
不同处理对油菜秸秆营养成分、体外瘤胃发酵特性及菌群结构的影响
2023-12-19 08:25魏金涛郭万正樊启文徐志宇
饲料工业 2023年23期
■ 赵 娜 魏金涛 郭万正 樊启文 黄 静 陈 芳 金 枫 徐志宇
(1.湖北省农业科学院畜牧兽医研究所,动物胚胎工程及分子育种湖北省重点实验室,湖北武汉 430064;2.农业农村部农业生态与资源保护总站,北京 100125;3.农业农村部资源循环利用技术与模式重点实验室,北京 100125)
油菜(Brassica campestrisL.)是我国最主要的油料作物之一,但大量油菜秸秆作为农业废弃物被丢弃,既造成资源的浪费也对环境产生压力。油菜秸秆由于适口性差、营养价值低、易霉变等原因,一直以来饲料化利用率低[1-4]。油菜秸秆中含有纤维素、蛋白质、矿物质等多种牛羊生长所需的营养物质,可部分替代常规粗饲料[5],合理开发利用油菜秸秆弥补我国养殖业饲草短缺的现状是国情所需。目前,养殖业尝试使用油菜秸秆作为饲料原料的改良途径主要有发酵、青贮、氨化等,如王亮[6]用微生物处理油菜秸秆提高了锦江黄牛对其粗蛋白、能量和纤维的利用率。陈东等[7]将籽粒苋与油菜秸秆混合青贮,并加入糖蜜、乳酸菌、复合酶等,提高了其粗蛋白、可溶性碳水化合物含量及相对饲用价值。孟春花等[8]和张建丽等[9]采用氨化法降低油菜秸秆纤维含量、提高其粗蛋白质含量及瘤胃体外发酵产气量等,改善了湖羊对油菜秸秆的消化利用率。王亦闻等[10]先进行好氧发酵,再高温蒸煮破坏油菜秸秆表皮纤维结构,然后利用乳酸菌进行厌氧发酵微贮来提高油菜秸秆的饲用品质。油菜秸秆发酵后营养品质明显改善,可部分替代玉米秸秆等粗饲料,提高动物日增重和养分表观消化率。但上述报道的经处理后的油菜秸秆营养价值、动物消化利用率差异较大。体外模拟瘤胃法通过模拟牛瘤胃消化环境、加入瘤胃液维持微生物多样性等方式可达到还原瘤胃消化过程的目的,具有消除动物个体间差异、成本低、高效率等优点[11]。本试验采用高压、酶菌协同发酵等不同处理方式提升油菜秸秆的营养价值,采取体外模拟瘤胃法探究对其体外产气、瘤胃发酵特性、瘤胃菌群结构等的影响,评价其饲用价值,以期为油菜秸秆饲料化利用提供参考。
1 材料与方法
1.1 材料
油菜秸秆:品种为中油杂11,籽实成熟期收获,产地为湖北省咸宁市。
1.2 油菜秸秆的处理
粉碎油菜秸秆(A组):油菜秸秆粉碎后过40目筛。
高压处理油菜秸秆(B组):将粉碎后过40目筛的油菜秸秆用BXM-60VE 立式高压灭菌器在121 ℃0.22 MPa高压处理30 min。
酶菌协同发酵油菜秸秆(C 组):称取粉碎后过40 目筛的油菜秸秆,加入0.5%糖蜜(广西产甘蔗糖蜜)和4×106CFU/g 枯草芽孢杆菌(广州市微元生物科技有限公司生产,下同)、4×104CFU/g 产阮假丝酵母,2×105CFU/g 地衣芽孢杆菌、4×105CFU/g 乳酸菌、2 000 U/g 纤维素酶(南宁庞博生物工程有限公司生产);调节含水率为60%,混匀后用带呼吸阀的PE 袋包装,每袋装500 g,室温(28~32 ℃)堆放发酵,20 d采集样品。
