
基于F 基因的鸽新城疫病毒分子特征分析
2023-12-09 14:26苗书魁魏玉荣
现代畜牧兽医 2023年11期
汪 萍,苗书魁,魏玉荣,夏 俊
( 新疆畜牧科学院兽医研究所,新疆 乌鲁木齐 830011 )
新城疫是由副黏病毒科禽腮腺炎病毒属禽副黏病毒Ⅰ型(APMV-1,也称新城疫病毒)引起的病毒病,在我国被列为二类动物疫病[1]。新城疫病毒(NDV)不断进化共产生21 种基因型,可感染240 多种鸟类,在遗传、毒力、抗原性和寄主范围等方面具有高度多样性[2]。1975年,新城疫第三次大流行从鸽子开始,传播到世界各地[3-4]。鸽新城疫于20 世纪80 年代传入我国,APMV-1 基因型Ⅵ是造成这一流行病的主要原因,也称鸽副黏病毒-1(PPMV-1)[5]。NDV 是一种负链RNA 病毒,NDV 基因组可编码6 种结构蛋白:核蛋白(NP)、磷蛋白(P)、基质(M)、融合蛋白(F)、血凝素-神经氨酸酶(HN)和大蛋白(L)以及两种非结构蛋白V和W,其中定位于病毒囊膜的F蛋白通常被认为是NDV毒力的分子标记[6]。随着新疆畜牧产业结构调整,养鸽业已成为和田地区的主导产业之一,除了采取对规模化养殖场鸽子进行疫苗免疫的预防措施外,还需了解鸽新城疫流行现状及病毒的分子特征。本试验以新疆和田地区规模化养鸽场流行病学调查阳性样品为研究对象,进行基于F基因的鸽新城疫病毒分子特征分析,为制定系统、有效的鸽新城疫防控策略提供参考。
1 材料与方法
1.1 试验材料
2020—2022年,本实验室分别从新疆和田地区6个县市的规模化肉鸽养殖场采集81份临床症状疑似感染NDV的鸽子肺和脾脏组织,冷链运输至实验室-20 ℃保存。
1.2 主要试剂
RNA 提取试剂盒购自QIAGEN 公司;One step RTPCR kit 购自Takara 公司;DNA 凝胶回收试剂盒购自AXYGEN 公司。F基因特异性引物序列为5'-AGGCACCCAACGTGCTGTCG-3';5'-ACGGAGACTCAAGGGCCACC-3'[2],由生工生物工程(上海)股份有限公司合成。
1.3 RNA提取及RT-PCR扩增
组织样品用无菌剪刀剪碎,于组织匀浆器中充分研磨后置于含抗生素的PBS(0.01 mol/L、pH 值7.0~7.4)中,制成浓度为10%的混悬液,冻融3 次,8 000 r/min 离心10 min,取上清提取病毒RNA。按QIAGEN RNeasy MINI Kit 说明书提取病毒RNA,利用One step RT-PCR kit 进行RT-PCR,取2 μL提取的RNA进行RT-PCR。
反应体系:PrimeScript 1 Step Enzyme Mix 1 μL、上下游引物各0.5 μL(20 μmol/L)、2×1 Step Buffer 12.5 μL、Template RNA 1 μL、RNase Free dH2O 9.5 μL。
反应条件:50 ℃反转录30 min;94 ℃ Taq 酶激活2 min;94 ℃ 30 s, 57 ℃ 30 s,72 ℃ 1 min,30 个循环;72 ℃延伸10 min。RT-PCR产物经1.5%琼脂糖凝胶电泳,样品出现约535 bp目的片段判为NDV核酸阳性。统计阳性结果,计算各地区阳性率。
1.4 F基因扩增与序列分析
将RT-PCR 鉴定阳性样本,取5 μL 提取的RNA 于50 μL反应体系进行RT-PCR扩增。
