
中国大中型陆栖哺乳动物地理区边界探测及其驱动因素
2023-12-26 02:43吴朝宁王结臣
生态学报 2023年23期
迟 瑶,刘 璐,吴朝宁,王结臣,4,*
1 浙江工商大学旅游与城乡规划学院, 杭州 310018
2 南京大学地理与海洋科学学院,自然资源部国土卫星遥感应用重点实验室,江苏省地理信息技术重点实验室, 南京 210023
3 中船智海创新研究院有限公司, 北京 100094
4 江苏省地理信息资源开发与利用协同创新中心,南京 210023
地理隔离是指种群间受地理屏障阻碍的影响,而无法顺利进行基因交流的现象[1]。由于受到地理隔离的作用,物种的分布会被限制在局部区域内[2],进而阻碍了生物种群间的基因流动,威胁到种群的生存繁殖[3—4]。物种空间分布格局的形成受到物种相互作用和非生物因素的影响[5]。地理隔离是影响生物在地理学上空间分布特征的主要因素之一,主要表现为距离隔离[6]和环境隔离[7]。生物在进入隔离的新环境后发生选择进化,而后地理隔绝的环境又保持了生物的多样性,因此,地理隔离在生物多样性的演化中扮演了很重要的角色[8]。当前自然环境和人类活动的共同作用使得人地交互的生态系统模式愈渐复杂,导致地理隔离界线的识别分析较为困难。生物地理区边界可能是表征对物种具有隔离效应的界线,通过识别以不同物种组合为特征的生物地理区域,可以揭示物种在空间上的分组方式,为探讨各动物类群的隔离机制提供空间模式和框架。
物种共存并相互作用以形成具有相似分类或生态生理特征的集群[9]。这种集群分布的形成反映了物种对当前阶段环境历史变化的适应[10—11]。构造运动、地形和气候环境等多种因素共同塑造了陆地生物多样性的演变,形成了不同的生物地理区域[12—13]。在19世纪,学者们试图将整个世界的陆地表面划分为动物地理领域[14—15]。随着物种分布数据和新的多元方法的日益增加,利用定量方法对生物地理区域的研究迅速增加。近几十年来,大多数研究根据物种发生数据对生物区域进行分类,并通过聚类为具有地理意义的组来显示区域之间的关系[16—17]。相比传统基于相似性指数聚类的方法,基于物种和格网单元的二分网络聚类方法,在大陆尺度下根据物种分布数据划分生物地理区域更为有效[18]。
定量化的生物地理区域研究也为探讨生物地理区域与环境之间的关系开辟了新的途径。基于丰富的环境因子数据和数理统计学等方法,国内外学者进一步开展了生物地理分布与各种环境因素之间关系的定量研究。相关分析可以用于检验不同生物分类群地理分布模式的一致性[19],广义线性模型适用于分析物种和环境变量之间的非线性关系[20],空间回归模型等常被用来探究环境因子对物种空间分布的影响程度[21]。随着计算机技术的发展,机器学习等复杂模型在模拟物种分布方面具有很强的预测能力,经常被用来探索和预测哪些环境因素影响了物种的空间分布[22—23]。景观、气候、地形以及人类社会进程都会影响动物的分布[24—25]。已有研究表明,生物地理区的范围界线由广泛的地球物理过程、历史演化和生物因素共同决定[26]。随着人口激增,人类对生态系统施加的压力也在逐渐增强。人类干扰引起的负面影响可能导致全球生物多样性锐减,进一步限制物种分布范围。城市化等土地利用转化通过改变地表结构扰乱生态群落,导致栖息地破碎和退化,从而威胁脊椎动物的活动可达范围[27]。
尽管生物地理区受多种地理环境因子的影响是普遍共识,但关于其边界的定量研究却鲜见,且各因子在不同区域的影响程度尚不明确。探索生物地理区边界的驱动因素及空间分异特征对分析生物多样性空间演变和规划重要保护区域等具有重要意义。且多类群物种对隔离屏障的响应模式难以统一,大中型哺乳动物在复杂地形内和极端环境条件下的食物能量供给更难支持其进行持续跨越障碍或长期栖息,地理环境屏障对大中型动物扩散的阻隔效应更为明显,从大中型动物入手研究生物地理区的划分有助于揭示地理隔离效应。
基于此,本研究针对大中型陆栖哺乳动物类群,利用四叉树数据结构的自适应分辨率算法构建的二分网,引入Infomap聚类方法探测的生物地理区域。从温度、降水和地形的梯度变化角度,基于空间自回归模型和地理加权逻辑回归(GWLR)模型分析影响生物地理区域边界的驱动因素,进而探究气候与地形变化对大中型陆栖哺乳动物的隔离效应。以此为解释其它不同类群动物的空间分布格局与环境变化之间的关系提供一定的借鉴意义。
1 研究数据
1.1 物种分布数据
