
烟酰胺对泌乳奶牛血液指标、抗氧化及抗炎能力的影响
2023-12-19 08:25南冰禹臧长江李晓斌李云梦仇娅琪杨开伦
饲料工业 2023年23期
■ 南冰禹 李 微 臧长江 李晓斌 刘 拓 李云梦 仇娅琪 杨开伦
(新疆农业大学动物科学学院,新疆肉乳用草食动物营养重点实验室,新疆乌鲁木齐 830052)
烟酰胺(nicotinamide,NAM)和烟酸(nicotinic Acid,NA)是维生素B3的两种不同存在形式,二者均以吡啶为基本结构,在体内可以相互转化,NAM 的物理性质比NA 的物理性质更稳定,且NAM 是血液中主要的存在形式[1]。NA/NAM 具有缓解氧化应激、炎症反应及调控脂质代谢的功能,NA 可以通过抑制关键酶活性发挥抗脂解作用,而NAM 可以通过调节沉默信息调节因子表达量调节脂质代谢,二者可以通过调节细胞自身氧化还原状态缓解机体氧化应激[2]。反刍动物可以通过微生物合成、饲粮供给及色氨酸合成途径获得NA,NAM/NA 可以通过被动扩散的方式或以Na+依赖载体介导的扩散方式被胃、小肠吸收,主要的吸收部位是小肠上端,NA 被肠黏膜吸收后会转化为NAM,与蛋白质在组织中结合,进而形成烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD,辅酶Ⅰ)和烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADP,辅 酶Ⅱ)[3-4]。NA/NAM 在体内主要以NAD 和NADP 两种辅酶的形式参与细胞分解代谢、合成代谢等多种重要反应[5]。在高精料饲粮的育肥的牛羊或高产奶牛的生产中,机体合成B 族维生素不能满足自身需要,进而影响机体营养物的代谢,导致生产性能降低[6]。研究表明,添加NA 可以促进瘤胃微生物生长,增加原虫数量及微生物蛋白产量,进而提高机体对氮的利用率,提高动物生产性能[7-8]。血液生化指标不仅可以反映机体健康状况,同时也可在一定程度上反映动物对饲料养分的利用情况,其值会随机体所处的环境、自身生理状况、营养水平等变化而发生改变[9-10]。血液中抗氧化物酶/氧化产物含量可以反映机体的氧化应激程度,炎症因子/抗炎因子含量可以一定程度上反映机体的受损程度[11-12]。研究发现,围产期补饲NAM有效降低了奶牛血液中三酰甘油含量、非酯化脂肪酸含量及β-羟丁酸的含量[1]。Si 等[13]发现NA 显著下调了炎症因子白细胞介素-6和肿瘤坏死因子-α水平,且灌服NA 后高脂组试验鼠血浆中丙二醛水平显著下降。热应激条件下补饲NAM 后,提高了奶牛血液中免疫球蛋白及超氧化物歧化酶的含量,显著降低了非酯化脂肪酸及三酰甘油的含量[14-15]。本试验给泌乳中后期奶牛灌服不同水平NAM,探究其对泌乳奶牛血液生化指标、抗氧化及抗炎能力的影响,旨在为NAM 在动物生产中的应用提供数据参考。
1 材料与方法
1.1 试验时间及地点
本试验于2021 年8 月—11 月在新疆昌吉市吉缘牧业有限公司开展。
1.2 试验材料
NAM 为白色粉末,纯度为99%,购自新疆天康饲料有限公司。