固态二次发酵油菜秸秆(D 组):称取粉碎后过40 目筛的油菜秸秆,加入0.2%米曲霉,调节含水率为60%并混匀后装入玻璃瓶用报纸封口室温放置5 d;之后加入0.5%糖蜜和4×106CFU/g 枯草芽孢杆菌、4×104CFU/g 酵母菌,2×105CFU/g 地衣芽孢杆菌、4×105CFU/g 乳酸菌;调节含水率为60%并混匀后用带呼吸阀的PE袋包装,每袋装500 g,室温(28~32 ℃)堆放发酵,15 d后采集样品。
将采集的各组样品经65 ℃烘干、粉碎过40 目筛后按四分法取出约500 g样品,装入样品袋中,密封干燥保存,用于后续试验。
1.3 体外产气法评价不同加工处理油菜秸秆饲用价值
1.3.1 瘤胃液供体动物
本试验的瘤胃液供体为3 头装有永久瘤胃瘘管、体重(600.0±25.0) kg 的成年黑安格斯阉牛,每日饲喂2 次,共计8 kg全混合日粮(日粮精粗比为5∶5,其中青贮玉米2.2 kg,麦秸1 kg,豆腐渣0.8 kg,混合精料4 kg),自由饮水。
1.3.2 体外发酵试验设计与操作
按照随机试验设计,将体外发酵试验分为5 组,每组设3 个重复,其中空白组不添加底物,作为产气量的校正;A、B、C、D 组分别以油菜秸秆、高压处理油菜秸秆、酶菌协同发酵油菜秸秆、固态二次发酵油菜秸秆样品为发酵底物。
瘤胃液于晨饲前1 h 内采集,使用4 层纱布过滤混匀后39 ℃保温。各组分别称取粉碎过40目筛后的500 mg样品装于150 mL厌氧发酵瓶中,接种50 mL人工唾液[12]和25 mL瘤胃液,通入高纯度氮气后立即盖上胶塞,拧紧瓶盖,连接“AGRS-Ⅲ型微生物发酵微量产气全自动记录装置与软件系统”进行39 ℃恒温体外产气培养,分别读取发酵0、4、8、12、24、48、72 h气体体积,计算不同时间点的体外产气量,并计算累计净产气量。
体外发酵24、72 h 时采集发酵瓶中的发酵底物,测定发酵底物pH,然后将发酵瓶中的发酵底物离心,取1 mL 上清液加入1.5 mL 离心管中,用于测定挥发性脂肪酸浓度;另取4 mL 上清液加入至5 mL 离心管中,用于测定氨态氮浓度。
采集发酵瓶中的72 h 发酵底物,采用16S 高通量测序技术测定发酵底物中菌群结构。采用Illumina MiSeq 平台,利用细菌16S rDNA 基因通用引物343F TACGGRAGGCAGCAG 和798R AGGGTATCTAATCCT,针对细菌16S rDNA基因V3~V4区进行双端测序。
另外同样处理一批样品,在体外培养48 h 后,将发酵瓶取出,迅速放入冰水浴中终止发酵,用气密注射器抽取发酵瓶中的10 µL 气体,立刻用气相色谱仪检测甲烷、氢气、氧氮、二氧化碳等气体成分;将发酵瓶中的消化残渣转入孔径48 µm 的尼龙袋,封口、过滤并漂洗后置于65 ℃烘干,测定主要营养成分的降解率。
1.3.3 体外产气量及降解率计算
