
牛蛙胚胎发育的形态特征观察
2023-12-25 00:59陈秋羽郑晓婷仲劲松陈智兵梁雪莹董宏标张家松
南方水产科学 2023年6期
陈秋羽,郑晓婷,仲劲松,陈智兵,王 莹,梁雪莹,董宏标,张家松
1.大连海洋大学 水产与生命学院,辽宁 大连 116023
2.中国水产科学研究院南海水产研究所/农业农村部南海渔业资源开发利用重点实验室,广东 广州 510300
3.中洋渔业 (清远) 有限公司,广东 清远 513129
4.广东省农业技术推广中心,广东 广州 510145
牛蛙 (Ranacatesbeiana) 属两栖纲、无尾目、蛙科,其肉质鲜嫩、细软爽口,蛋白质和必需氨基酸含量高且脂质含量较低,加工方便,深受消费者喜爱[1]。牛蛙皮中可提取一种生物活性肽,可作为新型抗菌肽开发原料[2];而牛蛙的脂肪可通过生物技术处理生成一种天然油脂,因其具有优秀的热稳定性和氧化稳定性,在营养食品及制药市场上有很大的潜力[3]。2021 年,我国牛蛙养殖产量超过60 万吨,三产产值近千亿[4]。然而,由于目前我国牛蛙养殖技术落后、种质退化以及对种蛙滥用药物等原因,虽每年投放蝌蚪苗种近20 万尾进行繁育,但仅三成可养成至商品蛙规格[5];因此,高质量苗种培育是关键的技术环节。
在脊椎动物中,两栖纲动物具有独特的变形期,其早期胚胎发育过程及胚后幼体发育过程在苗种培育、比较胚胎学和分子机制研究中均具有重要意义[6-7]。刘楚吾和陈信初[8]于1987 年系统研究了机械振荡、温度、pH、溶解氧对牛蛙受精卵孵化率的影响;王雪虹[9]进一步研究了温度骤变和机械振荡对不同发育阶段牛蛙胚胎的影响。目前,对牛蛙胚胎发育时期的研究仅简单划分了受精卵的发育时期[7],只有手绘发育过程图,并未对其发育过程进行全面划分,也未记录每个发育时期所需的生物积温。因此,本研究通过观察牛蛙的胚胎发育过程,详细记录牛蛙胚胎从合子阶段的受精卵发育至孵化完成阶段的鳃盖闭合期的各时期形态变化特征、发育所需时间及生物积温,充实其影像学资料及生物学资料,旨在为牛蛙的苗种培育提供基础资料。
1 材料与方法
1.1 实验材料
1.1.1 种蛙驯养
牛蛙种蛙取自位于广东省清远市阳山县白莲村的中洋渔业 (清远) 有限公司养殖基地。雌雄种蛙在车间内的水泥养殖池分开驯养,水源为山泉水,24 h 常流水,平均水温为 (21.5±0.5) ℃,溶解氧质量浓度为 (9.10±0.75) mg·L-1,车间内室温为 (26.5±0.5) ℃。每日投喂牛蛙配合饲料2 次,投喂时间为9:00 和18:00,日投喂量为牛蛙体质量的7%~15%。
1.1.2 种蛙选择
选择规格较大、腹部紧致有弹性、四肢形态正常、后腿有劲、眼睛有神、处于发情期且趾端婚垫特征明显的8 只雄蛙作父本,平均体质量为 (723.75±40.90) g;选择腹部膨大柔软圆润、形态正常、皮肤光滑油亮、眼睛有神、轻压腹部能挤压出少许卵粒的8 只雌蛙作母本,平均体质量为 (503.75±23.93) g ;在配对繁殖前3 d 对雌蛙进行禁食,以减少其体内脂肪积累,保证产卵顺利与卵子的质量[10]。