1.3 试验设计与分组
选择胎次[(2.23±0.62)胎]、泌乳天数[(170±50) d]、产奶量[(36.17±7.40) kg/d]相近且处于泌乳中后期的健康荷斯坦奶牛40头,随机分为4组,每组10头,分别为对照组(CK组)和试验组(NAM7、NAM11、NAM15组),CK 组饲喂基础饲粮,NAM7、NAM11、NAM15 组分别在CK 组的基础上灌服7、11、15 g/d NAM。试验期共75 d,其中预试期15 d,正试期60 d。预先将7、11、15 g/d NAM 分别溶于50 mL 离心管中,加水充分溶解。NAM 组除每日灌服NAM 溶液之外,其余条件与CK 组完全相同,NAM 溶液于每日晨饲时进行灌服。
1.4 饲养管理及日粮组成
试验动物均在同一饲养管理条件下进行,自由采食、自由饮水。试验期泌乳牛饲粮参照NRC(2001)标准进行配制,基础饲粮组成及营养水平见表1。
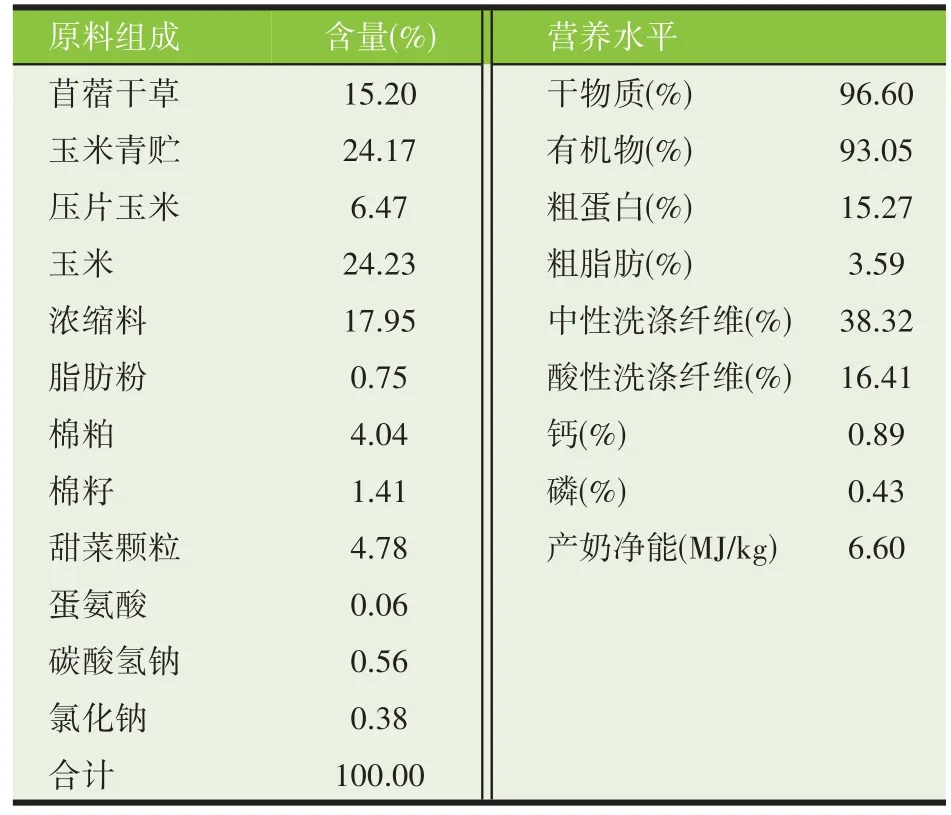
表1 基础日粮组成及营养水平(干物质基础)
1.5 样品的采集与处理
于试验最后1 天晨饲前在牛尾根部采集静脉血,1 份置于含肝素钠的5 mL 采血管中,采集血液样品10 mL,3 500 r/min 条件下离心15 min,吸取血浆并分装至2 mL 离心管中,用于测定血液生理指标;另一份置于5 mL 普通采血管(无抗凝剂)中,采集血液样品10 mL,室温条件下静置30 min 后3 000 r/min 离心10 min,吸取血清并分装至2 mL 离心管中,用于测定血清抗氧化及炎症因子。收集的血清、血浆样品均置于-20 ℃保存。
1.6 测定指标及方法
1.6.1 血液生化指标
使用生化组贝克曼AU5821自动分析仪测定血浆中葡萄糖、尿素氮、高密度脂蛋白、低密度脂蛋白等含量;血浆中非酯化脂肪酸、β-羟丁酸含量采用试剂盒法(比色法)测定,试剂盒购自北京华英生物技术研究所。
1.6.2 血清中抗氧化指标及炎症因子
血清中抗氧化指标及炎症因子的测定方法如表2所示。

表2 血清抗氧化指标及炎症因子的测定
1.7 数据处理
试验数据用Excel 2010 软件进行初步整理,用SPSS 19.0 统计软件进行单因素方差分析,多重比较采用Duncan’s 法进行;试验结果以平均值表示,变异程度用标准误(SEM)表示。差异显著性判定标准为:P<0.01 表示差异极显著,P<0.05 表示差异显著,0.05<P<0.10表示有趋势。
2 结果与分析
2.1 NAM对泌乳奶牛血液生化指标的影响
由表3可知,灌服不同水平NAM对泌乳奶牛血浆中谷丙转氨酶活性、谷草转氨酶活性及碱性磷酸酶活性影响差异不显著(P>0.05)。与CK 组相比,试验组血浆中尿素氮含量显著降低(P<0.05),且NAM7组血浆中尿素氮含量低于NAM11 组和NAM15 组,三组间差异不显著(P>0.05);试验组总蛋白、白蛋白、球蛋白含量均高于CK 组,其中NAM7 组血浆中总蛋白、白蛋白和球蛋白含量均高于NAM11 组和NAM15 组,三组间无显著差异(P>0.05);试验组血浆中葡萄糖含量均显著高于CK 组(P<0.05),其中NAM7 组血浆中葡萄糖含量高于其他两个试验组,但差异不显著(P>0.05);与CK 组相比,试验组血浆中三酰甘油含量显著降低(P<0.05),三个试验组间无显著差异(P>0.05);灌服不同水平NAM 对非酯化脂肪酸、β-羟丁酸、总胆固醇的影响差异不显著(P>0.05)。