动物分布数据主要来自《中国哺乳动物多样性及地理分布》[28]中所记录的物种实际分布范围。《中国哺乳动物多样性及地理分布》中物种分布图是参照国内外的资料,并依据蒋志刚团队野外考察结果绘制的。本研究利用扫描仪对书籍进行数字化扫描,裁剪出物种分布范围图片,利用ArcGIS软件对图片进行校正配准、对物种分布范围进行矢量化处理,以获取各物种分布范围。并对各物种名录和基本信息数据进行补充修订,对其分布范围进行核对,以此得到各物种分布图。本次哺乳动物红色名录共评估了12目55科245属673种哺乳动物,其中陆栖哺乳动物包括11目629种,排除人类物种和在中国境内已灭绝的3种犀牛,包括大独角犀、爪哇犀和双角犀,以及濒临绝种的钓鱼台鼹后,共计11目42科209属624种,物种所属目涉及鲸偶蹄目、劳亚食虫目、鳞甲目、灵长目、啮齿目、攀鼩目、奇蹄目、食肉目、兔形目、翼手目以及长鼻目。目前,对于大、中、小型哺乳动物的体重划分没有明确的界定标准,一般来说,体重超过15kg的为大型食肉动物[29]。本文所分析的陆栖哺乳动物(除灵长目)体重均在15kg以上,故统称为大中型陆栖哺乳动物,共5目11科31属73种(表1),具体统计情况和物种的体重体型及濒危等级等详细属性见表2。通过投影变换使各物种分布图具有统一的Albers等面积圆锥投影。

表1 大中型陆栖哺乳动物分类统计表

表2 大中型哺乳动物的属性信息统计表
1.2 驱动因子数据
本研究收集了重要地形地貌数据和相关环境因子数据。其中,青藏高原在中国境内的边界数据来源于国家青藏高原科学数据中心张镱锂等人的《青藏高原边界数据总集》[30],主要盆地数据来源于美国地质调查局(USGS),包括准噶尔盆地、吐哈盆地、塔里木盆地、鄂尔多斯盆地、柴达木盆地、四川盆地等共15个盆地。选取的主要岛屿包括台湾岛和海南岛,来源于中国行政区划数据。我国重要地理分界线包括三级地势阶梯、三大自然区界线、“胡焕庸线”、秦岭-淮河线等数据来源于科技基础性工作专项“新世纪版《中华人民共和国国家大地图集》编研(2013FY112800)”,该数据集的空间基准采用北京1954大地坐标系,地图投影为双标准纬线等积圆锥投影,双标准纬线为25°N和47°N,中央经线为105°E。高程数据的空间分辨率为30m,来源于先进星载热发射和反射辐射仪全球数字高程模型(ASTER GDEM)。温度、降水数据的空间分辨率为30弧秒,来源于WorldClim数据集(http://www.worldclim.org/),WorldClim是一组可用于制图和空间建模的覆盖全球的网格化气候数据。人类足迹指数(HFI)是综合考虑建筑环境的范围、人口密度、电力基础设施、农田、牧场、道路、铁路和可通航水路等有关变量,来反映人类对自然系统施加压力强度的位置信息数据。人类足迹指数数据来源于美国宇航局地球观测系统数据和信息系统发布的2005年第2版“全球人类足迹数据集” (https://sedac.ciesin.columbia.edu/data/set/wildareas-v2-human-footprint-geographic)。所有地理环境因子数据均转换到Albers等面积圆锥投影。
2 研究方法
2.1 信息地图生物区方法
本研究基于信息地图生物区(Infomap Bioregions)算法识别我国大中型陆栖哺乳动物的生物地理区域,进而提取可能对大中型陆栖哺乳动物类群的分布扩散产生阻隔效应的主要界线。Infomap Bioregions方法可以基于物种分布数据识别特定分类单元的生物区域。已有研究表明基于物种和格网单元的二分网络聚类方法受到抽样偏差的影响较小,识别结果会更加独立于分析尺度,优于单独基于格网单元间物种相似性的聚类方法[9,18]。Infomap Bioregions方法的关键步骤为:(1)利用四叉树映射生成分级数据。该方法基础输入数据为物种分布数据,需要包括物种名称及对应的空间位置信息。将物种记录添加到与相应物种范围多边形相交的最小尺寸的单个格网单元中,然后基于自适应分辨率算法,根据输入数据的数量和空间分布自动调整网格大小。自适应分辨率算法通过将输入数据映射到四叉树数据结构来实现。每个格网单元会被递归地细分为四个象限,进而将地理空间分层划分为越来越小的次级格网单元。为了使分辨率适应数据的密度,需要为每个格网单元指定最大容量(例如100个物种出现记录)和最小容量(例如10个物种出现记录)。递归细分所有记录多于最大单元容量的格网单元,直到达到指定的最小单元容量。如果细分后的格网单元所包含的物种少于指定的最小单元容量,则算法会恢复前次划分以避免创建数据点太少的区域。基于以上标准,Infomap Bioregions可以同时识别数据丰富的高分辨率生物区域和数据稀疏的低分辨率生物区域,从而避免所有生物区域的过度拟合和欠拟合。(2)利用网络映射生成二分网络。Infomap Bioregions将物种聚合成地理格网单元时,形成了一个以物种和格网单元为两类节点的二分网络。每个物种都通过未加权链接连接到它所在的每个格网单元,而非通过记录数量对链接进行加权,从而避免聚类结果对空间偏差采样的敏感。(3)基于网络聚类识别生物区域。Infomap Bioregions基于Infomap算法进行聚类,生成的集群包含格网单元和物种,同时定义了不同的生物区域。Infomap算法基于随机游走算法的基础上,将社团检测转变为编码问题,通过最小描述长度原则来寻找最优社区,以展示大型网络内部的集聚特征[31]。Infomap Bioregions中使用Infomap算法划分生物地理区域,可使区域内各节点联系较为紧密,物种组成更为相似,区域间各节点联系较为稀疏,物种组成差异明显,因此各生物地理区域的分界线则可能对应于对物种分布有明显阻隔的界线。