1.1.3 受精卵采集
用100 目筛绢网在种蛙产卵池中分别围出1 m×1 m×1 m 的网兜,网兜远离出水口和入水口。适当调节水深控制在约0.4 m,保持24 h 流水。每个网兜内放入1 对种蛙,其中4 对种蛙成功抱对产卵。在种蛙抱对产卵后,立即用筛绢网捞兜轻轻捞取受精卵,转移到孵化盆,每个盆内放入300 颗受精卵,并把产卵后的种蛙取出放回养殖池中。受精时间从种蛙抱对产出第一批散开在水面的卵后的第5 分钟开始计时。蛙受精卵外有胶质膜,产出后吸水膨大,漂浮于水面;若产出的受精卵未及时捞起,则易下沉不便于采卵;转移蛙卵时避免震动,以免导致受精卵发育畸形[8-9]。
1.2 胚胎发育观察
1.2.1 孵化准备
选取方形塑料盆 (0.54 m×0.39 m×0.32 m) 作为孵化盆,将孵化盆的水位保持在0.2 m。水源皆为山泉水,水温为(24.5±0.4) ℃,光照周期为14 L∶10 D,光照强度为1 000~1 400 lx。pH 和溶解氧的设置参照刘楚吾和陈信初[8]的实验,pH 保持在7.0~7.5,溶解氧质量浓度在7 mg·L-1以上,每12 h 检测水中溶氧量,采用曝气石持续增氧。蝌蚪脱膜前,每天8:00 换水1/3,及时处理胚胎的代谢产物和死掉的胚胎。每组设3 个平行。
1.2.2 胚胎发育观察
用2 mL 的吸管从孵化盆中取10~15 颗受精卵均匀地铺于载玻片上,在光学显微镜(LICHEN XSP-02,10×4 倍) 下观察并拍照,详细记录各时期的发育特征及所需时间。使用高精度电子游标卡尺测量受精卵的径长。每期每组随机取20~30 颗受精卵进行观察,参考谢永广等[11]的实验方法,以40%以上胚胎达到某期确定为该发育时期的起始时间,60% 以上胚胎达到某期确定为该发育时期的结束时间。受精卵在卵裂阶段时,胚胎发育较快,每10 min 观察1 次胚胎发育情况;进入囊胚阶段后,发育速度变缓,每30 min 观察1次胚胎发育情况。生物积温的计算参考邢君霞等[12]的研究,以每期的发育所需时间和平均水温计算胚胎发育该期的生物积温,计算公式为:
式中:K为生物积温(h·℃);T为该阶段发育时的平均水温(℃);n为某一阶段发育至下一阶段所需时间 (h)。
1.3 数据处理与分析
使用Excel 2021 软件对实验数据进行统计,用Adobe Photoshop 2022 软件处理图片。
2 结果
根据实验观察,并参照水产动物的胚胎发育过程[13],可将牛蛙胚胎发育过程划分为7 个阶段。从受精卵开始为合子阶段;受精卵出现倍增性的细胞分裂,为卵裂阶段;伴随细胞的继续分裂分化,胚体形成一个多细胞的囊状胚体,为囊胚阶段;囊胚形成后,由于胚体的原肠作用,细胞会进行内陷和外包,形成原肠胚,为原肠胚阶段;之后,胚体继续发育会形成具有神经管、脊索的胚体,此为神经胚阶段;神经胚阶段后,胚体上逐渐形成器官,此为器官形成阶段;此后至鳃盖完全闭合后,为孵化完成阶段。为了进一步细化牛蛙的胚胎发育过程,参照前人对蛙类的胚胎发育时期划分[7-9,13-14],并结合本实验的观察结果,将牛蛙胚胎发育的7 个阶段再细分为最明显且最具代表性的24 个时期 (图1)。