表3 NAM对泌乳奶牛血液生化指标的影响(n=10)
2.2 NAM对泌乳奶牛抗氧化能力的影响
由表4 可知,灌服不同水平NAM 后,血清中超氧化物酶歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶含量均高于CK 组,其中NAM7 组血清中三种物质含量均高于CK、NAM11组和NAM15组,但无显著差异(P>0.05);各试验组血清中丙二醛水平均低于CK 组,且NAM7 组相较于其他三组,血清中丙二醛含量最低,但差异不显著(P>0.05)。

表4 NAM对泌乳奶牛抗氧化能力的影响(n=10)
2.3 NAM对泌乳奶牛抗炎能力的影响
由表5可知,灌服不同水平NAM降低了泌乳奶牛血清肿瘤坏死因子-α、白细胞介素-1β、白细胞介素-6 含量,且NAM7 组血清中肿瘤坏死因子-α、白细胞介素-1β、白细胞介素-6 含量低于其他两个试验组,但无显著差异(P>0.05);各试验组血清白细胞介素-10含量均高于CK组,且NAM7组高于其他两试验组,但无显著差异(P>0.05)。

表5 NAM对泌乳奶牛抗炎能力的影响(n=10,pg/mL)
3 讨论
3.1 NAM对泌乳奶牛血液生化指标的影响
机体组织器官的功能以及营养代谢情况,可以间接地通过血液生化指标来反映。谷草转氨酶、谷丙转氨酶以及碱性磷酸酶可以间接反映肝功能健康状况,当肝脏发生异常时,3 种酶活性均会升高[16-17]。本试验中,各组间3 种酶活性均无显著差异,表明灌服不同水平NAM 对肝功能及健康状况无明显影响。血浆中尿素氮、总蛋白、白蛋白以及球蛋白的含量是反映机体蛋白质代谢状况的指标,其中球蛋白还与机体免疫机能相关,尿素氮含量降低说明机体对氮的利用率提高;血浆中葡萄糖可以反映机体糖代谢水平[18]。研究表明,添加过瘤胃烟酸(RPNi)或NA 可以增加血液中葡萄糖的含量,降低尿素氮的水平[19-20];本试验结果与之一致。本试验中,各试验组血浆中尿素氮水平均显著低于CK 组,葡萄糖含量显著高于CK 组;灌服NAM 可能刺激瘤胃微生物生长、增加瘤胃内原虫数量,促进瘤胃微生物蛋白合成或改变微生物蛋白组成,促进碳水化合物向丙酸型发酵模式转变,为机体提供能量[4,6-8];相较于细菌蛋白,原虫蛋白更容易被机体吸收利用,进而提高了机体对氮的利用率[4];同时NAM 还可能增强细胞NADPH 水平,改善细胞质氧化还原的平衡,进而使血中葡萄糖含量上升[21-22]。血液中总胆固醇、三酰甘油、非酯化脂肪酸等含量与脂代谢有关[23-24]。三酰甘油分解是脂肪动员的第一步,脂肪被脂肪酶逐步水解,水解产物为非酯化脂肪酸及甘油,二者被释放入血供其他组织氧化利用。