
温度、光照强度和盐度对冬青叶马尾藻生长和光合生理的影响
2023-12-25 00:55徐孟杰李青楠姚丹丹邹潇潇黄惠琴鲍时翔宫春光
南方水产科学 2023年6期
徐孟杰,李青楠,姚丹丹,邹潇潇,黄惠琴,鲍时翔,宫春光,朱 军,穆 丹
1.河北农业大学 海洋学院,河北 秦皇岛 066003
2.中国热带农业科学院热带生物技术研究所/海南热带农业资源研究院/海南省热带农业资源保护与利用重点实验室,海南海口 571101
3.佳木斯大学 生物与农业学院,黑龙江 佳木斯 154007
4.中国热带农业科学院海南省海洋生物功能性成分研究与利用重点实验室,海南 海口 571101
海藻场是一种以大型海藻群落为主的典型近岸海洋生态系统,其在维持海洋生物多样性、降低海区富营养化、提高海洋碳汇等方面发挥着重要作用[1-2]。以褐藻门的马尾藻 (Sargassum)、巨藻(Macrocystis) 等作为支撑的海藻场分布最为广泛[3-4]。但随着全球气候变化、近海养殖和过度采收等原因,天然海藻场退化形势严峻[5-7]。为促进海藻场的保育和恢复,日本、韩国等多个国家开展了海藻场内大型海藻多样性及其生长和繁育特性等的研究[8-9]。
马尾藻是海南岛近岸海藻场的重要建群种,其中冬青叶马尾藻 (S.ilicifolium) 作为在海南分布最为广泛且生物量最大的马尾藻之一[10-11],在维持海藻场生态作用方面发挥着重要作用。在马尾藻属中,已有部分与藻场构建相关物种的生长特性、光合生理、营养需求等的研究报道。例如,栾青等[12]开展了不同温度和光照条件对铜藻 (S.horneri) 的生长、光合色素和营养组分影响的研究,发现不同培养条件对生长率、色素积累和褐藻多酚等有显著影响;张建伟等[13]报道了环境因子对瓦氏马尾藻(S.vachellianum) 生长及光合作用的影响,获得了其适宜生长的环境条件;韦莹莹等[14]研究了温度对各个生长时期鼠尾藻 (S.thunbergii) 的生长特性、叶绿素含量、表观光合速率和呼吸速率等生理指标的影响,结果表明不同生长时期的鼠尾藻具有不同的温度适应性。然而,目前有关冬青叶马尾藻基础生物学特性的研究报道很少,仅见少量关于环境因子对其幼苗发育和繁育特性等方面的报道[15-16]。本文以冬青叶马尾藻为研究对象,探究了温度、光照强度和盐度对其生长和光合生理的影响,为冬青叶马尾藻的资源保护和海藻场修复提供了基础数据。
1 材料与方法
1.1 试验材料
从海南省青葛港海藻场 (110°68'E—110°70'E、19°32'N—19°34'N) 采集藻体健康的冬青叶马尾藻,将其及时带回实验室,置于灭菌海水中浸泡数小时,用毛刷去除附着在藻体上的多毛类、甲壳类等动物。在温度 (20±1) ℃ 、光照强度40 μmol·m-2·s-1、盐度32‰和光周期12L∶12D 的条件下暂养7 d,使冬青叶马尾藻的生长状态趋于同步化。然后用手术刀切除藻体的假根,选取长度约10 cm 的幼苗为试验材料。
1.2 试验方法
根据前期预试验结果,设置温度、光照和盐度试验,具体如下:
1) 温度试验:设置6 个温度处理梯度 (10、14、18、22、26 和30 ℃),每个处理3 个重复;其他培养条件为光照强度40 μmol·m-2·s-1,光周期12L∶12D,盐度32‰。