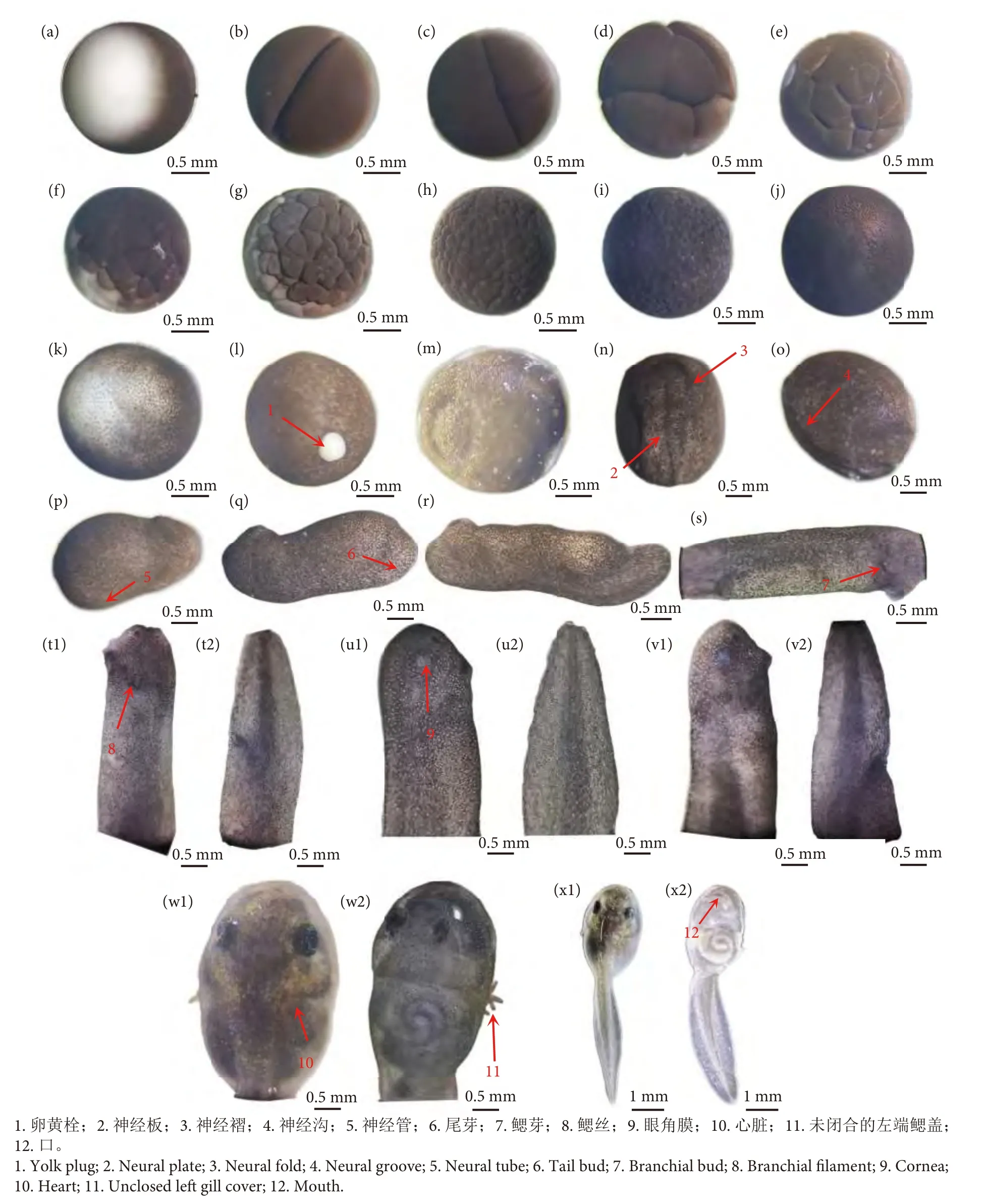
图1 牛蛙胚胎发育分期及各期的形态特征注:a.受精卵;b.2 细胞期;c.4 细胞期;d.8 细胞期;e.16 细胞期;f.32 细胞期;g.64 细胞期;h.囊胚早期;i.囊胚晚期;j.原肠早期;k.原肠中期;l.原肠晚期;m.神经板期;n.神经褶期;o.胚胎转动期;p.神经管期;q.尾芽期;r.肌肉效应期;s.心跳期;t1.鳃血循环期 (头部侧面图);t2.鳃血循环期 (尾部侧面图);u1.开口期 (头部侧面图);u2.开口期 (尾部侧面图);v1.尾血循环期 (头部侧面图);v2.尾血循环期 (尾部侧面图);w1.右端鳃盖闭合期 (头部背面图);w2.右端鳃盖闭合期 (头部腹面图);x1.鳃盖闭合完成期 (背面图);x2.鳃盖闭合完成期 (腹面图)。Fig.1 Developmental stages and morphological characteristics of embryo of R.catesbeianaNote: a.Fertilized eggs; b.2-cell stage; c.4-cell stage; d.8-cell stage; e.16-cell stage; f.32-cell stage; g.64-cell stage; h.Early blastula; i.Late blastocyst;j.Early gastrula stage; k.Mid-gastrula stage; l.Late gastrula stage; m.Neural plate stage; n.Neural folds stage; o.Embryo rotation stage; p.Neural tube stage;q.Tail-bud period; r.Muscular contraction stage; s.Heart beat stage; t1.Gill blood circulation period (Head side); t2.Gill blood circulation period (Tail side);u1.Opening period (Head side); u2.Opening period (Tail side); v1.Tail blood circulation period (Head side); v2.Tail blood circulation period (Tail side);w1.Right gill cover closure period (Head back); w2.Right gill cover closure period (Head abdomen) x1.Gill cover closure period (Back view); x2.Gill cover closure period (Abdomen view).