产气量(mL)=某时间段内瓶气体产生量(mL)-对应时间段内空白瓶气体产生量(mL)
累计净产气量(mL)=某时间段产气量(mL)-对应时间段3支空白瓶平均产气量(mL)
产气动力学参数计算:
将不同样品在不同时间点(发酵0、2、4、6、8、10、12、16、20、24、30、36、48、72 h)的产气量带入基于Ørskov[13]产气模型,根据非线性最小二乘法原理计算产气参数a、b、c值。
式中:GP——t时刻的产气量(mL);
a——快速产气部分(mL);
b——慢速产气部分(mL);
c——慢速产气部分的产气速度(%/h);
t——时间(h)。
某种营养物质体外降解率(%)=100×(底物质量×某种营养成分含量-残渣质量×某种营养成分含量)/(底物质量×某种营养成分含量)
1.4 检测方法及饲用价值计算
干物质含量的测定参照GB/T 6435—2014,粗灰分含量的测定参照GB/T 6438—2007,粗蛋白含量的测定参照GB/T 6432—2018,总能采用全自动氧弹热量计(Parr 6200,美国)测定,中性洗涤纤维和酸性洗涤纤维采用F-2000 全自动纤维测定仪测定。发酵底物中氨态氮含量采用靛酚比色法;挥发性脂肪酸含量利用气相色谱仪(Agilent 6890N GC system)测定;pH测定采用便携式pH测量仪。
油菜秸秆饲料分级指数(GI)参照以下公式[14]计算:
干物质随意采食量(DMI,%)=120/NDF
可消化干物质(DDM,%)=88.9-0.779×ADF
总可消化养分(TDN,% DM)=-1.291×ADF+101.35
产乳净能(NEL,MJ/kg)=[(1.044-0.011 9×ADF)/0.45]×4.184
饲料分级指数(GI,MJ/d)=NEL×DMI×CP/NDF
1.5 试验数据处理
采用SAS 9.24软件NLIN程序确定指数模型中a、b、c值等参数;试验数据经Excel 2016 初步整理后进行单因素方差分析并采用邓肯氏法里德多重比较。以P<0.05为差异显著性判断的标准。统计结果以“平均值±标准差”表示。
2 结果与分析
2.1 不同处理对油菜秸秆主要营养成分的影响
由表1可知,油菜秸秆(A组)总能值为17.94 MJ/kg、粗蛋白含量5.30%、粗灰分含量7.06%、中性洗涤纤维含量73.36%、酸性洗涤纤维含量52.29%;与A 组相比,B 组总能、粗蛋白、粗灰分、中性洗涤纤维、酸性洗涤纤维含量没有显著变化(P>0.05);C 组和D 组的总能、粗蛋白均显著提高(P<0.05),中性洗涤纤维和酸性洗涤纤维含量显著下降(P<0.05);C 组和D 组间总能、粗蛋白含量差异不显著(P>0.05),D 组的中性洗涤纤维含量和酸性洗涤纤维含量显著低于C 组(P<0.05);C、D 组饲草分级指数显著高于A、B 组(P<0.05),且D组显著高于C组(P<0.05)。

表1 不同处理的油菜秸秆主要营养成分变化(绝干物质基础)
