品进行16S rRNA测序及物种注释,共得到 24409个OTU,其中春季、夏季、秋季各有16499、17111、19559个OTU。3个季节共有的OTU数目为11486,约占总数的47.1%;春季和夏季独有的OTU数目较为相似,分别占OTU总数的8.1%和8%,而秋季独有的OTU数目最多,约占全部的13.1%。可见,秋季草本根际土壤中细菌物种数最多,春季和夏季较少(图1)。

图1 不同季节根际土壤细菌OTUs Venn图
如表2所示,研究区域狗牙根与牛鞭草两种草本的根际土壤细菌群落α多样性指数存在显著的季节性差异,在春、夏、秋三次取样中,Chao1、Shannon指数、Observed-Otus均表现为秋季显著大于春夏两季(P<0.05)、PD-whole-tree指数在秋季显著大于春季(P<0.05)。
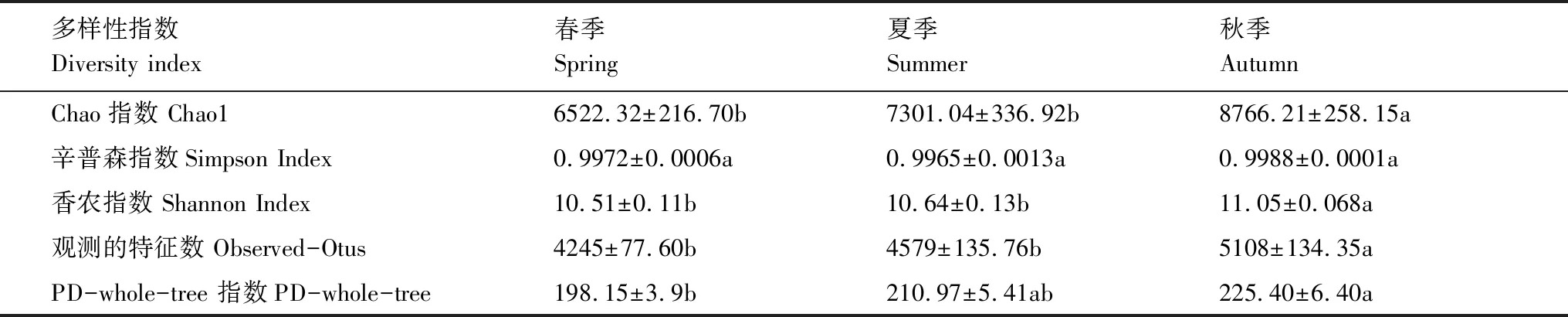
表2 根际土壤细菌群落α多样性指数的季节变化
2.3 根际土壤微生物群落结构组成
通过对3个季节植物根际土壤样本的高通量测序,共获得1882871条高质量序列,其中最少序列为27715条,最多序列为233350条,平均104604条。这些序列的99.87%可以分类到不同的细菌门,0.09%与数据库进行对比后分类到古菌门类。
所有样本共检测到细菌60门、151纲、245目、268科和403属。图2和图3分别展示了不同季节各样地门(Phylum)和纲(Class)分类水平的细菌群落结构。由图2可知,3个季节适生草本根际土壤中检测到的主要门有变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、浮霉菌门(Planctomycetes)、硝化螺旋菌门(Nitrospirae)、厚壁菌门(Firmicutes)、芽单胞菌门(Gemmatimonadetes)和未知菌门(WS3)。其中,变形菌门、酸杆菌门、拟杆菌门是根际土壤中占主导地位的细菌门类,约占细菌总数的70.46%。各个季节根际细菌群落在组成上较为相似,其中仅放线菌门的相对丰度在春季显著低于秋季(P<0.05),芽孢杆菌门的相对丰度在夏季显著低于秋季(P<0.05),其余各门在季节间分布较均匀,差异不显著。纲分类层次(图3)的统计表明,样品中优势菌纲主要有α-变形杆菌纲(Alphaproteobacteria)、β-变形菌纲(Betaproteobacteria)、酸杆菌纲(Acidobacteria-6)、δ 变形菌纲(Deltaproteobacteria)、γ变形菌纲(Gammaproteobacteria),约占全部样品的51.41%,其中,仅α-变形杆菌纲相对丰度在春季显著高于夏季(P<0.05)。