在本实验中水温为 (24.5±0.4) ℃时,受精卵发育至肌肉效应期需要67 h 49 min,总积温为1 896.95 h·℃;到鳃盖闭合期共耗时177 h 38 min,总积温为4 249.71 h·℃。整个发育过程呈现前期发育速度较快、后期较慢的趋势。胚胎发育各个时期的特征、耗时与积温见表1 和图1。
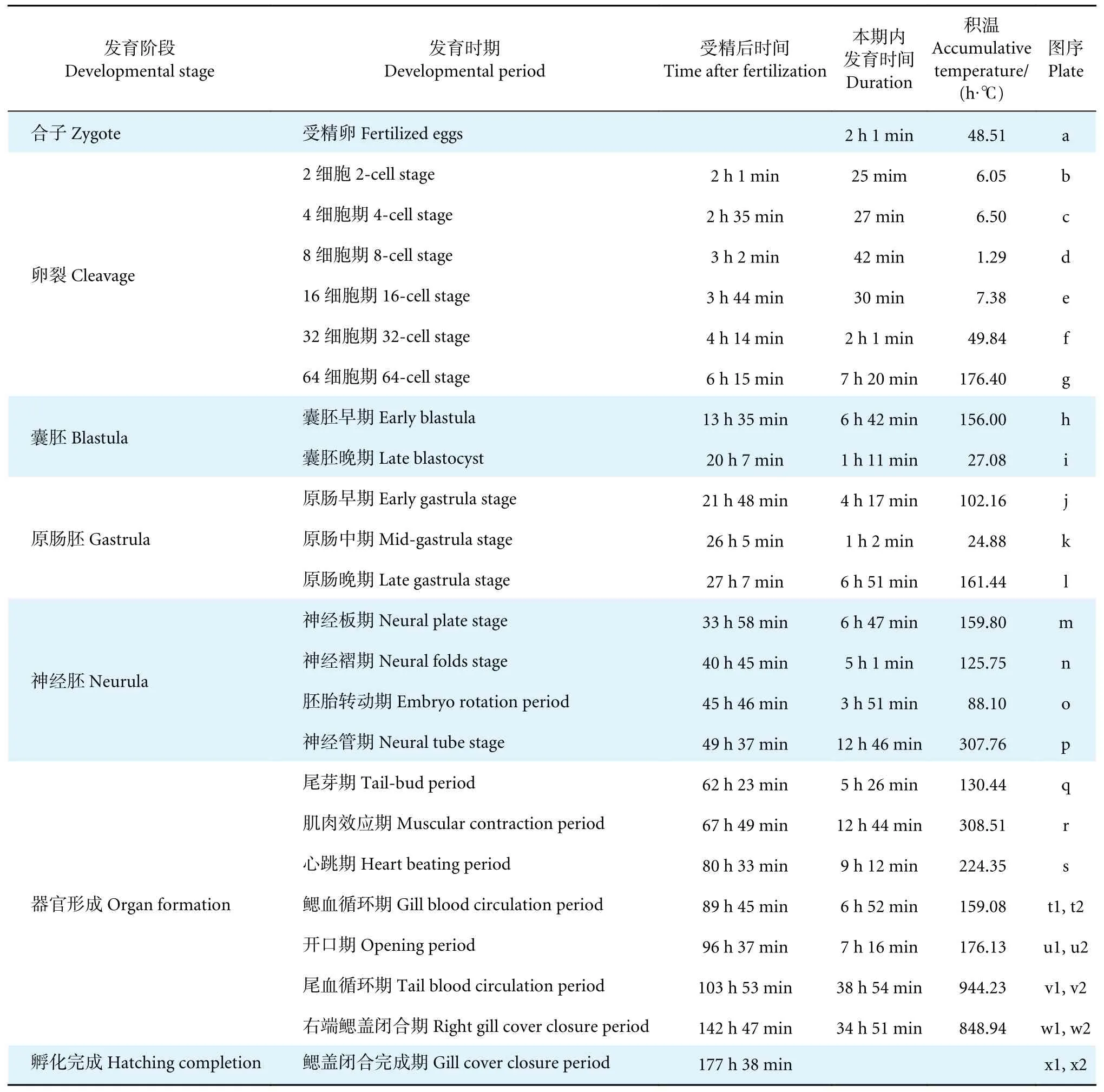
表1 牛蛙胚胎发育分期Table 1 Embryonic developmental stages of R.catesbeiana
2.1 合子
牛蛙受精卵自受精后到第一卵裂沟出现为止 (0~2 h 1 min),此阶段为合子阶段,生物积温为48.51 h·℃。整个卵呈球型,卵径为 (1.41±0.31) mm。牛蛙受精卵属端黄卵,呈极性分布,分为植物极和动物极。植物极卵黄集中,为灰白色,表面积大约占整个卵的1/3;动物极是原生质集中的地方,有黑色素,呈黑棕色。受精卵外围附着一层卵膜,卵膜吸水后膨胀,卵将沉底且能自转,植物极转动朝下,动物极转动朝上 (图1-a)。
2.2 卵裂
牛蛙的卵裂类型为不均等完全卵裂,整个卵裂阶段共耗时2 h 1 min~13 h 35 min,生物积温为304.99 h·℃。自受精2 h 1 min 后,受精卵的动物极出现一条纵沟,被分为不完全相等的2 个卵裂球,为2 细胞期,此时期持续25 min (图1-b);受精2 h 35 min 后,第二条卵裂沟出现,为纬裂,垂直于第一条卵裂沟,受精卵此时分裂为4 个卵裂球,为4 细胞期,此时期持续27 min (图1-c);受精3 h 2 min 后,受精卵出现第3 次卵裂,此时出现的卵裂沟将受精卵分为8 个大小不等的卵裂球,其动物极的卵裂球较植物极的小,此期为8 细胞期,持续42 min (图1-d);受精3 h 44 min 后,第4 次卵裂出现,将受精卵分裂为排列不甚规则且16 个大小不一致的卵裂球,为16 细胞期,持续30 min (图1-e);受精4 h 14 min 后,受精卵进入32 细胞期,经历第5 次卵裂,此次分裂不规则,受精卵被分裂为32 个形状各异的分裂球,持续2 h 1 min (图1-f);受精6 h 15 min 后,受精卵经历第6 次卵裂,被不规则分裂为64 个分裂球,动物极的卵球较植物极的多且小,此期为64 细胞期,持续7 h 20 min (图1-g)。
2.3 囊胚
经历6 次卵裂后,受精卵进入囊胚阶段,此阶段共耗时13 h 35 min~21 h 48 min,生物积温为183.09 h·℃。自受精13 h 35 min 后,受精卵被分裂为大小不等的卵裂球,受精卵表面凹凸不平,有明显卵裂沟,整体呈桑葚状,此期为囊胚早期 (图1-h)。在受精20 h 7 min 后,受精卵进入囊胚晚期,可见其表面界限明显的卵裂沟逐渐变得不清晰,但能分辨出有卵裂沟在受精卵表面 (图1-i)。