2) 光照试验:设置6 个光照强度梯度 (10、20、40、80,120,140 和200 μmol·m-2·s-1,每个处理3 个重复;其他培养条件为温度22 ℃,光照周期12L∶12D,盐度32‰。
3) 盐度试验:设置6 个盐度梯度 (16‰、20‰、24‰、28‰、32‰、36‰ 和40‰),每个处理3 个重复;其他培养条件为:温度22 ℃,光照强度40 μmol·m-2·s-1,光周期12L∶12D。
每3 d 更换1 次海水,测定特定生长率 (Specific growth rate,SGR) 和光合色素含量等指标,试验周期为25 d。
1.3 测定方法
1.3.1 特定生长率
每5 d 称量一次鲜质量,计算SGR 以反映藻体鲜质量的变化。
式中:RSGR表示特定生长率 (%·d-1);W0表示初始藻体的鲜质量 (g);Wt表示最终藻体的鲜质量(g);t0表示初始天数 (d);t表示试验时间 (d)。
1.3.2 最大光量子产量
用MINI-PAM 测定,样品暗处理适应20 min后,首先提供0.3 μmol·m-2·s-1的测量光,测得最小荧光值 (F0) ,然后提供0.8 s 约4 000 μmol·m-2·s-1的饱和脉冲,测得最大荧光值 (Fm) 。最大光量子产量 (Fv/Fm) 的计算公式[17]为:
式中:QFv/Fm表示最大光量子产量;F0表示最小荧光值;Fm表示最大荧光值。
1.3.3 光合色素含量测定
取0.05 g 藻体剪碎,用2 mL 80% (体积分数)的丙酮溶液避光浸泡24 h,然后定容到10 mL。用756 PC 型紫外可见光分光光度计测定溶液在664、647、639 、630、510 和480 nm 处的吸光度值。
叶绿素a、叶绿素c和类胡萝卜素的质量分数计算公式为:
式中:WChl-a、WChl-c和Wcar分别表示叶绿素a、叶绿素c和胡萝卜素的质量分数 (mg·g-1);A645表示645 nm 处的吸光度值,其他A值类同;V表示提取液体积 (mL) ;N表示比色前稀释倍数;M代表所取藻体质量 (g) 。
1.4 数据分析
使用Excel 2007 软件进行数据处理和绘图,用SPSS 22.0 软件进行单因素方差分析,设显著性水平为P<0.05。数据以“平均值±标准差 ()”表示。
2 结果
2.1 温度、盐度、光照对冬青叶马尾藻生长的影响
由图1 所示,随着培养时间的延长,14~30 ℃下藻体鲜质量逐渐增加,其中22 ℃和26 ℃组的藻体生长最快,而30 ℃组培养15 d 后的藻体鲜质量增长缓慢,藻体开始软化。10 ℃组藻体的鲜质量呈下降趋势,培养15 d 后其叶片严重腐烂,主茎也开始变软、断裂。

图1 不同温度条件下冬青叶马尾藻的鲜质量曲线Fig.1 Curves of fresh mass of S.ilicifolium at different temperatures
不同盐度下藻体的鲜质量随培养时间的延长而增加,其中20‰ 和24‰ 组藻体培养的前15 d 生长速度较快,15 d 后生长速度开始减慢 (图2) 。盐度为16‰ 时,藻体叶片硬化,易脱落;盐度为36‰时,藻体叶片颜色变淡、变硬。