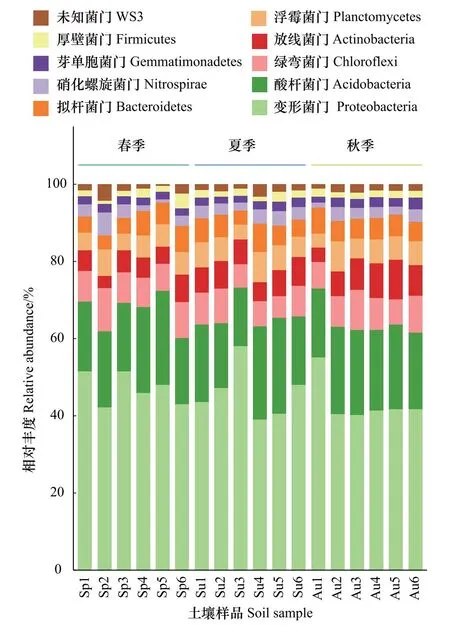
图2 不同季节根际土壤样品中优势细菌门的相对丰度
2.4 根际土壤细菌群落与环境因子相关性
图4展示了各季节对细菌群落结构贡献率较大的理化因子与主要细菌门类的相关关系,其结果表明,春季适生草本根际土壤中94.26%的细菌群落变化是由土壤理化性质引起的,此外,RDA图的轴1解释了81.04%,轴2进一步解释了13.22%,对根际细菌群落结构贡献率较大的有土壤氧化还原电位(ORP)和TP;此外,变形菌门、硝化螺旋菌门与土壤pH值和含水量呈显著正相关关系,与土壤ORP呈显著负相关关系。

图4 不同季节环境因素对土壤细菌群落结构特征影响的冗余分析
夏季第一物种轴与第二物种轴分别可以解释78.17%、17.36%的物种数据变化,累计解释了95.53%的物种-环境关系,对夏季根际细菌群落贡献较大的有土壤AK和TP,其中土壤AK对细菌群落有显著影响;此外,土壤AK、TK、TP和TN与放线菌门、变形菌门呈正相关关系,与硝化螺旋菌门、厚壁菌门和绿弯菌门呈负相关关系。

2.5 根际土壤细菌群落功能预测
通过对京都基因与基因组百科全书(KEGG)数据库进行比对,共获得6类一级生物代谢通路功能(图5):代谢、遗传信息处理、环境信息处理、细胞过程、有机系统、人类疾病。在分布于6个代谢途径的细菌群落中,共鉴定了45个二级KEGG数据库中功能直系同源物(KO)组。其中,代谢、人类疾病、遗传信息处理和环境信息处理途径最为普遍,分别占27.47%、19.4%、18.23%和14.06%。

图5 不同季节根际细菌预测功能基因一级功能丰度图
对预测基因二级功能层丰度进行分析(图6),发现根际细菌群落主要涉及膜运输(10.69%)、翻译(9.69%)、氨基酸代谢(8.84%)、碳水化合物代谢(7.09%)、辅助因子和维生素的代谢(5.78%)、能量代谢(Energy metabolism)(5.38%)、复制和修复(5.14%)等45个二级功能,其中14个功能基因的丰度表现出显著的季节性差异(图6)。