2.4 原肠胚
在囊胚晚期后,受精卵进入原肠胚阶段,此阶段共耗时21 h 48 min~33 h 58 min,生物积温为288.50 h·℃。自受精21 h 48 min 后,受精卵表面的卵裂沟已经无法分辨,此时原肠早期的卵裂球更小,且轮廓逐渐模糊;由于其内胚层细胞的内陷,受精卵表面可见有微凹陷部分,微凹陷部分的背侧为背唇,此期为原肠早期 (图1-j)。在受精26 h 5 min 后,受精卵表面变得光滑,只能分辨出有卵裂线不见卵裂沟;且其背唇位置的细胞逐渐下包,植物极细胞内卷,逐渐形成圆形胚孔,胚孔中有白色凸起为卵黄栓,此期为原肠中期 (图1-k)。受精27 h 7 min 后,受精卵随着动植物极细胞的外包和内卷,胚孔逐渐变小,直至被完全包入内部,此期为原肠晚期 (图1-l)。
2.5 神经胚
在原肠胚阶段之后,受精卵进入神经胚阶段,此阶段共耗时33 h 58 min~62 h 23 min,此阶段外胚层特定区域细胞内陷形成神经系统,生物积温为681.42 h·℃。自受精33 h 58 min 后,胚孔封闭,胚体已略变为梨形,侧面明显可见有神经板出现,此期为神经板期(图1-m)。受精40 h 45 min 后,肉眼可见受精卵的整个胚体逐渐拉长,已变为典型的梨形;神经板逐渐隆起形成神经褶,明显可见神经褶中间略有凹陷,此为神经褶期 (图1-n)。受精45 h 46 min 后,神经褶的两侧隆起逐渐向中间靠拢合并,形成神经沟,此时胚胎表面出现纤毛,由于纤毛的摆动可见胚体开始在膜内转动,为胚胎转动期,又称为神经沟期 (图1-o)。受精49 h 37 min 后,胚体继续拉长,神经沟逐渐消失合拢为一条线,从而形成神经管,在胚体上还可见到有尾芽的原基出现,此为神经管期 (图1-p)。
2.6 器官形成
自原肠胚后,胚体继续发育进入器官形成阶段,此阶段经历时间为62 h 23 min~142 h 47 min,生物积温为2 791.71 h·℃。卵受精62 h 23 min 后,胚体头部出现吸盘,鳃区开始逐渐分化出现,且在胚体后背部有明显的尾芽出现,此期为尾芽期 (图1-q)。受精67 h 49 min 后,胚体进入肌肉效应期,此时胚体已完全脱膜,其尾芽更加明显,胚体整体增长,在受到外部刺激时,胚体会做出头尾弯曲的环形收缩反应 (图1-r)。受精80 h 33 min 后,可见胚体尾部发育明显,在头部可见鳃芽出现;在显微镜下可观察到胚体的咽腹部有规则的心跳搏动,此为心跳期(图1-s)。受精89 h 45 min 后,胚体头部侧面有鳃丝长出,鳃丝有明显收缩,在显微镜下可见鳃丝的血管中有血液循环流动;在胚体尾部可见明显体节。胚体在孵化盆中常用吸盘侧吸附在盆壁上不动,仅在受到外界刺激时才会扭曲游动,此期为鳃血循环期 (图1-t1,1-t2)。受精96 h 37 min后,胚体进入开口期,其头部腹面可见口唇轮廓,两腮的鳃丝增长,并可见眼部的角膜变透明 (图1-u1,1-u2)。