有研究者认为,NA 可以通过阻断双甘油酯酰转移酶-2(此酶能将双甘油酯转化为三酰甘油)发挥作用,从而降低血浆三酰甘油和极低密度脂蛋白水平[25]。对人体的研究表明,NA 通过增加高密度脂蛋白,降低低密度脂蛋白和三酰甘油,有效地解决血脂异常[26]。Yuan 等[27]、刘秀梅[28]研究发现,补充RPNi 或NA 后血浆非酯化脂肪酸浓度降低,本试验结果与之相似。本试验中,NAM显著降低了血浆中三酰甘油的水平,同时各试验组低密度脂蛋白、非酯化脂肪酸、总胆固醇均低于CK组,非酯化脂肪酸及β-羟丁酸也有下降的趋势,但均无显著差异。灌服NAM 可能通过调节有关因子、抑制脂肪分解来调控脂质代谢[29]。
3.2 NAM对泌乳奶牛抗氧化能力的影响
抗氧化能力能够反映机体的健康状况,超氧化物歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶等酶构成了动物机体一线抗氧化防御系统,可防御机体器官和细胞遭受自由基的侵害;自由基的蓄积会对机体产生负面影响,如机体免疫功能降低、体重减轻、细胞生物膜受损、蛋白质变性等[30]。因此氧化功能/抗氧化机制的平衡对机体健康尤为重要。丙二醛是细胞中脂质氧化过程中的最终产物,可以用血液中丙二醛的含量间接反映机体细胞的受损伤程度[7]。超氧化物歧化酶是细胞内重要的抗氧化剂之一,是抵抗机体内超氧化物转化为H2O2的第一道防线,它能缓解细胞因治疗、照射和某些肿瘤引起的损伤[17]。谷胱甘肽过氧化物酶可以清除机体内的自由基,阻止生产脂质过氧化物。NA 和NAM 均被报道有缓解应激的作用。NAM/NA 可以调节线粒体中NAD+/NADH 值,NAD+/NADH值增加(或NADH/NAD+值降低),会使其电子泄漏率降低,同时可以减少超氧化物的生成量,减缓细胞氧化应激状态并提高其抗氧化能力[21,31]。孙先枝等[14]研究表明,NAM 可提高机体血中免疫球蛋白含量、超氧化物歧化酶也有升高的趋势,说明NAM 可以使机体抗氧化能力提升。NA 可以通过增加人主动脉内皮细胞氧化还原电位的途径,达到缓解血管氧化应激的效果,还能通过减少人肝细胞活性氧含量以及降低促炎因子白细胞介素-8含量的方式,发挥抗氧化应激的作用[32]。围产期奶牛补饲NAM 提高了体内抗氧化物酶活性,缓解了奶牛氧化应激[33]。奶山羊在围产期补饲NAM 提高了子代羔羊抗氧化以及免疫能力[29,34]。本研究结果与上述结果一致。本试验条件下,灌服不同水平NAM 后,与CK 组相比,各试验组血清中超氧化物歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶含量,均有升高的趋势。因此,推测奶牛补充NAM 可在一定程度上缓解机体应激状态,NAM 作为NAD+的前体物,可增加细胞NAD+/NADH 值(或使NADH/NAD+值降低),通过调节细胞的氧化还原状态进而调控机体应激状态,提高机体抗氧化能力。
3.3 NAM对泌乳奶牛抗炎能力的影响