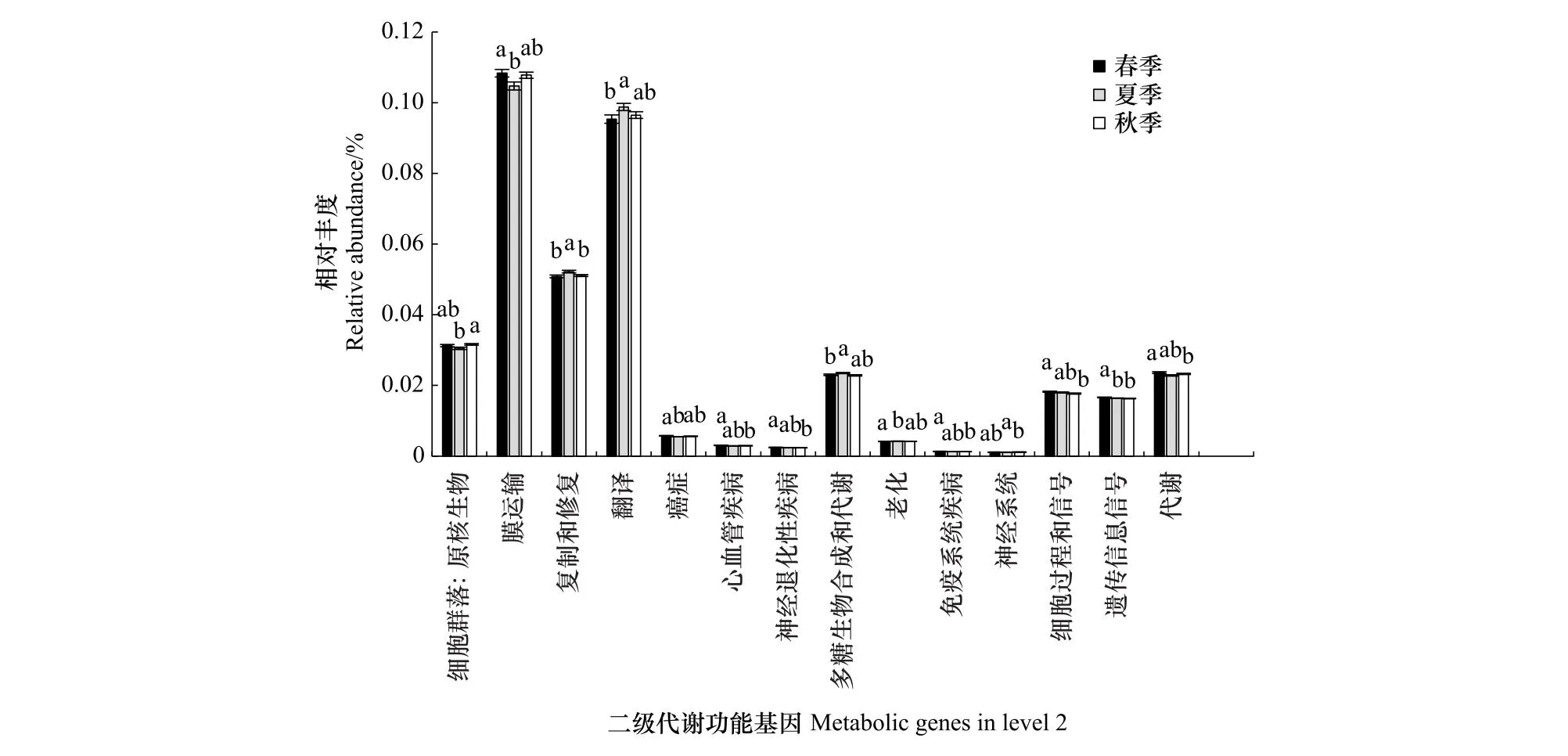
图6 不同季节根际细菌预测功能基因二级功能丰度图
3 讨论
3.1 不同季节适生草本根际细菌群落结构特征
三峡库区消落带适生草本根际土壤细菌优势门为变形菌门、酸杆菌门、拟杆菌门、绿弯菌门,这与大量研究土壤细菌的文献结果较为一致[22—23]。本研究中,3个季节细菌群落结构在门水平上组成较为相似,说明适生草本经过8 a的反复水淹,形成了较为稳定的根际细菌群落环境。变形菌门是根际细菌群落中丰度最大的一个门,现有的研究表明,变形菌门广泛分布于各种生境,如湖泊湿地[24]、盐渍化土壤[25]、温带针叶林土壤[26]等,其包括的α-变形杆菌纲、β-变形菌纲、δ变形菌纲和γ变形菌纲在纲水平所占比例也超过了41%。α-变形杆菌纲包含有植物共生菌(根瘤菌)等固氮细菌[27],春季α-变形杆菌纲的相对丰度显著高于秋季,对提高适生草本在退水初期固氮能力,进而促进植物快速生长具有重要意义。
放线菌门多数严格好氧,其分枝状的菌丝体能够产生各种胞外水解酶,降解土壤中各种难溶性有机物质以供细胞代谢所需的各种营养,在自然界物质循环、土壤改良等方面都具有重要作用[28]。本研究中,放线菌门在秋季的相对丰度均显著高于春季,这与春季消落带退水初期土壤含水量高有一定关系,土壤水分过多会降低其透气性,而土壤含氧量较高的环境更适合放线菌门的生长,同时,放线菌能够分解纤维素和木质素[29],秋季的土壤中,由于经过了一个生长季,可能有更多的纤维素和木质素,提供了更多的营养源供给放线菌的生长繁衍。Lauber等[30]发现芽单胞菌门更适合在土壤含水量较低的环境下生存,芽单胞菌门在夏季的相对丰度均显著低于秋季,这与取样年份研究区降水过多有关,且取样当年夏季水位线一直保持较高水平,这也在一定程度上说明了环境条件的变化会影响细菌群落的组成和分布。
3.2 不同季节适生草本根际土壤细菌群落与多样性的影响因素
利用多样性指数分析土壤细菌群落是一种非常有效的方法[31],土壤细菌多样性对提高土地生产力、增强胁迫环境下群落稳定性有着重要影响[32]。本研究发现,根际细菌α多样性的变化与季节显著相关,说明土壤环境的季节变化显著影响适生草本根际细菌群落的丰富度和均匀度,且根际细菌群落的α多样性指数都表现为秋季>夏季>春季,即根际细菌种类随时间推移逐渐增加。