受精103 h 53 min 后,胚体吸盘逐渐退化;其眼部可见明显的黑色眼球;外鳃基部可见鳃盖褶出现;尾部变透明且有血液流动;腹部变短变圆,在水中可短暂直立,此期为尾血循环期 (图1-v1,1-v2)。受精142 h 47 min后,胚体进入右端鳃盖闭合期,胚体头部变椭圆,可见两个黑色眼球;从背部可见右侧鳃盖已完全闭合,与体表合为一体,左边的鳃丝仍然露出;吸盘已完全退化,腹部可见漩涡状的肠道组织 (图1-w1,1-w2)。
2.7 孵化完成
受精卵历经177 h 38 min,此时已完全孵化为蝌蚪体,蝌蚪体侧的左端鳃盖闭合完成,且外鳃已完全消失;口部上下唇已发育完成,仅留一小孔通水;蝌蚪在水中已能完全自由地游动,并平衡地立于水中,此期为鳃盖闭合完成期 (图1-x1,1-x2)。
3 讨论
3.1 牛蛙受精卵发育阶段与其他蛙类的异同
本研究中,牛蛙的胚胎发育包括合子、卵裂、囊胚、原肠胚、神经胚、器官形成及孵化完成7 个阶段。与棘胸蛙 (Quasipaaspinosa)[11]、黑斑蛙 (Pelophylaxnigromaculatus)[15]、花臭蛙 (Odorranaschmackeri)[16]、凹耳臭蛙(O.tormota)[17]、斑腿泛树蛙 (Rhacophorusmegacephalus)[18]等两栖无尾目动物的胚胎发育大致相近,自受精卵经历卵裂后,发育成囊胚,再自囊胚转化为原肠胚,原肠胚发育成神经胚后开始形成器官,最后孵化完成,成为幼体。此外,结合本实验条件下牛蛙受精卵的发育过程,发现蛙类胚胎发育在经历卵裂阶段的32 细胞期时,卵裂球沟均会由深卵裂沟分化为浅卵裂沟[13,19]。此时表层细胞之间会形成紧密连接,这些连接会显著减少其能量代谢所需的氧气供应,所以此时胚胎表层细胞的发育能影响深层细胞的耗氧[20]。缺氧则会诱导激活其内部细胞的糖酵解[21],而在此时深层细胞可能处于缺氧状态,从而促进内部细胞开始分化,产生胚胎后续一系列的结构变化[22]。可见牛蛙的早期胚胎发育阶段遵循大多数两栖纲无尾目动物的胚胎发育模式。
1937 年,Pollister 和Moore[23]研究了木蛙 (R.Aylvatica) 的胚胎及幼体发育过程,将木蛙胚胎及幼体发育过程划分为23 个时期,却并未对各时期进行命名。Shumway[14]于1940 年参照木蛙胚胎发育时期,将美洲豹蛙 (R.pipiens)的胚胎及幼体发育过程划分为25 个时期,且以每时期最具特征性的表现为各时期命名。二者不同的是,Pollister 和Moore[23]将右端鳃盖闭合期与鳃盖闭合完成期归为了同一时期,而Shumway[14]将这两期单独划分了出来,笔者与其划分方式一致,将牛蛙的右端鳃盖闭合期与鳃盖闭合完成期分为不同的两期[23]。此后,学者们对黑斑蛙、花臭蛙、凹耳臭蛙、斑腿泛树蛙等胚胎发育过程[15-18]皆参照了Shumway[14]的分期方式进行划分。已有部分研究在无尾两栖类动物胚胎发育过程中的鳃血循环期之后、右端鳃盖闭合期之前划分出了鳃盖褶期[8,15-16,23]。本研究与谢永广等[11]、Shumway[14]、刘绍龙等[24]的发现一致,注意到鳃盖褶期常在尾血循环期出现,且出现时模糊。根据胚胎发育分期和命名的原则,即必须以发育过程中显著特征为划分依据,本文未将鳃盖褶期单独划分,鳃血循环期后即右端鳃盖闭合期。