2.2 不同处理对油菜秸秆主要营养成分体外降解率的影响
由表2 可知,与A 组相比,B 组酸性洗涤纤维体外降解率显著提升(P<0.05),干物质、中性洗涤纤维体外降解率并未显著提高(P>0.05),但是B 组粗蛋白的体外降解率仅为25.34%,显著低于其他组(P<0.05);C 组和D 组干物质、粗蛋白、酸性洗涤纤维体外降解率显著高于A 组和B 组(P<0.05),B、C、D 组中性洗涤纤维体外降解率显著高于A 组(P<0.05),其中C 组干物质、中性洗涤纤维、酸性洗涤纤维体外降解率分别比A 组提高了55.29%、43.20%、256.62%;D 组的粗蛋白体外降解率显著高于C 组(P<0.05),达到79.31%,干物质的降解率低于C 组(P<0.05)。

表2 不同处理油菜秸秆主要营养成分的48 h体外降解率(%)
2.3 不同处理对油菜秸秆体外产气和气体成分的影响
由表3 可知,4 个处理组的油菜秸秆0~72 h 体外产气过程中,产气量变化趋势基本相同,即随着体外发酵时间的延长,各处理组产气量逐渐上升,在48 h基本达到了产气平衡状态。其中D 组的12、24、36、48、72 h 体外累计产气量显著低于其他组(P<0.05);12~72 h 内同一时间点,A、B、C 组的体外累计产气量差异均不显著(P>0.05)。

表3 不同处理油菜秸秆的体外累计产气量(mL)
由表4 可知,4 组间快速产气部分、产气速度常数差异不显著(P>0.05);A 组和B 组间快速产气部分、慢速产气部分、潜在总产气量差异不显著(P>0.05);C 组慢速产气部分、潜在总产气量高于其他组(P<0.05),D 组慢速产气部分、潜在总产气量低于其他组(P<0.05)。

表4 不同处理油菜秸秆的体外产气参数
由表5 可知,体外发酵48 h C 组的甲烷含量高于其他组(P<0.05),其他各组间差异不显著(P>0.05);A组和C 组的体外产气二氧化碳占比高于B 组和D 组(P<0.05);4 个处理组氧氮含量差异显著(P<0.05),且D组>B组>A组>C组。
2.4 不同处理对油菜秸秆体外发酵参数的影响
由表6、表7 可知,不同处理油菜秸秆24 h 体外发酵底物总挥发性脂肪酸浓度为D 组>B 组>C 组>A组,且4 组间差异显著(P<0.05)。72 h 体外发酵底物中总挥发性脂肪酸浓度表现A、B 组差异不显著,但显著高于C、D 组(P<0.05)。4 个处理组油菜秸秆经体外发酵24、72 h 后产生的挥发性脂肪酸主要为乙酸、丙酸,且乙酸/丙酸差异不显著。不同处理油菜秸秆的体外发酵底物24、72 h pH 组间差异不显著(P>0.05),但72 h 时pH 均略微上升。不同处理油菜秸秆24 h 体外发酵底物D 组氨态氮浓度为25.17 mg/100 mL,显著高于其他组(P<0.05);与24 h相比,72 h 体外发酵底物中氨态氮浓度除D 组外,均有所上升。
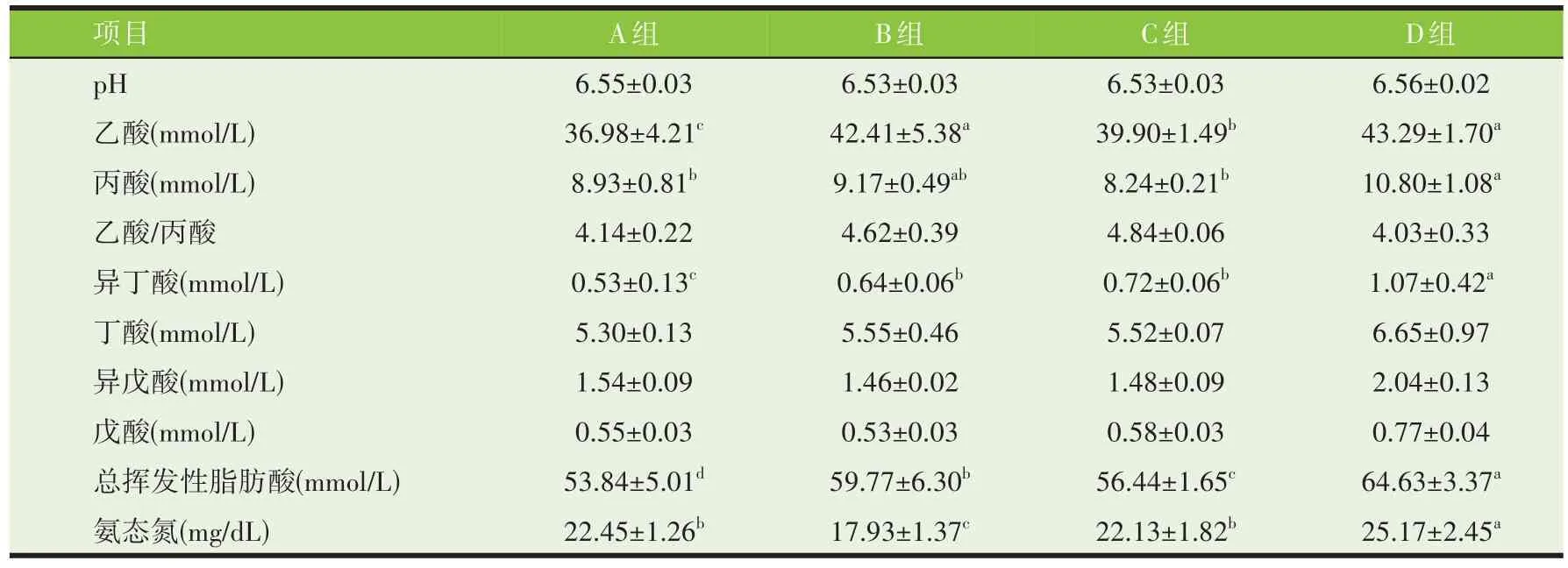