有学者通过研究呼伦贝尔草原土壤细菌多样性的季节变化特征,发现α多样性指数在夏季最高,其次为秋季[33]。但本研究结果与之不同,这与三峡库区反季节水位调度方式有关,即夏季低水位运行,冬季高水位运行,春季消落带水位逐渐下降,植物初露开始生长,细菌群落多样性最低;夏季温度最高,植被逐渐恢复生长,群落多样性较春季有所上升,但取样当年研究区夏季降水较多,土壤通气性和水分入渗减少,厌氧条件会限制好氧微生物的生长[34];秋季降水较夏季减少,植物生长繁茂,植被的生长可以为细菌提供分泌物等一系列资源,故秋季群落多样性最大。总体而言,这些结果表明,适生草本根际细菌群落多样性对季节性环境波动有明显响应。
土壤温度、湿度、pH值、有机和无机养分水平的变化可以影响微生物群落的结构[35]。RDA分析显示,影响研究区适生草本根际土壤细菌群落结构的理化因子主要有土壤ORP、土壤有机质、土壤全磷和全钾等。Husson证明了土壤ORP是土壤、植物、微生物系统的主要驱动因素[36],这与本研究结果相似,消落带为水陆交错地带,干湿交替的土壤环境下,土壤氧化还原电位降低,促进还原物质产生,有利于有机质及氮元素的积累[37],从而对细菌群落结构造成影响;土壤有机质对秋季细菌群落结构有显著影响(P<0.05),有机质含量与土壤肥力存在着密切的关系,可为植物提供营养物质,促进植物的生长发育,同时也可改变土壤的物理性质,促进土壤微生物的活动[38],本研究中土壤有机质含量与细菌群落结构显著相关,其原因是随着人工修复草本植被年份的增加,土壤肥力得到很大改善,土壤根系分泌物增加,进而导致微生物群落结构的改善。
3.3 不同季节适生草本根际土壤细菌功能特征
植被对土壤细菌群落结构的影响会进一步影响土壤细菌功能组成[39],目前三峡库区消落带土壤微生物研究主要集中在细菌群落多样性[40],对细菌功能方面鲜有研究。PICRUSTt较宏基因组研究成本低,更加方便快捷,已广泛的用于土壤[41]、水体[42]等不同生境的微生物功能研究。
PICRUSt2功能预测结果表明,三峡库区消落带适生草本根际土壤中共有代谢、遗传信息处理、环境信息处理、细胞过程、有机系统、人类疾病6个一级功能层和膜运输、翻译、氨基酸代谢、碳水化合物代谢等45个二级功能层。土壤代谢功能在一级功能层中占比最高,这与对湿地土壤细菌功能研究的结果一致[43]。部分细菌类群可通过代谢产生抗生素、抑菌蛋白及促生长激素等,从而提高消落带适生草本植物抑制病原菌的能力,促进植物的生长[44]。