3.2 影响牛蛙受精卵发育耗时的因素
所有生物体都需要以负熵为食,即生物体在吸收外界环境中的热能后,再向外界环境释放热能,以维持其能量代谢。能量代谢是维持生命组织的主要因素,所以人们将生物体的发育与其过程中消耗的能量代谢联系起来[25]。胚胎发育时期的环境温度则是影响胚胎发育形态、时间及发育速度的外部因素[26]。在胚胎发育至幼体的过程中,主要由受精卵中的卵黄提供能量,此过程中,水温升高会提高卵黄中的蛋白降解酶活性,卵黄吸收率和代谢活性随之增加,加快胚胎的发育速度[27]。研究发现,林蛙 (R.chensinensis)[28-29]、木蛙[30]、棘胸蛙[31]等胚胎发育速度与水温呈正相关[32],牛蛙也有类似现象[8-9]。
影响牛蛙胚胎发育耗时的因素除温度外,还有光照条件[33]、养殖密度[34]、盐度[35]等环境因素[36],而受精卵的卵径大小则为影响胚胎发育耗时的内在因素。在水温相似的条件下,饰纹姬蛙 (Microhylafissipes) 受精卵卵径为(0.97±0.02) mm,发育至鳃盖闭合期仅需82 h 36 min[37];本实验中的牛蛙受精卵卵径为 (1.41±0.31) mm,发育至鳃盖闭合完成期需要177 h 38 min,总积温为4 249.71 h·℃;棘胸蛙受精卵卵径为 (4.31±0.69) mm,发育至鳃盖闭合期需要288 h 44 min,总积温5 462.03 h·℃[11]。由此可见,受精卵卵径越大所需发育时间越长,所需生物积温更高,这一规律与大多数鱼类中胚胎发育时间与卵径的关系也相符[38]。这可能由于较大的卵裂球在生长发育中需要更多的营养和更高的生物积温以维持其正常发育[39],而产生这种现象的主要原因还需进一步研究。
3.3 孵化过程中水体环境对牛蛙受精卵的影响
在胚胎的早期发育过程中,溶解氧、pH 等环境因子对胚胎发育的影响极大[40-41]。有研究表明,蛙类胚胎的耗氧量会随着发育时间增加而增加,养殖生产过程中溶解氧含量过低可导致胚胎发育迟缓,甚至幼苗死亡[8,42];而王春青和吕树臣[43]发现当水中溶解氧过高则会引起蝌蚪的气泡病,若不及时救治将引起大量死亡。在受精卵孵化过程中,由于胚胎发育阶段的酸碱调节机制尚未发育完全[44-45],pH 也会显著影响胚胎形态及孵化率[46]。当pH 过高时,会导致云纹石斑鱼 (Epinephelusmoar) 胚胎的孵化率降低[47],这与刘楚吾和陈信初[8]的研究结果一致;而当pH 过低,则会引起海湾扇贝 (Argopectenirradia) 胚胎的畸形率上升[45]。因此本实验在孵化过程中将水中pH 严格控制在7.0~7.5,溶解氧质量浓度在7 mg·L-1以上。在本实验中也尽量避免蛙卵的移动,以免造成牛蛙胚胎的死亡。王雪虹[9]认为机械振动刺激会导致蛙卵的死亡,Xie 等[48]的实验证明慢性机械刺激会造成卵裂球的损害,诱导裂原活化蛋白激酶的磷酸化,从而减缓胚胎的生长。