2.2 驱动因子的选取
以往研究表明,气候突变、地形起伏明显以及人类活动强度大的区域会对大中型陆栖哺乳动物的分布扩散产生阻隔效应。选取WorldClim数据集中反映气候的19个气温、降水因子,通过高程数据映射地形障碍,采用人类足迹指数来映射人类活动强度对生态系统的影响程度。由于气温降水变量间存在相关性,会严重影响模型的拟合效果。采用皮尔逊相关系数量化气候变量间两两相关性程度,见表3。基于K-means聚类方法将相关性程度高的变量划分至同一组中,共聚类为四组,其中年平均气温Bio1、最热月份的最高温度Bio5、最冷月份的最低温度Bio6、最湿季度的平均温度Bio8、最干燥季度的平均温度Bio9、最热季度的平均温度Bio10、最冷季度的平均温度Bio11为一组,平均昼夜范围Bio2、年降水量Bio12、最湿月份的降水Bio13、最干燥月份的降水Bio14、最湿季降水Bio16、最干燥季度的降水Bio17、最热季降水Bio18、最冷季降水Bio19为一组,等温性Bio3、降水量季节性变化Bio15为一组,气温季节性变动系数Bio4、年温度范围Bio7为一组。每组保留一个因子,结合相关的文献研究,选取年平均气温Bio1、气温季节性变动系数Bio4、年降水量Bio12和降水量季节性变化Bio15四个与哺乳动物分布密切相关的指标来代表气候特征,其中,气温季节性变动系数是月平均气温的标准差,降水量季节性变化是月降水量的标准差与月平均降水量之比。这四个变量的线性变换组合与最热月份最高温度、最冷月份最低温度、最湿月份降水量、最干月份降水量等其它温度、降水变量联系紧密,具有代表性。筛选后的四个变量间,年平均气温与年降水量的相关性最高,其对应皮尔逊相关系数为0.642,小于0.7,说明四个变量间不存在强相关关系,符合后续建模条件。

表3 19个温度、降水因子相关性
由于大中型哺乳动物的分布边界往往与气候、地形的突变点相关,本文基于基础的地理环境相关数据,进行一定的变换,以更好的反映环境突变特性。针对气候数据,中心像元的气候障碍计算为中心像元与周围八邻域像元之间的变异系数。变异系数是一组数据相对于总体平均值的变异程度,可以消除量纲的影响,此处用于检测与周围环境的气候特征有明显差异的区域单元。针对高程数据,通过计算中心像元与周围八邻域像元之间的平均差(即差绝对值和的平均值)表示海拔高度差异,进而反映中心像元的地形障碍强度[13]。最终选择驱动生物地理区域边界存在的变量指标(表4),其中,气候因素包括年平均气温变异系数、气温季节性变动系数的变异系数、年降水量变异系数和降水量季节性变化变异系数,相比于初始数据,经过转换后的气候因素解释变量更能反映我国大陆气候突变趋势特征;地形因素解释变量的地形障碍是基于高程数据转换而来,相比于直接的高程值更能反映地形突变界线;人类活动强度因素选择综合考虑城市建设、人口密度和农田牧场以及交通设施等有关变量而来的人类足迹指数指标作为解释变量,来反映研究区域内人类活动强度的空间差异性,值越高的地区表明人类活动对生态系统的影响越强,则越有可能影响大中型陆栖哺乳动物的空间扩散。

表4 驱动生物地理区域边界存在的变量指标
本研究采用全局莫兰指数(Global Moran′sI)[32]量化评价各解释变量的空间自相关程度,基于空间自回归模型量化选择主要驱动因子。空间自回归模型的两种经典模型是考虑自变量空间自相关性的空间误差模型和考虑因变量空间自相关性的空间滞后模型,研究表明,空间误差模型更适合于有关物种分布数据的回归分析,因为物种扩散会受到相邻区域气温、降水、地形以及人类活动强度等环境因素的影响[33]。空间误差模型的表达式如下所示:
Y=Xβ+λWu+ε
(1)
式中,Y是代表因变量的n×1维向量,本研究表示是否为生物地理区域边界;X是代表自变量的n×(k+1)矩阵,k是自变量因子数,本研究表示筛选出的相关环境因子;β是代表模型待估回归系数(含截距)的(k+1)×1维向量;W是n×n维的空间权重矩阵,u表示误差项,Wu是反映生物地理区域边界形成不仅与当前区域环境因素有关,也与相邻区域的环境因素有关的空间误差效应;λ是代表空间自相关误差项的待估系数;ε是代表模型残差的n×1维向量,是独立同分布。利用赤池信息量准则(AIC)模型评价拟合度。计算衡量模型拟合程度的决定系数R2,一般认为,R2>0.4时,拟合效果较好。采用Z检验进行回归系数的显著性检验。
2.3 地理加权逻辑回归
采用地理加权逻辑回归方法评估各因子在不同区域对生物地理区域边界形成的影响程度差异。地理加权逻辑回归模型(GWLR)是基于传统逻辑回归和地理加权回归的拓展模型,考虑了空间非平稳性现象,在局部区域对每个坐标点运用加权最小二乘法回归进行参数估算,每个坐标点对应一组回归系数。多用于解决因变量为二分类变量的情况[34]。传统的逻辑回归模型未考虑模型变量的空间异质性,假设其为平稳状态,拟合结果无法全面反映变量的空间关系[35]。在对具有空间结构的数据进行拟合分析时,应考虑空间地理位置变化对因变量结果的影响,基于地理加权逻辑回归模型的拟合效果往往优于传统逻辑回归模型[36]。