碳和氮的数量和质量可以通过调节细菌群落组成来驱动地下生态功能[45]。二级功能层中,膜运输、翻译、氨基酸代谢、碳水化合物代谢的功能基因丰度最高,总计超过了36%。土壤细菌活跃的膜运输功能有利于维持细菌各项生命活动,是土壤细菌正常发挥其功能的基础[46]。氨基酸代谢主要通过脱氨作用、转氨作用、联合脱氨或脱羧作用分解成α-酮酸、胺类及CO2,是碳代谢和氮代谢的中枢;碳水化合物合成与代谢则与氮、磷循环呈正相关,间接说明土壤细菌丰富度越高越有利于氮、磷的转化和迁移[47],适生草本根际细菌中高丰度的此类功能基因,证明了根际土壤细菌在氮磷循环中发挥着重要的作用,同时氨基酸代谢可助细菌吸收氨基酸[48],利于适生草本在消落带特殊生境下的存活与繁衍。根际土壤细菌α多样性指数在春季最低,而膜运输基因则在春季表现出最高丰度,这一现象与生态系统的自我调节机制有关,高丰度的膜运输功能基因可调节溶质如离子和小分子等通过生物膜,以促进根际土壤细菌在消落带退水初期快速生长。同时,各代谢途径在不同季节中也表现出一定差异,这与不同季节的植物相关活动有关,植被凋落物和根系分泌物的季节差异会改变土壤细菌组成和多样性,进而对土壤功能微生物产生特定的影响[49]。
尽管PICRUSt2功能预测能够对细菌功能进行分析,但由于该功能预测没有基因数据支持,存在一定缺陷[50],后续可结合宏基因组技术,对根际细菌与适生草本的相互作用机制进行进一步的研究。
4 结论
三峡库区适生草本植物根际土壤细菌多样性在秋季显著大于夏秋两季,而细菌群落组成在各季节差异不大,其中变形菌门、酸杆菌门、拟杆菌门、绿弯菌门为适生草本根际土壤中的优势菌门。土壤ORP和TP对春季细菌群落影响较大,AK对夏季细菌群落影响显著,OM和SWC对秋季细菌群落影响显著。根际土壤细菌功能预测表明,三峡库区消落带适生草本根际土壤中与C、N、P有关的细菌代谢功能丰度较高。
猜你喜欢
中华建设(2023年10期)2023-10-07
少儿美术(2020年8期)2020-11-06
四川地质学报(2020年2期)2020-05-31
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
现代园艺(2017年13期)2018-01-19
山西农经(2016年3期)2016-02-28
小说月刊(2014年1期)2014-04-23