本实验在孵化牛蛙受精卵的过程中,每日换水1/3 并及时处理水中胚胎的代谢产物和死掉的胚胎。这是由于蛙类胚胎发育过程所产生的代谢废物会导致水体中的氨氮(NH+4)、亚硝酸盐 (NO-2-N) 等含量增加,可能会直接或间接影响胚胎及幼体的发育[49]。且在淡水鱼类的受精卵孵化过程中常见有水霉病,此为卵菌性疾病,会显著影响受精卵孵化的成活率[50-51]。在蛙类受精卵孵化中同样常见,多发生于水质较差、卵块密度太大、水温骤然下降等情况,尤其在连绵阴雨天气的季节最易出现。一旦有卵感染上水霉病,卵四周即会长出肉眼可见的灰白色菌丝,若不及时处理,菌丝则会蔓延至其他正常卵团中,导致大量胚胎死亡。因此,提前做好预防、保持良好的水质和环境对促进胚胎健康发育和提高受精卵成活率至关重要。
4 小结与展望
综上所述,本研究通过观察牛蛙早期胚胎发育的过程及其变化特征,将牛蛙胚胎发育过程划分为7 个阶段24 个时期。在水温 (24.5±0.4) ℃的条件下,胚胎从合子阶段的受精卵发育至器官形成阶段的肌肉效应期需要6 7 h 49 min,总积温为1 896.95 h·℃;至孵化完成阶段的鳃盖闭合期共耗时177 h 38 min,总积温为4 249.71 h·℃。此外,本研究详细记录并补充了牛蛙早期胚胎发育所经历每个时期的变化特征及清晰的影像学资料,可为其高质量育种提供参考依据。
基于本研究的结果,可进一步通过改变胚胎的发育环境[52],研究环境因素对胚胎发育的影响,探究这些环境因素与基因调控之间的相互作用[53-54]。两栖类动物具有较强的再生能力,如非洲爪蟾 (Xenopuslaevis) 在胚胎发育过程中一些特定的组织和器官在被损伤后可再生[55-56]。受此启发,可通过研究牛蛙蝌蚪是否具有再生能力并分析其再生的分子机制和细胞信号通路[57],了解再生在不同情况下的作用机制可为干细胞生物学、细胞可塑性研究提供参考[58]。
猜你喜欢
中国生育健康杂志(2024年1期)2024-01-13
当代水产(2023年2期)2023-05-19
南方医科大学学报(2022年6期)2022-06-30
生殖医学杂志(2021年9期)2021-09-24
当代水产(2021年4期)2021-07-20
当代水产(2019年7期)2019-09-03
天工(2019年7期)2019-01-20
阅读与作文(小学高年级版)(2016年12期)2016-12-27
浙江农业学报(2016年3期)2016-10-31
浙江中西医结合杂志(2015年8期)2015-05-24