与传统逻辑回归模型类似,GWLR的表达式中,位置i在边界处(Y=1)的概率为P,未在边界处(Y=0)的概率为(1-P),则位置i在边界处的概率表示为:
(2)
式中,z为位置i对应的局部回归模型的线性函数:
z=α0(ui,vi)+α1(ui,vi)xi1+α2(ui,vi)xi2+…+αn(ui,vi)xin
(3)
对概率P进行对数变换,则有:
(4)
之后,基于加权最小二乘法估算回归系数:
(5)
(6)

(7)
式中,dij表示回归位置点i和观测点j的距离,b表示带宽,即选择在多大的距离阈值内或多少近邻数来构建GWLR模型,采用修正的赤池信息量准则(AICc)方法来选择最佳带宽,AICc值越小对应的带宽越优。
2.4 模型拟合效果检验
受试者工作特征曲线(ROC),适用于解释变量为连续变量、因变量为二分类变量的情况,可应用于诊断二分类判别效果的准确性,克服了其他传统准确性评价指标的局限性[37]。 ROC曲线下的面积(AUC)是准确度的度量,值越接近1,模型的准确度越高。本研究采用ROC曲线进行地理加权逻辑回归模型的拟合效果检验,通过计算AUC的值进行量化评价,当AUC>0.85,可表明自变量对二分类因变量的解释能力较高[38],反映出模型具有良好的区分能力。
3 结果与分析
3.1 基于Infomap Bioregions的生物地理区域探测
本文基于Infomap Bioregions算法识别我国大中型陆栖哺乳动物的生物地理区域,进而提取可能对其分布产生阻隔效应的主要界线。排除仅分布在台湾的台湾梅花鹿(Cervustaiouanus)、台湾鬣羚(Capricornisswinhoei)与仅分布在海南的海南麂(Muntiacusnigripes)、东方坡鹿(Panoliasiamensis)的特有种,将剩余69种物种的分布范围面数据作为输入数据。为反映物种分布数据密度的空间差异,将格网单元范围限制在0.125°和0.25°之间,以通过较高分辨率限制,来更好地识别数据丰富的区域。指定每个格网单元的最大容量为5个物种出现记录和最小容量为1个物种出现记录,构建以物种和格网单元为两类节点的二分网络,试验次数和集群成本采用默认的推荐值1,基于Infomap算法进行聚类,合并具有相似物种分布的格网单元,进而识别不同的生物区域。剔除结果中面积很小且分散的不具有生物地理学意义的区域后,我国大中型陆栖哺乳动物的生物地理区域识别结果如图1所示。
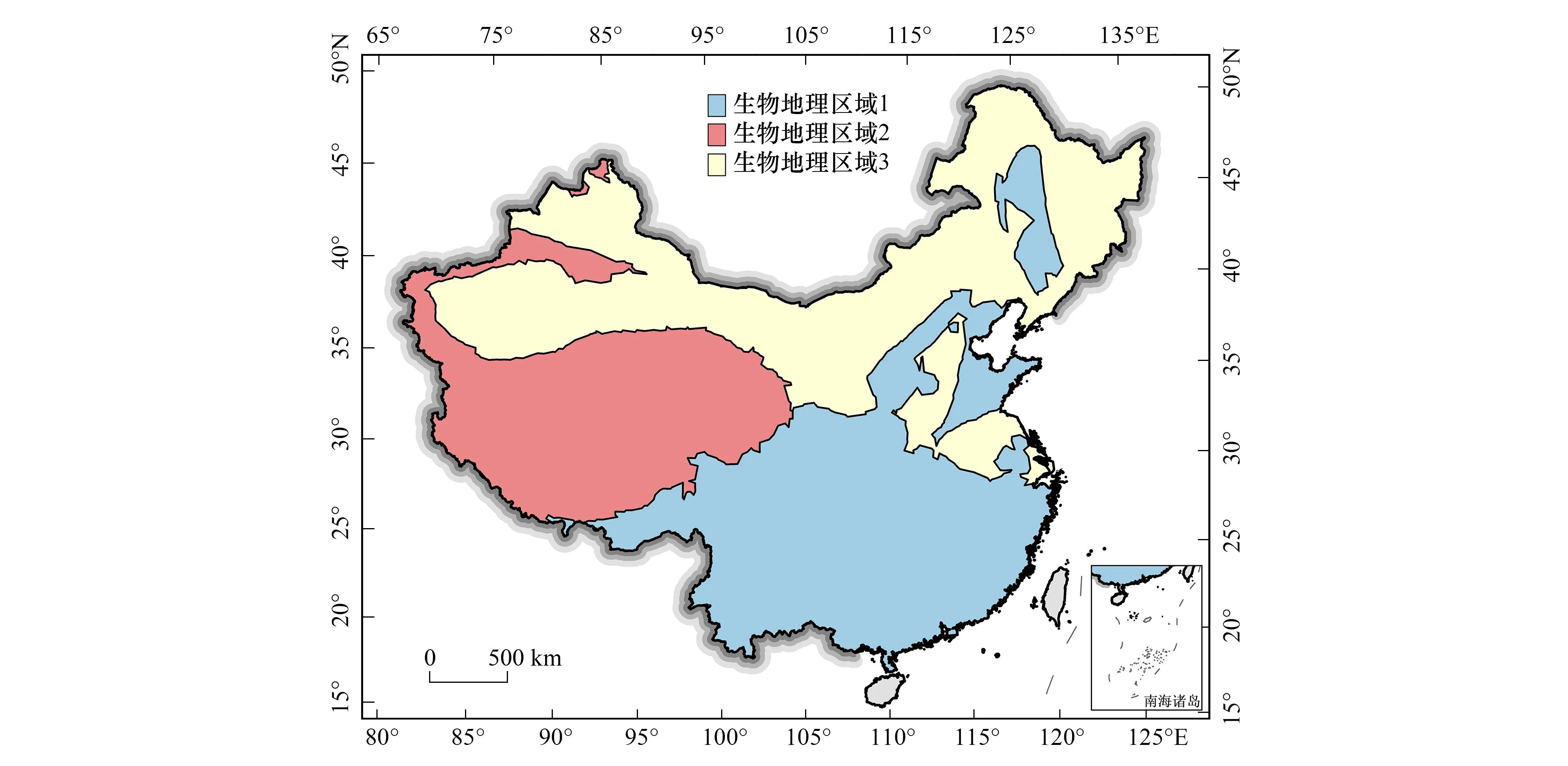
图1 中国大中型陆栖哺乳动物生物地理区域识别结果
基于我国大中型陆栖哺乳动物分布数据识别出的三大生物地理区域,生物地理区域1以秦岭-淮河以南的地区为主,向西延伸至青藏高原东南方向气候湿润的横断山脉地带;生物地理区域2主要位于青藏高原除东南区域以外的地区,并向北沿塔里木盆地边缘延伸至准噶尔盆地、吐哈盆地与塔里木盆地的中间地带;生物地理区域3主要分布区与西北干旱区类似,但在秦岭-淮河线以北的东部季风区,生物地理区域1与生物地理区域3呈交错分布格局,生物地理区域1主要分布在温带季风性气候的东北平原、吕梁山脉与太行山脉中间地带以及华北平原的东部沿海区。生物地理区域2和生物地理区域3的分界线与青藏高寒区和西北干旱区的分界线较为吻合,即与青藏高原北部边界线相符;生物地理区域1和生物地理区域2的分界线与青藏高寒区和东部季风区的分界线明显不同,但与我国季风区与非季风区界线的分布趋势较为相似。
3.2 生物地理区域边界的主要驱动因素
本研究基于50km的分辨率计算各解释变量对应数据(图2)。各气候要素对应的变异系数可反映对应要素与周边区域的差异程度,变异系数值越大,差异越明显,局部异质性越高。我国年平均气温整体呈“南高北低”趋势。年平均气温变异系数值变化趋势与我国三级阶梯分界线较为吻合,年平均气温的局部异质性从一级阶梯至三级阶梯由西向东逐渐降低,青藏高原、塔里木盆地、四川盆地、鄂尔多斯盆地以及东北平原和华北平原等重要地形边缘的变异系数明显高于周边区域。生物地理区域2的边界线附近的变异系数值明显较高,东部区域的生物地理区分界线年平均气温局部异质性低于西部区域的分界线。气温季节性变动系数整体上呈由南向北逐渐递增的梯度分布,说明我国全年气温波动程度由南向北逐渐变高。变异系数高值区主要在青藏高原边缘以及塔里木盆地边缘,生物地理区域2的边界线变异系数值较高,且明显高于东部生物地理区边界。我国年降水量整体呈由东南向西北递减的趋势,青藏高原西北部与西北干旱区的沙漠地带年降水量明显较少。整体上生物地理区域边界所在区域的年降水量基本处于较低水平。非季风区的年降水量局部异质性明显高于季风区,对应西北区生物地理分界线局部异质性高于东南区分界线。降水量季节性变化在东部季风区秦岭-淮河线以南区域明显较低,青藏高原中西部区域、华北平原和东北平原的降水量季节性变化尤为明显。变异系数高值区主要在我国西北地区,生物地理区域2与生物地理区域3的分界线变异系数值较高,且明显高于其他区域生物地理区边界。地形障碍可反映地形起伏程度大小,我国三级阶梯地势分界线处的地形障碍值明显较高。人类足迹指数可反映人类活动强度,以胡焕庸线为分界线的东西两侧人类活动强度差异显著,东部生物地理区边界所在区域的人类足迹指数值明显较大。