表6 不同处理油菜秸秆的24 h体外发酵参数
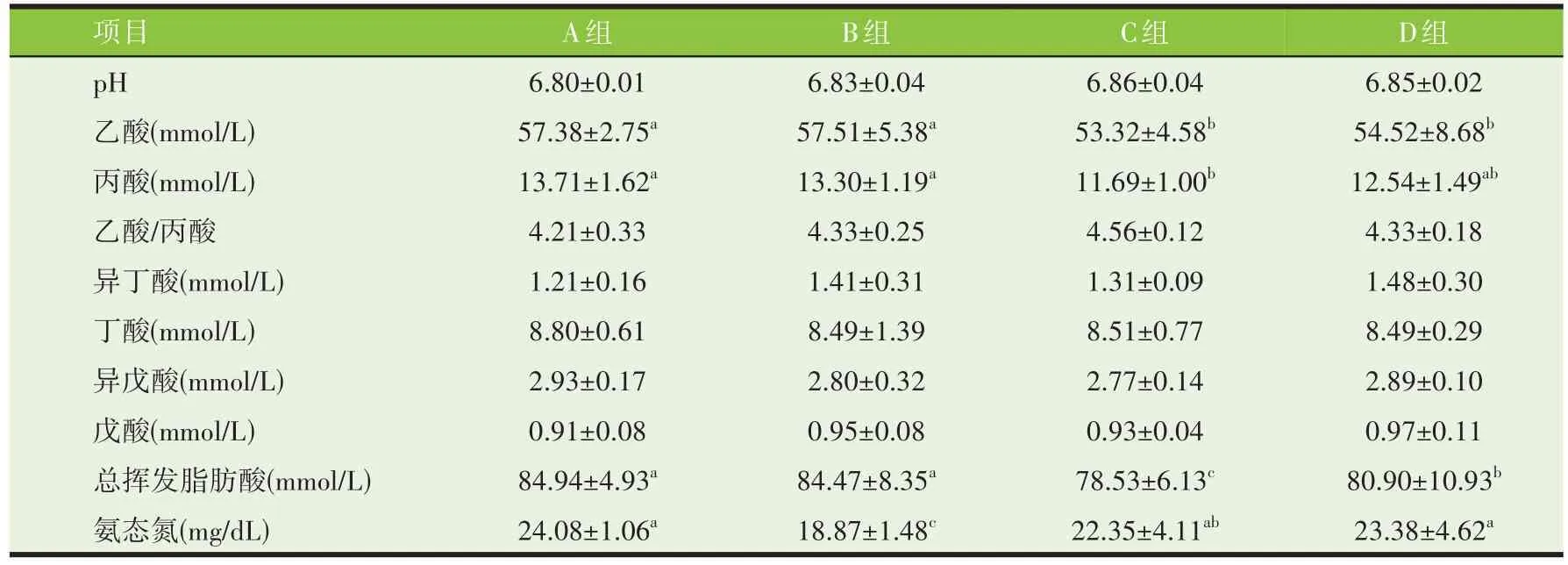
表7 不同处理油菜秸秆的72 h体外发酵参数
2.5 不同处理对油菜秸秆体外发酵底物中菌群结构的影响
本试验采集了4个组的牛瘤胃体外发酵底物共计12个样品。测序下机原始测序数据(raw reads)数据量分布范围为79 063~81 787,在质控之后的高质量测序标签(clean tags)数据量分布范围为32 986~50 949,clean tags经过去除嵌合体得到valid tags(即最终用于分析的数据)数据量分布范围为29 623~46 130,各样本去噪扩增单碱基精度的代表序列(ASV)个数分布范围为503~932。
韦恩图(见图1)反映样品之间共有、特有的ASVs数目,结合ASVs 所代表的物种,样品进行差异统计,采用ANOVA 算法计算出差异ASV 个数为97 个,差异门个数为3 个,差异属个数为23 个。ASVs 数目顺序为A 组>B 组>D 组>C 组,不同处理的油菜秸秆改变了瘤胃内菌群结构。图2 中样品稀释曲线中每条曲线代表一个样本,在本试验的测序深度下,检测到的ASVs 数目不再增加,可以覆盖样品中的大多数微生物。
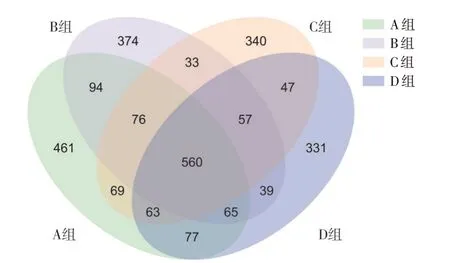
图1 不同处理组间瘤胃菌群ASVs 韦恩图

图2 样品多样性指数稀释曲线
由表8 可知,A 组Chao1 指数显著高于D 组(P<0.05),其他组间差异不显著(P>0.05);物种分布越均匀,香农指数数值越大,A 组香农指数显著高于C 组(P<0.05),A、B 组和D 组香农指数差异不显著(P>0.05),B、C 组和D 组香农指数差异不显著(P>0.05);辛普森指数越高,表明群落多样性越高,A、B 组和D组辛普森指数显著高于C组(P<0.05)。

表8 不同处理油菜秸秆瘤胃微生物菌群Alpha多样性指数分析
采用主坐标分析(PCoA)方法比较各组瘤胃微生物Beta 多样性。图3 为不同处理组瘤胃菌群样本分别在坐标系的不同空间区域分布,主坐标分析结果显示,各组瘤胃微生物区系存在差异。

图3 不同处理油菜秸秆瘤胃微生物菌群PCoA (2D)分析
门水平菌群组成和相对丰度见表9。不同处理油菜秸秆在牛瘤胃体外发酵底物样品中共发现14 个菌门,其中拟杆菌门、厚壁菌门、变形菌门在各组中均为丰度最高的三大菌门,各组瘤胃液中三大菌门的相对丰度总和都超过94%。A 组拟杆菌门的相对丰度(45.60%)显著高于其他组(P<0.05),C 组拟杆菌门的相对丰度(34.77%)显著低于其他组(P<0.05);A 组和D 组的厚壁菌门的相对丰度(34.99%、32.50%)显著高于B、C 组(P<0.05);C 组变形菌门的相对丰度为38.51%,A 组(14.36%)最低,各组间差异显著(P<0.05)。与A 组相比,B 组及C 组牛瘤胃液中拟杆菌门、厚壁菌门相对丰度显著降低,变形菌门的相对丰度显著提高(P<0.05)。可见,油菜秸秆经处理后,可以显著提高牛瘤胃液中优势菌门变形菌门的相对丰度,显著降低了拟杆菌门、厚壁菌门相对丰度。