图2 不同盐度条件下冬青叶马尾藻的鲜质量曲线Fig.2 Curves of fresh mass of S.ilicifolium at different salinities
不同光照强度下冬青叶马尾藻鲜质量与培养时间呈正相关 (图3) 。而在140 μmol·m-2·s-1下藻体培养15 d 后,生长速度减慢,叶片呈淡黄色;在200 μmol·m-2·s-1下,藻体几乎停止生长,没有新的侧枝;在10 μmol·m-2·s-1下,藻体培养20 d后,出现掉叶现象,低光照情况下不利于冬青叶马尾藻的生长。
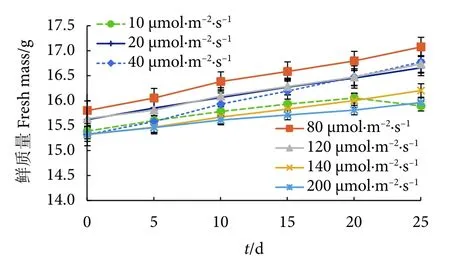
图3 不同光照条件下冬青叶马尾藻的鲜质量曲线Fig.3 Curves of fresh mass of S.ilicifolium at different light conditions
在不同温度梯度 (10、14、18、22、26 和30 ℃) 培养25 d 后,冬青叶马尾藻的SGR 变化见图4-a。随着温度的升高,其SGR 呈先升高后降低的趋势。在6 个温度梯度组中,SGR 在 22 ℃组达到最高 (4.60 %·d-1),其次为 18 ℃组 (3.92 %·d-1)。藻体在 10 ℃组出现负增长 (-2.28 %·d-1),并出现腐烂和变软自溶现象,其SGR 与其他组差异显著(P<0.05)。
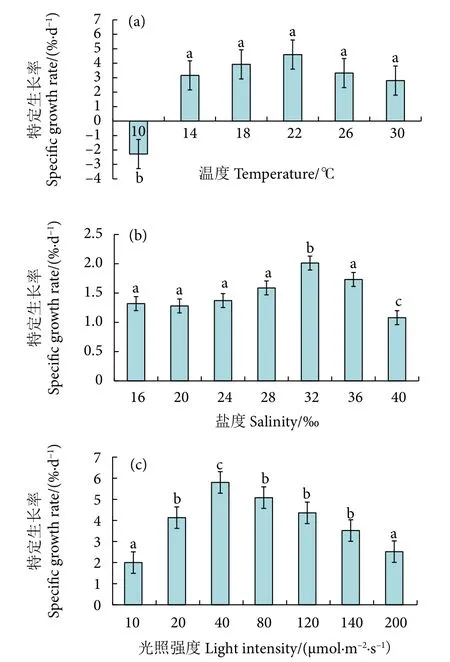
图4 不同温度、盐度和光照条件下培养 25 d 后冬青叶马尾藻的特定生长率注:不同小写字母表示处理间差异显著 (P<0.05);后图同此。Fig.4 Specific growth rate of S.ilicifolium after 25-day cultivation at different temperatures,salinities and light intensitiesNote: Different lowercase letters represent significant differences between different treatments (P<0.05).The same case in the following figures.
在不同盐度梯度 (16‰、20‰、24‰、28‰、32‰、36‰和40‰) 下培养25 d 后,冬青叶马尾藻的SGR 变化见图4-b。随着盐度的增加,SGR 呈先升高后降低的趋势,盐度为32‰时藻体生长最快,SGR 达到最高值[(2.012 ± 0.088)%·d-1],与其他组差异显著(P<0.05)。盐度为40‰ 时,在培养后期藻体出现变软和叶片腐烂的现象,SGR 降至(1.080 ± 0.082) %·d-1,与其他组差异显著(P<0.05)。
在不同光照强度梯度 (10、20、40、80、120、140 和200 μmol·m-2·s-1) 培养25 d 后,冬青叶马尾藻的SGR 变化见图4-c。随着光照强度的增大,SGR 呈现先升后降的趋势。在光照强度为40 μmol·m-2·s-1时藻体生长最快,SGR 为 (5.800 ±0.895) %·d-1,且与其他组差异显著 (P<0.05)。当光照强度从40 μmol·(m2·s)-1逐渐增加到200 μmol·(m2·s)-1时,SGR 呈下降趋势,由 (5.800±0.895) %·d-1降至 (2.520±0.480) %·d-1。