免疫细胞和非免疫细胞受刺激会产生细胞因子,细胞因子是一种具有多种功能、可以调节机体免疫应答的蛋白质分子[35]。炎症因子是细胞因子的一种,主要参与炎症反应,机体受到外界刺激时,会产生免疫应答并分泌大量的炎症因子,传递给免疫细胞(如巨噬细胞、淋巴细胞等),使机体进入应激状态,抵制致病菌的侵入,同时也可能进一步释放炎症因子,引发更强的炎症反应。肿瘤坏死因子-α是炎症反应过程中出现最早且最重要的炎性介质,是炎性反应的促发剂。白细胞介素-1β主要由活化的单核-巨噬细胞产生,是促炎性细胞因子,主要通过刺激炎症和自身免疫病相关基因的表达来调节免疫细胞,介导T、B 细胞的活化、增殖与分化。白细胞介素-10 是一种抗炎因子,它可以刺激B 细胞分化增殖,促进抗体生成。Guo等[36]研究发现,NA 可以显著提高奶牛乳腺组织的免疫水平,降低牛奶和血清中白细胞介素-6、肿瘤坏死因子-α和白细胞介素-1β的水平。本试验条件下,NAM降低了泌乳奶牛血清中肿瘤坏死因子-α、白细胞介素-1β、白细胞介素-6 含量,提高了血清中白细胞介素-10 含量。NA 主要作用于GPR109a 和GPR109b 受体,该受体在脂肪细胞和免疫细胞中表达,在脂肪细胞中,GPR109a活化可以减少三酰甘油脂解,减少游离脂肪酸对肝脏的动员,GPR109a可以通过抑制NF-κB信号通路/介导T 细胞达到缓解细胞炎症的目的[37]。Almquist 等[38]研究表明,LMQUISTGPR109a 在乳腺上皮细胞(BMECs)和炎症反应中高表达,激活GPR109a可以显著降低BMECs 中白细胞介素-6、白细胞介素-1β和肿瘤坏死因子-α的表达,从而抑制乳腺炎,因NA 是GPR109a 的配体,亦可以缓解炎症反应。NAM 可以通过抑制活性氧的生成及促炎因子的表达从而缓解炎症[39-40]。孙先枝等[14]研究表明,NAM 可以使血中肿瘤坏死因子-α含量降低。奶山羊在围产期补饲NAM 可减少其血液中促炎因子的浓度,同时提高了子代羔羊的免疫能力[29,35]。NA 能使小鼠肺泡炎症细胞中肿瘤坏死因子-α、白细胞介素-6、白细胞介素-8 的含量减少[41]。因此,推测灌服不同水平NAM,可以通过抑制核因子-κB(NF-κB)通路的方式,减少炎症因子的表达,从而缓解细胞的炎症反应[22]。
4 结论
灌服不同水平烟酰胺可以提高泌乳奶牛血浆葡萄糖含量、降低血浆中三酰甘油及尿素氮的含量,进而提高自身能量转化及利用效率,改善机体氧化应激状态,在实际生产中建议烟酰胺适宜添加量为7 g/d。
猜你喜欢
医学食疗与健康(2021年25期)2021-05-12
现代畜牧科技(2021年2期)2021-03-19
中国生殖健康(2020年2期)2021-01-18
河南畜牧兽医(2020年5期)2020-01-14
河南畜牧兽医(2020年1期)2020-01-11
河南畜牧兽医(2020年21期)2020-01-10
河南畜牧兽医(2020年21期)2020-01-10
中国生殖健康(2018年2期)2018-11-06
农民致富之友(2018年18期)2018-07-14
中华老年口腔医学杂志(2016年6期)2017-01-15