反应体系:PrimeScript 1 Step Enzyme Mix 2 μL、上下游引物各1 μL(20 μmol/L)、2×1 Step Buffer 25 μL、Template RNA 3 μL、RNase Free dH2O 18 μL。
反应条件:50 ℃反转录30 min;94 ℃预变性2 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,35 次循环;72 ℃延伸7 min。RT-PCR 产物经1.5%琼脂糖凝胶电泳,出现约1 600 bp目的片段判为F基因扩增阳性。凝胶回收后送生工生物工程(上海)股份有限公司测序。
使用Lasergene7.0 软件中Clustal W 和MegAlign 功能进行同源性分析,MEGA5.0软件进行遗传进化分析,NCBI在线工具MSA Viewer比较不同F基因的分子特征。
2 结果与分析
2.1 规模化养鸽场鸽新城疫流行病学调查结果
和田地区6 个规模化养鸽场中选择出现粪便稀薄,呈黄绿色或黄白色,扭颈、瘫痪等神经症状的病鸽,采集病料进行鸽新城疫病毒检测。
RT-PCR结果显示,81份样品中有12份样品扩增到目的片段,和田地区鸽新城疫阳性率14.81%(12/81),其中县市1 阳性率11.77%(2/17),县市2 阳性率11.11%(1/9),县市3阳性率18.75%(3/16),县市4阳性率15.79%(3/19),县市5阳性率11.11%(1/9),县市6阳性率18.18%(2/11)。部分样品RT-PCR产物电泳结果见图1。
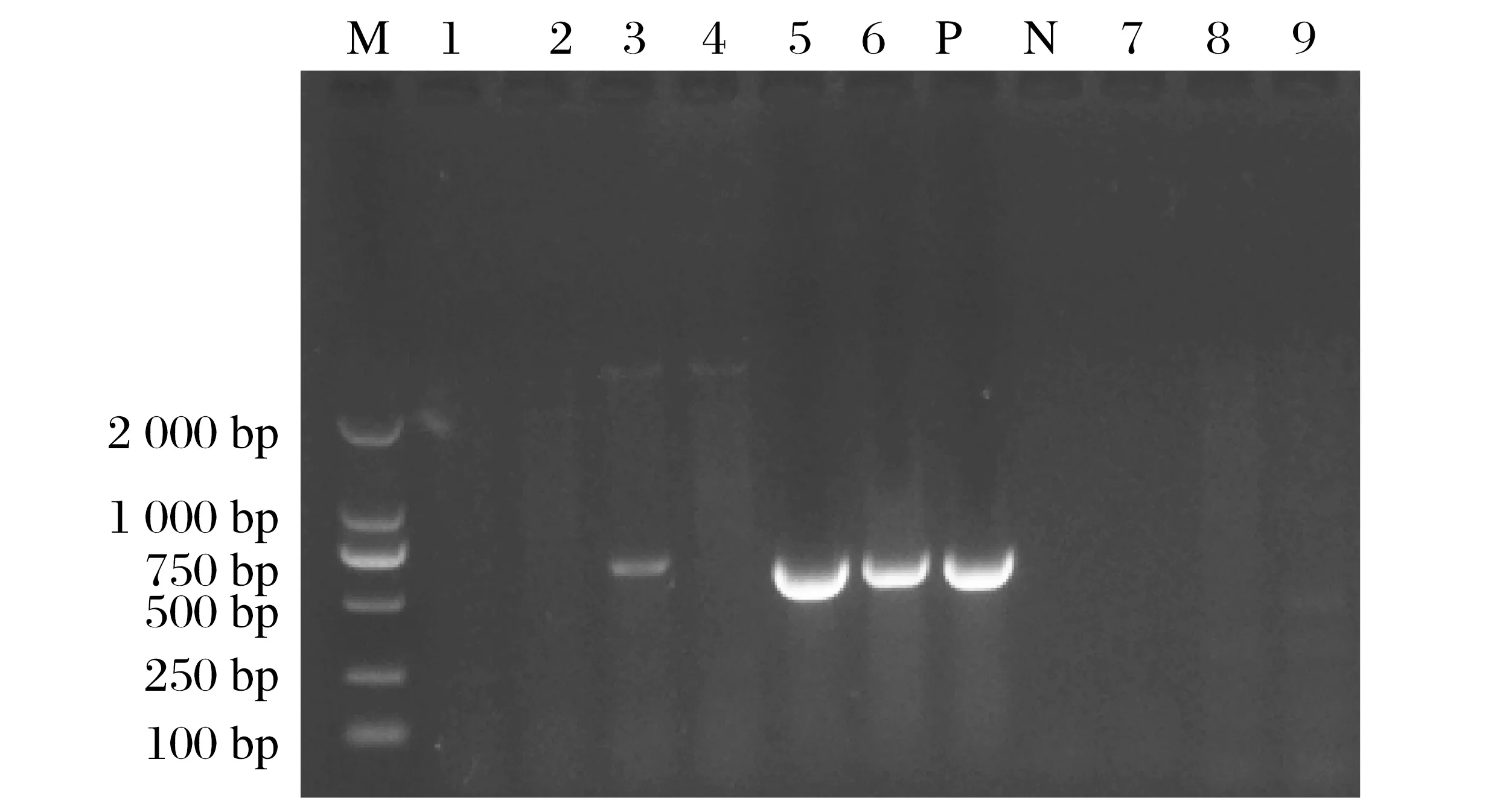
图1 部分样品RT-PCR产物电泳结果Fig.1 Electrophoretic results of RT-PCR products of some samples
2.2 基于F基因的核苷酸同源性分析(见图2、图3)
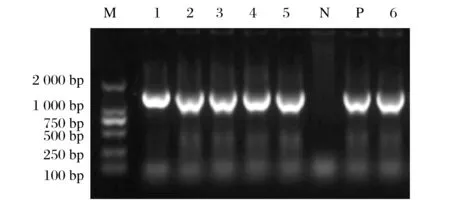
图2 F基因RT-PCR扩增结果Fig.2 RT-PCR amplification results of F gene

图3 不同毒株F基因核苷酸同源性分析Fig.3 Nucleotide homology analysis of F gene in different strains
从流行病学调查阳性样品中选择分别代表不同县市和养鸽场的6株F基因扩增产物进行纯化后测序,见图2。
由图2可知,所测6株序列与GenBank中PPMV-1的F基因序列一致,分别命名为:NDV/PIGEON/XJHT/446/2020/F、NDV/PIGEON/XJHT/467/2020/F、NDV/PIGEON/XJHT/491/2020/F、NDV/PIGEON/XJHT/472/2021/F、NDV/PIGEON/XJHT/802/2022/F 以及NDV/PIGEON/XJHT/812/2022/F。上述6株分离毒株与国内外11株F基因核苷酸序列进行同源性分析,结果见图3。
由图3 可知,6 株F基因之间核苷酸同源性为97.7%~99.9%,与甘肃、宁夏来源的分离株核苷酸同源性为97.3%~99.5%,与国内其他省份来源的分离株核苷酸同源性为97.2%~98.6%。6 株分离株F基因与基因Ⅱ型疫苗株LaSota 株、HB1 株、VG/GA 株、Clone30 株核苷酸同源性为83.5%~84.7%,与基因Ⅲ型Mukteswarz 株核苷酸同源性为84.9%~85.7%。
2.3 基于F基因的遗传进化分析