表9 不同处理油菜秸秆对牛瘤胃微生物门水平相对丰度的影响(%)
属水平菌群组成和相对丰度见表10。不同处理油菜秸秆在牛瘤胃体外发酵底物中共发现199 个属,其中相对丰度排名前10 的分别为理研菌科(Rikenellaceae_RC9_gut_group)、气单胞菌属(Comamonas)、F082 属、拉氏梭菌(Lachnoclostridium)、真杆菌-产粪甾醇真细菌([Eubacterium]_coprostanoligenes_group)、克里斯滕森菌科_R-7 群(Hristensenellaceae_R-7_group)、不动杆菌属(Acinetobacter)、Muribaculaceae属、纤维杆菌属(Fibrobacter)、普氏菌属(Prevotella)。A 组理研菌科(Rikenellaceae_RC9_gut_group)的丰度显著高于C、D 组(P<0.05),与B 组差异不显著(P>0.05);A 组气单胞菌属丰度显著低于其他组(P<0.05)。
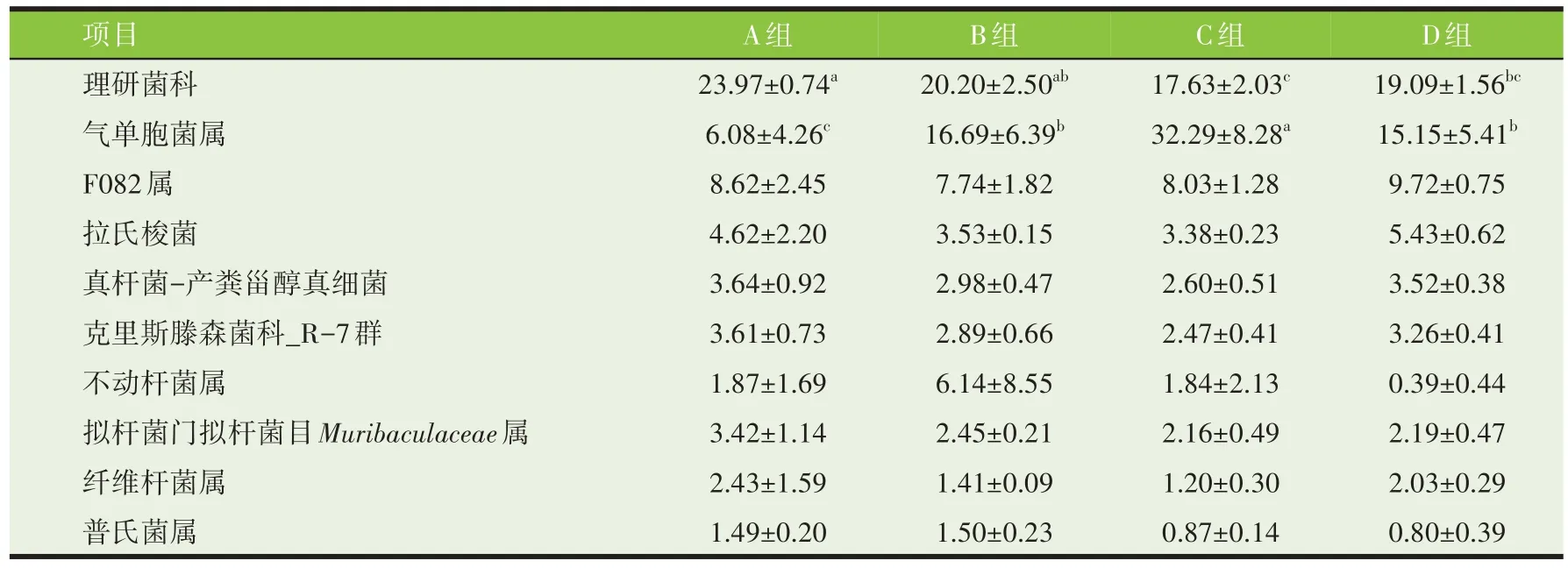
表10 不同处理油菜秸秆对牛瘤胃微生物属水平相对丰度的影响(%)
3 讨论
3.1 不同处理对油菜秸秆主要营养成分及体外降解率的影响
本试验采用的油菜秸秆样品与黎力之等[15]从江西、湖北采集的油菜秸秆总能、粗蛋白、中性洗涤纤维、酸性洗涤纤维、粗灰分含量基本接近。油菜秸秆的总能、中性洗涤纤维和酸性洗涤纤维含量高,粗蛋白含量低,其较高的木质素、纤维素含量导致动物及微生物难以直接有效利用。饲料中性洗涤纤维和酸性洗涤纤维瘤胃降解率反映了粗饲料纤维在瘤胃内消化的难易程度,本试验中油菜秸秆的干物质、中性洗涤纤维、酸性洗涤纤维的体外降解率均为最低。
物理法、化学法以及生物处理等方法常被用来提高秸秆的适口性、营养价值和消化率[16]。本试验中,油菜秸秆经高压处理后主要成分含量没有显著变化;但高温高压可能改变了油菜秸秆的纤维素、半纤维素和木质素的空间链接状态[17],使得其体外酸性洗涤纤维降解率显著提升;同时高温高压条件可能使油菜秸秆的化学成分间发生了美拉德反应,产生难以消化的聚合物,导致其体外粗蛋白降解率较低。
学者们探索了多种微生物、酶等对秸秆的生物预处理效果。真菌菌丝体可破坏秸秆表层结构中难降解的木质素蜡质层[18],龚剑明等[19]采用不同真菌发酵油菜秸秆,纤维得到降解,但同时导致了有机物的浪费。李立波等[20]发现枯草芽孢杆菌可以分泌纤维素胞外酶降解木质纤维素、增加蛋白质含量、提高秸秆的利用率。纤维素酶可增大木质纤维素的孔隙,降低其聚合度,提高反刍动物对粗饲料的消化率[21]。混菌发酵生产菌体蛋白、降解秸秆的效率高于单菌[22]。好氧固态发酵目前在秸秆品质改良中占据主导地位。