2.2 温度、盐度、光照对冬青叶马尾藻光合色素含量的影响
不同温度下,冬青叶马尾藻光合色素质量分数变化见图5-a。其中,叶绿素a和叶绿素c表现出相似的变化趋势:当培养温度介于10~26 ℃时,两种光合色素随温度的升高而增加,均在26 ℃时达到最高值,分别为 (0.347±0.033) 和 (0.192±0.038) mg·g-1,且与10~22 ℃组的差异显著 (P<0.05) 。另外,类胡萝卜素质量分数虽然也呈现随温度升高逐渐增加的趋势,但各组之间差异不显著(P>0.05) 。
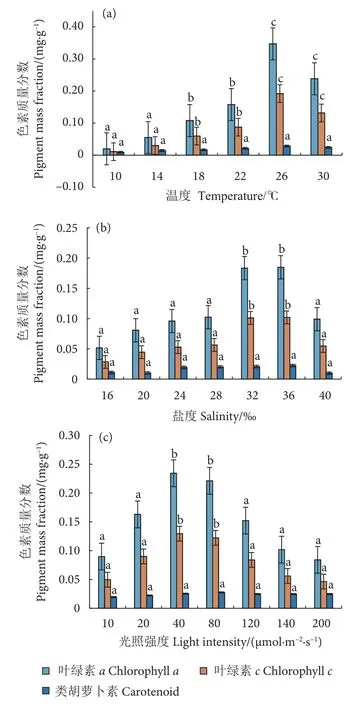
图5 不同温度、盐度和光照强度条件下培养 25 d 后冬青叶马尾藻光合色素质量分数变化Fig.5 Changes of photosynthetic pigment mass fraction of S.ilicifolium after 25-day cultivation at different temperatures,salinities and light intensities
不同盐度下,冬青叶马尾藻光合色素的质量分数变化见图5-b。随着盐度的增加,叶绿素a和叶绿素c均呈先升后降的趋势,最高值均出现在盐度36‰组,32‰组次之,且显著高于其他组(P<0.05) 。当盐度介于16‰~28‰时,各组之间3 种光合色素质量分数差异均不显著 (P>0.05) 。
不同光照强度下,冬青叶马尾藻光合色素质量分数变化见图5-c。随着光照强度的增加,叶绿素a和叶绿素c呈先升后降的趋势,最高值均出现在光照强度40 μmol·m-2·s-1组,80 μmol·m-2·s-1组次之,且这两组显著高于其他组 (P<0.05) 。各组之间的类胡萝卜素质量分数差异均不显著 (P>0.05)。
2.3 温度、盐度、光照对冬青叶马尾藻Fv/Fm 的影响
温度对冬青叶马尾藻Fv/Fm的影响如图6-a 所示。随着温度的升高,Fv/Fm呈先升后降的趋势,10 ℃时Fv/Fm最低 (0.518±0.061),22 ℃时最高(0.757±0.024),但各组之间差异不显著 (P>0.05)。
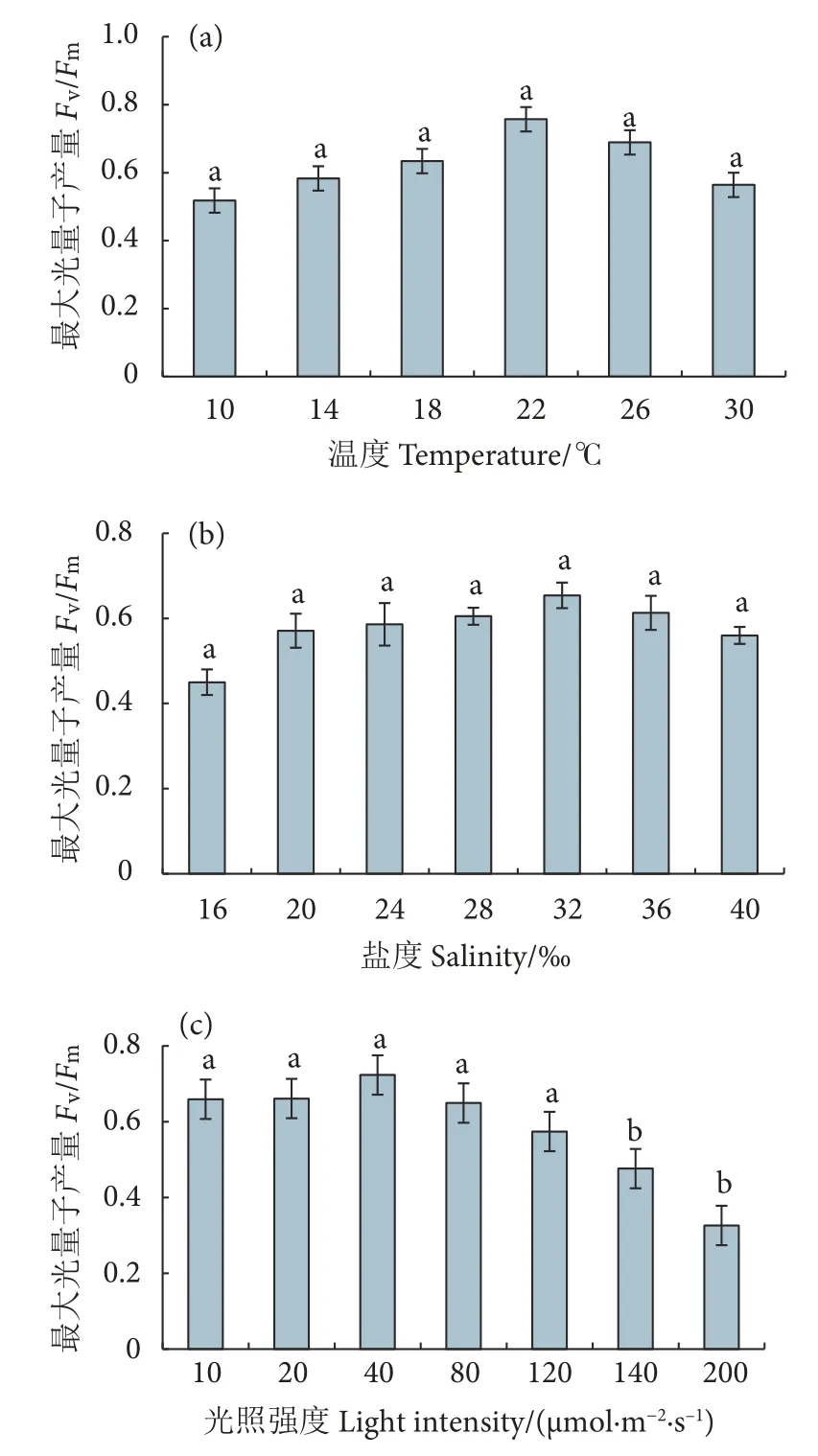
图6 不同温度、盐度和光照强度培养 25 d 后冬青叶马尾藻的最大光量子产量 (Fv/Fm)Fig.6 Maximum quantum yield (Fv/Fm) of S.ilicifolium after 25-day cultivation at different temperatures,salinities and light intensities
盐度对冬青叶马尾藻Fv/Fm的影响较小 (图6-b),随着盐度的升高,其Fv/Fm呈先升后降的趋势,但各组之间差异不显著 (P>0.05)。