图2 研究区初始环境因子和对应解释变量数据的分布
计算各解释变量的莫兰指数(Moran′sI)和方差膨胀因子(VIF)(表5),Moran′s I结果均大于0.5且Z得分均大于2.58,表明通过了0.01水平下的显著性检验;VIF结果均小于4,表明变量之间不存在明显的多重共线性,可以参与模型建模。由于本研究物种分布数据是我国范围内的数据,解释变量年平均气温CV、气温季节性变动系数CV、年降水量CV、降水量季节性变化CV、地形障碍在我国大陆边界区域表现出了明显的高值特征,为避免边缘数据对实验结果的误差影响,排除我国大陆边界线50km缓冲带内的数据参与建模。

表5 各解释变量的空间自相关性检验及多重共线性检验结果
空间自回归分析结果显示,基于6个解释变量构建的空间自回归模型的AIC相比仅考虑空间随机效应(不考虑解释变量)构建的空间自回归模型的AIC下降了147.98,且决定系数R2为0.53,大于0.4,表明针对大中型陆栖哺乳动物的生物地理区域边界的空间自回归模型拟合结果较好。
基于空间自回归模型探究气候、地形及人类活动与大中型陆栖哺乳动物生物地理区域边界的全局关系,估计结果如表6所示,年平均气温CV的回归系数为0.179,在5%的水平下显著;气温季节性变动系数CV的回归系数为0.694,在0.1%的水平下显著;年降水量CV的回归系数很小,为-0.076,且P值为0.584,不显著,表明年降水量的局部异质性与生物地理区边界的联系并不明显;降水量季节性变化CV的回归系数为0.338,在1%的水平下显著;地形障碍的回归系数为0.228,在10%的水平下显著;人类足迹指数的回归系数为0.645,在0.1%的水平下显著。综合对比各解释变量的回归系数结果,在10%及以下的显著性水平,年平均气温CV、气温季节性变动系数CV、降水量季节性变化CV、地形障碍和人类足迹指数与生物地理区边界均呈正相关,其中,气温季节性的异质性与人类足迹指数是影响程度最大的变量,其次是降水量季节性的异质性和地形障碍,相比之下,年平均气温的局部异质性对生物地理区边界的作用较弱。回归结果表明,生物地理区边界是气候突变、地形障碍和人类活动强度协同作用下表现出的结果。

表6 空间自回归模型估计结果
3.3 基于GWLR模型的生物区域边界驱动因素分析
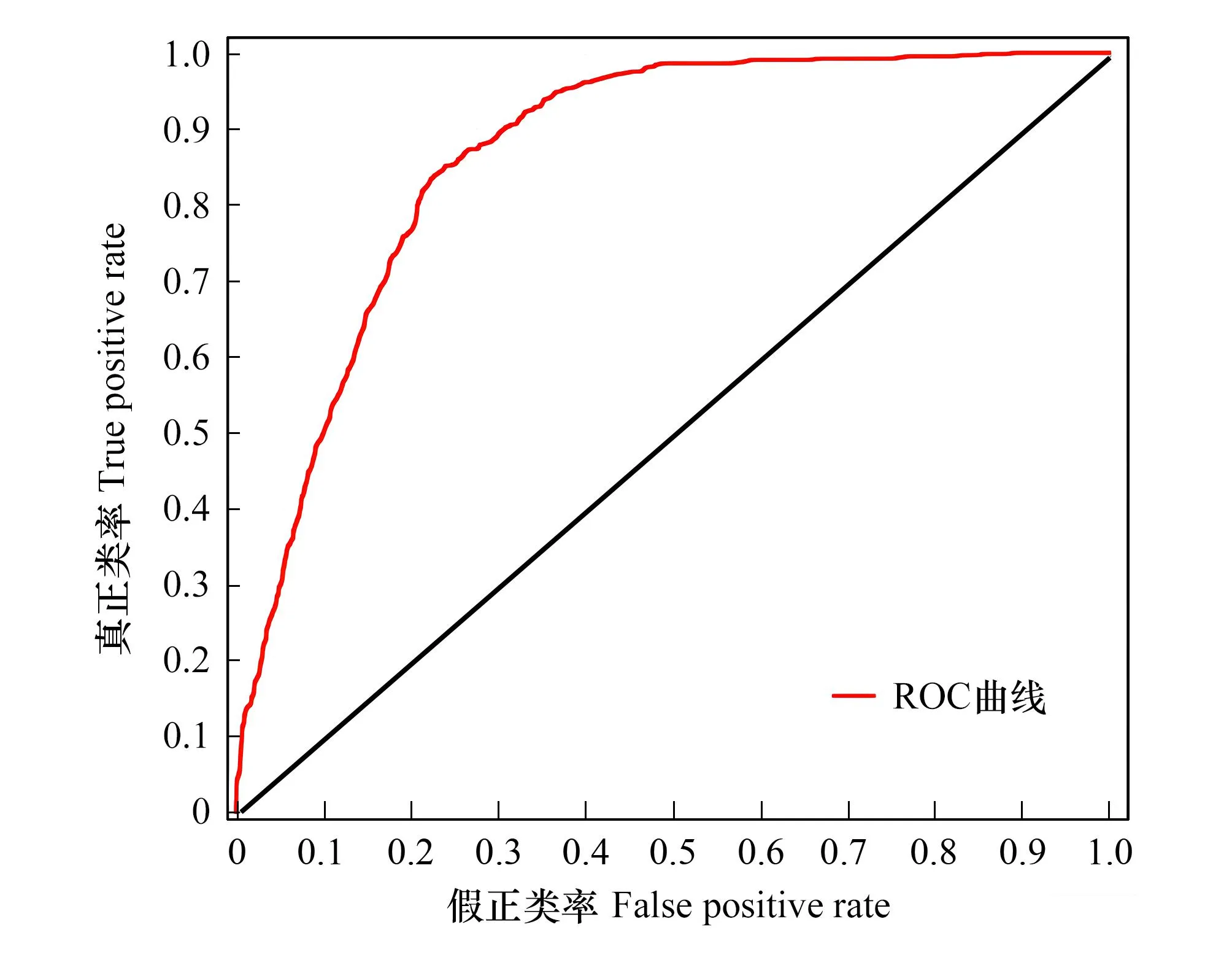
图3 地理加权逻辑回归模型拟合效果检验受试者工作特征曲线(ROC)
研究通过量化气候突变、地形障碍以及人类活动强度因素,基于GWLR模型解释不同类群生物区域边界可能的隔离成因,评估不同地理因素阻碍大中型陆栖哺乳动物生物区边界扩张的程度差异。基于空间自回归模型的估计结果,选取通过10%及以下显著性检验的解释变量即年平均气温CV、气温季节性变动系数CV、降水量季节性变化CV、地形障碍和人类足迹指数参与地理加权逻辑回归建模。GWLR模型的空间权重函数采用固定高斯核函数,运用区间搜索法在250km到1000km的区间内基于AICc准则选择带宽,最终确定的带宽为650km。GWLR模型对应的AICc值为2268.040,GWLR模型的异常解释百分比为29%,GWLR模型的AUC值(ROC曲线下面积)为0.866(图3),反映出GWLR模型拟合效果较好,各驱动因子对我国大中型陆栖哺乳动物的生物地理区边界的解释能力较好。
GWLR模型结果表示,从回归系数的均值来看,各解释变量均与生物地理区边界呈正相关。为方便直观分析GWLR模型中大中型陆栖哺乳动物生物地理区边界驱动因子回归系数的空间非平稳性特征,绘制GWLR模型中各解释变量估计系数的空间分布图和对应的检验值t值空间分布图。表7和图4表明,各解释变量的估计系数和检验值t值均在研究区域内呈明显的空间异质性特征,各解释变量的估计系数和检验值t值的空间分布趋势相一致。当t>1.65时,表示在10%的水平下正显著。

图4 各解释变量估计系数和检验值t的空间分布

表7 GWLR模型参数估计结果