为提高秸秆饲料预处理效率,多种方法经常联用。本试验采用好氧、混菌等多方法联用的酶菌协同发酵及固态二次发酵处理油菜秸秆。从试验结果来看,油菜秸秆经酶菌协同发酵、固态二次发酵后粗蛋白含量均显著提高,这一方面可能由于发酵过程中添加的产朊假丝酵母、枯草芽孢杆菌等微生物利用了发酵底物中的氮元素,产生了菌体蛋白[23-25];另一方面可能是发酵过程微生物活动消耗了部分有机物质等造成发酵底物浓缩等原因导致的。油菜秸秆经酶菌协同发酵、固态二次发酵后中性洗涤纤维和酸性洗涤纤维含量显著下降,中性洗涤纤维、酸性洗涤纤维的体外降解率也得到大幅提升,且酶菌协同发酵法处理后的油菜秸秆主要营养成分体外降解率较固态二次发酵法提高更为显著,这说明油菜秸秆经酶解发酵处理后,纤维组成有所改变,更易为反刍动物消化利用。
3.2 不同处理对油菜秸秆体外产气量和气体成分的影响
体外产气法通过模拟反刍动物瘤胃发酵过程进行粗饲料营养价值评价,产气量可以反映饲料原料被瘤胃微生物消化利用的情况[26]。Ørskov[27]采用体外产气技术研究不同营养物质在瘤胃的降解时发现,体外产气量与反刍动物干物质采食量、生长速度高度相关。据马绍楠等[28]报道,饲料在肉羊瘤胃中的干物质降解率与体外产气量之间存在一定的正相关。秸秆的木质化程度较高、碳水化合物含量少,决定其产气量整体偏低。本试验中,不同处理的油菜秸秆体外干物质降解率不同,同时在相应的时间内产气量与产气率也有差异;酶菌协同发酵处理组油菜秸秆的干物质降解率和总产气量均高于其他组;与上述前人研究结果不同的是,本试验中固态二次发酵组油菜秸秆的干物质降解率比油菜秸秆、高压油菜秸秆组高,但其体外累计产气量和潜在总产气量却偏低,这可能因为固态二次发酵过程微生物发酵利用了更多的碳水化合物,导致其在瘤胃消化时微生物活性降低,产气量减少。
不同处理组油菜秸秆体外产气气体成分略有差异。二氧化碳是主要温室气体,甲烷是强效温室气体,油菜秸秆由于粗纤维含量高,瘤胃发酵时产生的二氧化碳、甲烷较多,且大部分通过嗳气排出体外,所以在使用油菜秸秆配制日粮时要营养均衡,以降低温室气体的排放。
3.3 不同处理对油菜秸秆体外发酵参数的影响
瘤胃内挥发性脂肪酸主要由微生物发酵碳水化合物和氨基酸等产生,经瘤胃上皮吸收后通过血液循环输送至机体各部,可为反刍动物提供70%~80%的总能量。瘤胃中挥发性脂肪酸的含量和组成比例是分析瘤胃发酵模式的指标,本试验中4 组油菜秸秆经体外发酵后主要产生的挥发性脂肪酸均为乙酸和丙酸,且乙酸/丙酸差异不显著,说明4 种不同处理油菜秸秆在瘤胃中的发酵模式是基本一致的。
在瘤胃内粗蛋白一部分可快速降解成氨基酸和小肽,另外在微生物作用下约有50%会降解为氨。氨是瘤胃内重要的氮供体,可被瘤胃微生物直接利用,也可被瘤胃壁吸收[29]。最适合微生物生长的瘤胃氨态氮临界浓度6.0~30.0 mg/dL,瘤胃液内氨态氮浓度过低,会导致瘤胃微生物生长缓慢。本试验中,4 组油菜秸秆体外发酵底物中氨态氮含量虽有差异,但是均处于最适宜瘤胃微生物生长的浓度;高压油菜秸秆的粗蛋白体外降解率低,同时该组油菜秸秆体外发酵产生的氨态氮浓度也最低,这两者之间可能存在关联性。体外发酵底物中的氨态氮浓度24 h 时固态二次发酵组显著高于其他组,72 h时除固态二次发酵组外均有所上升,这可能是瘤胃消化过程中微生物利用了固态二次发酵油菜秸秆组的大部分氨。