光照强度对冬青叶马尾藻Fv/Fm的影响如图6-c所示。光照强度介于10~120 μmol·m-2·s-1时,冬青叶马尾藻的Fv/Fm差异不显著 (P>0.05),最高值出现在40 μmol·m-2·s-1(0.723±0.057);当光照强度高于140 μmol·m-2·s-1时,Fv/Fm开始显著下降(P<0.05)。
3 讨论
3.1 温度对冬青叶马尾藻生长和光合作用的影响
海水温度通过直接影响海藻的光合作用和呼吸作用间接影响藻体的生长和繁殖过程[18]。王永川等[19]发现,潮间带藻类的分布长期受到地理、化学、生物和物理因素的影响。对于大型底栖藻类,许多学者认为温度是最重要的影响因素之一。在本试验条件下,冬青叶马尾藻生长的适宜温度为18~26 ℃,与海南岛另一种常见马尾藻——匍枝马尾藻 (S.polycystum) 的适宜生长温度 (15~25 ℃)和分布范围相近[20];因此,类似的温度适应性,可能是它们分布范围相似的主要原因。本研究发现在22 和26 ℃培养条件下,冬青叶马尾藻生长最快,叶腋处生出分枝,藻体颜色鲜亮且叶绿素a、叶绿素c和类胡萝卜素的含量较高;当温度超过22 ℃时3 种光合色素含量均呈下降趋势,对瓦氏马尾藻[6]和孔石莼 (Ulvaperutsa)[21]的研究也得到类似结果,这可能与过高的温度不利于藻体的叶绿素积累有关。在10 ℃条件下,冬青叶马尾藻因藻体腐烂和变软自溶等现象而出现负增长,这与杨彬等[22]和赵素芬等[23]的研究结果一致,说明10 ℃对冬青叶马尾藻的生长造成了低温胁迫;在高温条件下 (30 ℃) 藻体鲜质量增长缓慢,藻体变软且光合能力下降,这与铜藻受高温胁迫的表现类似[24],暗示温度过高对藻体的胁迫可能对光系统II (PSII)造成了不可逆损伤。虽然低温和高温胁迫均能对冬青叶马尾藻的生长状态和生长速率产生影响,但是通过比较不同温度培养条件下该藻光合色素及最大光量子产量等生理指标的变化,笔者认为冬青叶马尾藻对低温胁迫更加敏感,这与其主要生长在热带海域的生态分布特征相符;因而其可能对低温的适应能力有限,相似的结论近期在对热带海草的低温耐受实验[25]中得到了印证。
3.2 盐度对冬青叶马尾藻生长和光合作用的影响
大型藻类生长与盐度变化密切相关,盐度可直接影响藻类的渗透压、信息传导等[26]。关于盐度胁迫对其他大型藻类的生长及生理生化特性的影响已有报道[27]。本研究表明,随着盐度的增加,冬青叶马尾藻的SGR 和光合色素含量均呈先升高后下降的趋势,最大值出现在盐度32‰ 组;当盐度低于16‰ 时,藻体叶片硬化且易脱落;盐度高于36‰ 时,藻体叶片出现褪色、变软和腐烂等现象,这可能与低盐或高盐胁迫导致其细胞膜结构损伤和产生大量活性氧有关。本研究还显示,随着盐度的增加,冬青叶马尾藻的Fv/Fm未发生显著性变化 (P>0.05) ,这与张玉荣等[24]报道的高盐和低盐胁迫不会导致铜藻幼苗的Fv/Fm发生显著变化的结果相似,该现象可能与马尾藻属物种具有较强的离子传递系统有关[28]。随着盐度的上升,叶绿素a含量呈明显的先升后降趋势,表明过高的盐度会阻断藻体光合色素的合成,从而影响其光合作用过程;而各个试验组的类胡萝卜素含量均无规律性的变化,这与宋新丹等[29]的研究结果相似。不同种类的海藻具有不同的盐度适应范围,本研究表明冬青叶马尾藻的适宜盐度为16‰~36‰,而海黍子(S.muticum) 和悬疣马尾藻 (S.filipendula) 的适宜盐度分别为20‰~35‰[30]和15‰~35‰[31]。此外,也有研究显示,同种海藻在不同地区的盐度耐受范围不同[32],因此,海藻的盐度适宜范围可能与其生活环境密切相关。