本研究关注的是与生物地理区边界相关的因素,着重分析各驱动因子的正显著作用区域。年平均气温CV的正显著作用区域集中在我国季风区,且由西向东逐渐加强,反映出在季风区,年平均气温的局部异质性对生物地理区边界的促进作用更为明显;气温季节性变动系数CV的正显著作用区域集中在我国中部和西部地区,且呈由东北向西南方向递增的趋势,生物地理区域2边界所在区域的估计系数值明显较高;降水量季节性变化CV的正显著作用区域集中在生物地理区3边界所在区域,其估计系数以内蒙古高原地区为中心,呈内高外低的辐射状分布;地形障碍的正显著作用区域集中在我国西部地区,主要在青藏高原中西部和塔里木盆地一带,且由东向西逐渐加强;除我国西北部的塔里木盆地区域外,其他区域人类足迹指数均呈正显著分布,其估计系数从我国东南方向起呈内高外低的辐射状分布,华北平原区域的估计系数值明显较高,其次是东北平原地带,生物地理区2边界所在区域的估计系数值则较低。
统计各地区所有正显著解释变量中最大估计系数值对应的解释变量(图5),分析生物地理区边界的主导驱动因子的空间异质性特征,结果表明,我国东部区域即生物地理区1与生物地理区3的边界线的主导影响因子是人类足迹指数,生物地理区1和生物地理区2的边界线的主导影响因子是气温季节性变动系数CV,生物地理区2和生物地理区3边界线在祁连山脉一带的主导影响因子主要是气温季节性变动系数CV,其余地区为地形障碍。统计不同主导因子的作用区域分布比例(表8),人类足迹指数占比50.62%,气温季节性变动系数CV占比26.44%,地形障碍占比20.71%,结果反映出人类活动对大中型陆栖哺乳动物分布表现出了显著影响。
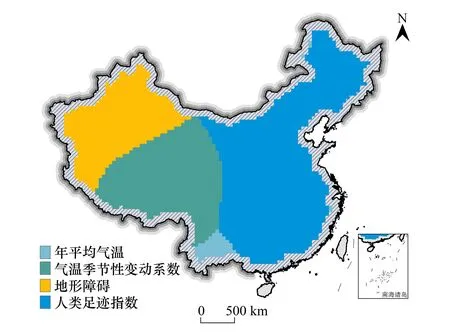
图5 生物地理区主导驱动因子

表8 不同主导驱动因子作用区域分布比例
3.4 生物地理区域边界与物种分布分析
统计各生物地理区域的物种组成成分,结果见表9。生物地理区域1共计49种物种,其中分布面积占比在80%以上的物种有29种,分布面积占比在20%以下的物种有8种,剩余24%的物种,主要为分布范围较为广泛的黑熊(Ursusthibetanus)、豺(Cuonalpinus)、野猪(Susscrofa)、狼(Canislupus)、狍(Capreoluspygargus),和分布在该生物地理区域边界线附近的喜马拉雅斑羚(Naemorhedusgoral)、西藏马鹿(Cervuswallichii)、四川梅花鹿(Cervussichuanicus)、四川马鹿(Cervusmacneilli)等。生物地理区域2共计40种物种,其中分布面积占比在80%以上的物种有7种,分布面积占比在20%以下的物种有20种,剩余33%的物种,主要为分布范围较为广泛的猞猁(Lynxlynx)、棕熊(Ursusarctos)、狼,和分布延伸至该生物地理区域东南边缘外横断山脉地带的雪豹(Pantherauncia)、岩羊(Pseudoisnayaur)、白唇鹿(Przewalskiumalbirostris)等,以及沿该生物区边缘分布的北山羊(Caprasibirica)、四川马鹿、天山盘羊(Oviskarelini)、四川梅花鹿、塔里木鹅喉羚(Gazellayarkandensis)、西藏马鹿和喜马拉雅斑羚等。生物地理区域3共计43种物种,其中分布面积占比在90%以上的物种有15种,分布面积占比在20%以下的物种有18种,剩余23%的物种,主要为分布较为广泛的狍、狼、野猪、棕熊、猞猁、豺等和沿该生物区边缘分布的塔里木鹅喉羚、天山盘羊、北山羊等。

表9 各生物地理区域物种面积占比
综合各目、科、属的物种分布与生物地理区边界的联系,探讨可能阻碍不同物种扩散的主要影响因素。长鼻目、奇蹄目和灵长目的大中型物种很少,且呈局部型分布特点(图6),各物种基本分属不同的生物地理区,无明显跨区现象,反映本研究识别的生物地理区分界线可以较好的表征对这3个目动物扩散的阻隔效应。与生物区边界相接的奇蹄目动物(蒙古野驴Equushemionus、西藏野驴Equuskiang和野马Equusferus)体型较大,一般栖息于山地草原、荒漠半荒漠地带,分布边界主要受地形障碍和气温季节性变动系数CV影响,表明其可能难以跨越地形起伏剧烈或者气温突变区域。对栖息于温带针阔林的川金丝猴来说,人类活动和气候突变可能是限制其边界扩张的关键因素。

图6 长鼻目、奇蹄目和灵长目大中型物种分布
食肉目中大中型动物在全国都有分布,共11种(图7)。整体上,猫科、熊科和大熊猫科动物分布边界与生物地理区边界较为接近。金钱豹(Pantherapardus)、云豹(Neofelisnebulosa)、黑熊、马来熊(Helarctosmalayanus)以及大熊猫(Ailuropodamelanoleuca)基本分布在生物地理区1,这些物种的扩散主要受气温突变和人类活动影响限制。喜寒物种猞猁、雪豹和棕熊主要分布在生物地理区域2即青藏高原区域,地形障碍和气候突变是限制该区域种群扩散的主要因素。此外,猞猁和棕熊以及黑熊在我国东北地区也有分布,其南向扩张受限与人类活动密切相关。相比而言,犬科动物分布更为广泛,相比自然因素,人类活动的影响可能是其在我国东部地区间断分布的主要原因。豺在我国境内的分布范围较小,整体上分布边界与生物地理区分界线也较为接近,东部地区扩散主要受人类活动影响,西部地区扩散更多受限于气温突变和地形障碍。