3.4 不同处理对油菜秸秆体外发酵底物中菌群结构的影响
反刍动物主要靠瘤胃中的细菌、真菌和原虫等分解利用纤维素物质。本试验中牛瘤胃体外发酵底物中共发现14 个菌门,其中拟杆菌门、厚壁菌门、变形菌门在各组中均为丰度最高的三大菌门。吴琼等[30]报道,安格斯牛瘤胃微生物以拟杆菌门、厚壁菌门和变形菌门为优势菌群,这与本试验结果一致,说明体外模拟瘤胃系统内的微生物群落维持了基本正常结构。
日粮的营养成分显著影响瘤胃微生物数量。本试验中不同处理导致油菜秸秆中营养成分改变,进而引起牛瘤胃微生物菌群结构发生改变。油菜秸秆组Alpha 多样性指数Chao1 指数、香农指数、Simpson 指数较高,说明油菜秸秆经高压、酶菌协同发酵、固态二次发酵处理后可能会导致瘤胃微生物多样性降低。在属水平上,各组发酵底物菌群中均以理研菌科Rikenellaceae_RC9_gut_group 丰度最高。Cheng 等[31]报道,Rikenellaceae_RC9_gut_group 与丁酸、异丁酸等挥发性脂肪酸含量呈显著正相关性,可能通过影响挥发性脂肪酸的浓度来调节滩羊肉脂肪沉积。但本试验中各组瘤胃发酵底物的Rikenellaceae_RC9_gut_group 相对丰度与丁酸、异丁酸含量并未呈现出明显的正相关。丛毛单胞菌属是各组瘤胃发酵底物相对丰度相对较高的微生物,丛毛单胞菌属可利用木质素[32],但油菜秸秆组丛毛单胞菌属丰度显著低于其他组,可能与油菜秸秆中木质素难以利用有关,其他组油菜秸秆的木质素结构可能得到了改变,更容易被丛毛单胞菌属分解利用,从而引起其丰度的增加。普雷沃氏菌属等被认为是瘤胃中非纤维性碳水化合物的主要降解者,本试验中发酵底物中非纤维碳水化合物含量低,这可能导致了各组发酵底物中普雷沃氏菌属微生物丰度较低。
4 结论
酶菌协同发酵及固态二次发酵处理均能明显改变油菜秸秆营养成分,但酶菌协同发酵更能提升油菜秸秆的营养成分体外降解率、体外累计产气量等指标,在牛瘤胃中发酵效果较好。
猜你喜欢
环境卫生工程(2021年5期)2021-11-20
中国比较医学杂志(2020年4期)2020-05-26
Defence Technology(2019年6期)2020-01-07
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
意林·全彩Color(2019年8期)2019-11-13
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
录井工程(2017年3期)2018-01-22
领导文萃(2017年10期)2017-06-05