3.3 光照强度对冬青叶马尾藻生长和光合作用的影响
光照对于大型海藻的生长发育至关重要。本研究显示冬青叶马尾藻的适宜光照强度为2 0~120 μmol·m-2·s-1,最适宜光照强度为40 μmol·m-2·s-1,过低或过高的光照强度均不利于其生长。低光照强度组 (10 μmol·m-2·s-1) 藻体出现“掉叶”现象,高光照强度组 (140 和200 μmol·m-2·s-1) 藻体生长开始放缓,叶绿素a含量显著降低,藻体叶片呈淡黄色,即出现“失绿”现象;当光照强度达到 200 μmol·m-2·s-1时,藻体出现变软、腐烂等现象,几乎停止生长。杨彬等[22]对莫氏马尾藻的研究也得出相似结果,说明当光照强度高于马尾藻的光饱和点时,可能会破坏光合色素合成,从而抑制藻类生长。
本研究结果显示,在较低的光照强度下,冬青叶马尾藻的最大光量子产量 (Fv/Fm) 随光照强度的增加而升高,但当光照强度超过40 μmol·m-2·s-1时,其Fv/Fm呈明显下降的趋势,表明高光照强度对其光化学活性有明显的抑制作用,这与逄少军等[33]对羊栖菜 (Hizikiafusifarme) 的研究结果一致。但是,也有研究发现一些马尾藻物种对高光强具有很好的适应性,例如海黍子 (S.muticum) 在200 μmol·m-2·s-1依然可以保持较高的特定生长率[34],而铜藻的最适光强可高达300 μmol·m-2·s-1[12],说明不同马尾藻对光照强度的适应范围可能存在较大差异。
综上所述,温度、盐度和光照强度对冬青叶马尾藻的生长和光合生理具有重要影响。冬青叶马尾藻的适宜温度为18~26 ℃,作为一种在热带地区广泛分布的大型海藻,冬青叶马尾藻对低温胁迫更为敏感,这也是限制其分布的主要原因之一。冬青叶马尾藻的盐度适宜范围为16‰~36‰,这与其生长环境的海水盐度密切相关。冬青叶马尾藻的光照强度适宜范围为20~120 μmol·m-2·s-1,过低的光照强度会导致藻体出现“掉叶”现象,过高则会破坏光合色素合成,藻体出现“失绿”或变软、腐烂等现象。因此,在对以冬青叶马尾藻为主的海藻场进行保育和恢复的过程中,应当充分考虑温度、盐度和光照强度对其生长和空间分布的影响,以促进海藻场的有效保护与修复。
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
中华建设(2020年5期)2020-07-24
大自然探索(2019年9期)2019-12-13
小学生必读(低年级版)(2018年6期)2018-09-08
儿童故事画报·自然探秘(2017年10期)2018-03-15
科技创新与应用(2017年20期)2017-07-15
——致坚守奋斗的你
音乐天地(音乐创作版)(2017年1期)2017-02-12
广东海洋大学学报(2015年4期)2016-01-13
中央民族大学学报(自然科学版)(2015年3期)2015-06-11
湖泊科学(2015年4期)2015-02-17