我国鲸偶蹄目的大中型物种较多,涉及鹿科、骆驼科、牛科、猪科4科(图8)。骆驼科的野骆驼(Camelusferus)是荒漠动物,适应寒冷、高温等极端环境,我国境内主要分布在生物地理区3西部的塔克拉玛干荒漠地区,地形障碍是限制其扩散的主要因素。野猪(猪科)可栖息于森林草地、灌木沼泽等多种多样的环境中,分布范围十分广泛,但其分布边界与生物地理区2的边界较为吻合,表明高寒环境和地形障碍会抑制野猪扩散。鹿科物种分布广泛,栖息在多样的生境中,其中四川梅花鹿、四川马鹿基本沿青藏高原东部边缘即生物地理区1和2的分界线分布。整体上,青藏高原中西部地区大中型鹿科物种极少,表明地形障碍和气候突变是阻碍鹿科动物向青藏高原内部扩散的关键因素。与鹿科类似,多数牛科物种基本分属不同的生物地理区域,岩羊、西藏盘羊(Ovishodgsoni)、中华鬣羚(Capricornismilneedwardsii)等少部分物种在横断山区地带跨区分布,这与横断山区复杂的地形气候特征有关,塔里木鹅喉羚主要分布在生物地理区3,同时也向南延伸至柴达木盆地区域,但并未进一步向青藏高原内部延伸,高寒环境是限制其扩散的主要影响因素。整体上,生物地理区2内外牛科物种差异明显,反映气候突变和地形障碍是牛科物种扩散的关键因素。
4 讨论
中国动物地理的分布传统上分属古北、东洋两界[39]。根据之前的研究,古北界和东洋界边界的位置从20°N跨越至40°N,存在着一定的分歧。Wallace将古北界和东洋界的边界设为30°N[15],而大多数基于专家的方案都遵循了这个提议,只做了轻微的修改。Smith使用哺乳动物的分布数据和系统分类信息与多维标度分析方法将全球哺乳动物分布划分为4个区域和10个亚区[40],古北界和东洋界的边界倾向于40°N。Kreft与Jetz基于全面的哺乳动物空间数据和聚类算法[17],建议该边界应位于更北处的北纬40°,而Proches与Ramdhani则建议在更南处的北纬20°放置这一边界[41]。本研究从大中型陆栖哺乳动物类群入手,利用四叉树数据结构的自适应分辨率算法构建的二分网络可同时识别数据丰富和数据稀疏的区域,使基于Infomap聚类方法探测的生物地理区域更加独立于尺度。基于Infomap Bioregions算法识别出中国大中型陆栖哺乳动物的三大生物地理区域,区域1与区域2的分界线接近于张荣祖[39]所划分的古北界和东洋界边界的位置,区域1与区域3的分界线与Smith[40]划分界线有重叠部分,如图9所示。已有研究通过研究鱼类区系在历史时空的变化过程[42],论证了青藏高原地区与古北区和东洋区在动物地理区划上应具有同等的地位。Holt等通过使用物种分布和系统发育信息,提出在东亚的25°N和40°N之间建立一个新的“中日(Sino-Japanese)领域”[43],包括中国东部大部分地区、青藏高原和日本。青藏高原作为长江、黄河、湄公河的发源地[44],其环境变化的特殊性创造了一个敏感而脆弱的栖息地,形成了许多特有物种。He等认为由于青藏高原的高度屏障和西北的干燥环境,中国东部和中国西部的物种差异更大[45],其研究结果将青藏高原作为一个独立的地理分区。本研究结果表明青藏高原是陆栖哺乳动物三个地理区域的一个独立划分,区域2与区域3的界线与Holt等[43]划分界线有相似部分,且区域1与区域3的分界线也与Holt等[43]划分界线重叠度较高(图9)。此外,本研究划分区域总体上空间分布区位特征与我国三大自然区类似,但区域边界分布存在明显差异。

图9 中国动物地理区划的分布图
物种生存依赖于各类自然地理环境条件。随着包括生产力假说、水-能源动态假说、环境稳定性假说、地史成因假说在内的各种假说的提出[46],越来越多的研究从不同角度揭示物种分布与影响因子、生态及非生态过程之间的关系。气候、地形、景观以及社会过程影响着动物多样性的分布,Medellín认为哺乳动物丰富度主要与年降雨量相关[47],Chi等研究表明海拔与降水量的协同效应最能解释中国哺乳动物丰富度的分布差异[48],林鑫等发现由归一化植被指数、生态系统类型数和气温年较差构成的回归模型对哺乳动物物种丰富度格局有较好的解释率[49],Tucker等证实在人类足迹指数较高的地区、动物活动频率降低[50]。动物地理分布受多种环境因素的共同影响,环境因素是形成动物分布格局的最重要驱动力。生物地理区受多种地理环境因子的影响是普遍共识,现有研究多以讨论动物地理区域的驱动因素为主。Xie等分析中国生物地理区划,认为山脉和河流是构成许多哺乳动物分布分区的主要屏障[51]。He等基于广义线性模型和偏差划分来研究当前气候、过去气候变化、植被和地质过程在动物地理区域形成过程中的作用,结果表明当前气候和地质过程解释了大部分偏差[45]。Antonelli结合以往研究,论述了板块构造、气候和山脉共同塑造了世界陆地生物多样性演变为不同的生物地理区域[12]。然而关于生物地理区域边界的定量研究却鲜见。生物地理区边界可能是表征对大中型陆栖哺乳动物类群具有隔离效应的界线,往往与地理隔离效应相关,这种隔离效应不仅局限于清晰可见的物理屏障,气候屏障和人类活动引起的隔离效应同样对大中型陆栖哺乳动物的空间分布格局具有重要影响。本文基于Infomap Bioregions划分的生物地理区的格局与我国的气候分区和地形分区存在相似之处,反映出大中型陆栖哺乳动物的分布扩散与地形、气候等密切相关,是多种外部环境因素共同作用下产生的结果。本研究未直接选择代表高程、温度、降雨量水平的数值数据,而是从温度、降水和地形的梯度变化角度入手,探究局部区域的气候与地形突变特征,以此反映气候与地形变化对大中型陆栖哺乳动物的隔离效应,进而更好地分析生物地理区边界的驱动因素。基于空间自回归模型的结果表明,相比年平均气温CV与年降水量CV,气温季节性变动系数CV、降水量季节性变化CV更能代表区域季节性气候变化、表征极端天气特征,对生物地理区域边界的影响明显高于年平均气温CV、年降水量CV。通常,大中型陆栖哺乳动物难以在极寒或者干旱等极端气候环境区域生存,这些区域会成为限制物种扩散的关键因素。基于GWLR模型的结果考虑了驱动因素的空间异质性,研究表明生物地理区边界与气候、地形以及人类活动的协同作用有关,生物地理区边界所在区域与温度、降水气候条件的急剧变化、地形起伏度大以及人类活动强度高的区域紧密相关,但不同区域生物地理区边界的各驱动因素的影响程度存在差异。He等认为中国动物地理区域在东、西地区分异的主要决定因素是气候[45],本研究结论与He等的研究结论较为一致,气温季节性变动系数CV是生物地理区1与生物地理区2边界分异的主导影响因子。生物地理区域1和生物地理区域2的分界线与季风区和非季风区界线的分布趋势较为相似;青藏高原西南部的横断山脉地带受季风气候的影响,温暖潮湿,物种丰富度高;而由横断山脉继续向西北方向延伸的青藏高原内陆区域属高山高原气候,表现出高寒干旱的特征,以适应高海拔、过度干旱、严寒和低氧水平的动物群为主[52];GWLR结果表明气温季节性变动系数CV是该区域生物地理边界的主导驱动因子,温度的突然转变是限制该区域内许多大中型陆栖哺乳动物扩散的关键因素。生物地理区域2和生物地理区域3的分界线与青藏高原北部边界线相符,我国西北地区由于干旱严重,主要居住着耐旱物种,受地形障碍影响明显;西北是以地形障碍为主导驱动因子的区域,气温季节性变动系数CV、降水量季节性变化系数CV均与生物地理区边界呈正相关,人类足迹指数的估计系数值较小,表明该区域的生物地理区边界主要与地形障碍和气候突变有关。此外,本文在探讨动物地理区边界的驱动因素时加入了人类影响因子,我国东部区域即生物地理区1与生物地理区3的边界线的主导影响因子是人类足迹指数,反映出人类活动对大中型陆栖哺乳动物分布表现出了显著影响。我国东部及中部区域人口密集,对生态系统的干扰作用较强,人类土地利用方式造成景观同质化、生态系统的功能退化,人类设施破坏了动物栖息地、限制了动物活动,并引起动物行为的变化[50]。由人类活动引起的生境变化,是导致物种生境丧失、自然生境破碎化、形成生境隔离的重要原因[53]。尤其对需要较大栖息地且较高食物能量供给的大中型陆栖哺乳动物的生存带来严峻挑战,人类活动导致的新型景观格局可能会加剧隔离对大中型陆栖哺乳动物空间分布的影响程度。综合物种分布分析来看,本研究探测的生物地理区边界可以较好的表征对大中型陆栖哺乳动物扩散的隔离效应,进而为解释不同类群动物空间分布格局与环境变化之间的关系提供理论支持。
5 结论
基于Infomap Bioregions算法识别出中国大中型陆栖哺乳动物的3个主要区域,不同生物地理区域的物种组成差异显著,其边界对大中型陆栖哺乳动物类群具有隔离效应。利用空间自回归模型分析不同驱动因子对生物地理区边界的影响程度差异,结果表明,除年降水量CV与边界联系不明显外,其余变量均在10%及以下的显著性水平与生物地理区边界呈正相关,生物地理区边界与气候突变、地形障碍和人类活动的协同作用有关。选取五个显著正相关的解释变量,构建GWLR模型分析各解释变量和边界间关系的空间异质性,进而分析我国大中型陆栖哺乳动物的生物地理区边界驱动因素的空间分异特征。生物地理区1与生物地理区3的边界线的主导影响因子是人类足迹指数,生物地理区1和生物地理区2的边界线的主导影响因子是气温季节性变动系数CV,生物地理区2和生物地理区3边界线在祁连山脉一带的主导影响因子主要是气温季节性变动系数CV。
猜你喜欢
英语世界(2023年10期)2023-11-17
儿童时代·幸福宝宝(2021年11期)2021-12-21
科学大众(中学)(2019年3期)2019-05-17
汽车观察(2018年10期)2018-11-06
证券法律评论(2018年0期)2018-08-31
小学科学(学生版)(2018年5期)2018-06-15
奇妙博物馆(2018年12期)2018-04-25
奇妙博物馆(2018年12期)2018-04-25
少儿科学周刊·少年版(2015年1期)2015-07-07
外语学刊(2014年6期)2014-04-18