分别与GenBank 中43 株代表不同基因型PPMV-1 的F基因进行核苷酸序列比对,并构建进化树,结果见图4。
由图4 可知,所测6 株F基因序列均属Ⅵ.2.1.1.2.2,其中NDV/PIGEON/XJHT/802/2022/F 与分离自宁夏的MG840654.1 亲缘关系最近,同在一个小分支内;而其他5 株F基因(NDV/PIGEON/XJHT/446/2020/F、NDV/PIGEON/XJHT/467/2020/F、NDV/PIGEON/XJHT/491/2020/F、NDV/PIGEON/XJHT/472/2021/F、NDV/PIGEON/XJHT/812/2022/F)均在同在一个小分支内;6 株F基因序列均与甘肃的OM640464.1 亲缘关系较近,与基因Ⅱ型疫苗株亲缘关系最远。
2.4 F蛋白主要功能区的分子特征分析(见表1)

表1 6株F蛋白功能域中氨基酸突变Tab.1 Amino acid mutation in functional domain of six F protein
由表1 可知,对比6 株F 蛋白和10 株国内流行株主要功能区域的分子特征,结果显示,F 蛋白裂解位点序列均为112R-R-Q-K-R-F117,6株分离株F蛋白N端信号肽区的第4、5位点有苯丙氨酸、苏氨酸的插入,第6位点丝氨酸突变为酪氨酸。6株分离株F蛋白融合肽区均有两处氨基酸突变(V121I、A132S),HRa 区均有1 处氨基酸突变(V179I),其中NDV/PIGEON/XJHT/802/2022/F 在HRb、HRc 有一个氨基酸突变(L298F 和K483R),在跨膜区有2个氨基酸突变(V509I、V512I)。
3 讨论
除APMV-1基因型Ⅵ外,APMV-1基因型ⅩⅪ也会引起鸽新城疫暴发。2017 年,巴基斯坦鉴定出APMV-1 基因型ⅩⅪ.1.1 和ⅩⅪ.1.2[7]。2011—2015 年,赵振振[8]从我国部分地区分离到基因Ⅵb 亚型鸽新城疫病毒11 株。2021—2022 年,彭真奇等[9]从不同地区鸽群中分离到8 株NDV 代表株,均属于基因Ⅵ型;Zhan 等[10]通过对186 株中国PPMV-1分析证实目前中国亚基因型Ⅵ.2.1.1.2.2最多,其次是亚基因型Ⅵ.2.1.1.2.1、Ⅵ.2.2.2和Ⅵ.1。本试验中的6 株鸽新城疫流行株鉴定为亚基因型Ⅵ.2.1.1.2.2,与目前APMV-1 基因Ⅵ亚型在我国鸽群中流行的特征相符。综上,Ⅵ.2.1.1.2.2目前是我国鸽新城疫主要流行基因型。
据报道,融合肽和HR区的氨基酸替代或NDV跨膜结构域的替换可能会影响F蛋白的融合活性,进而影响病毒的致病性[11]。本试验中6株F蛋白的主要功能区域均发生了点突变,其中NDV/PIGEON/XJHT/802/2022/F 在HRb、HRc和跨膜区出现1~2个氨基酸突变。
在不断开发新城疫疫苗的背景下,鸽NDV 的进化选择压力增加。有研究报道,NDV的所有编码基因和一些非编码区中均发现了来自疫苗谱系和循环病毒谱系的自然同源重组,活疫苗株可能通过与循环病毒的同源重组在NDV 进化中发挥作用[12-13]。有研究发现,F 蛋白裂解位点序列112R-R-Q-K-R-F117虽然具有强毒株的特征,但NDV的毒力指标如鸡脑内接种致病指数低于0.7。Zhan等[10]在2020年从江苏、安徽和河南省的病鸽中鉴定出7株PPMV-1毒株,其F蛋白切割位点序列是典型强毒特征,但根据鸡胚最小致死量的平均死亡时间(MDT)和脑内致病指数(ICPI)值分析,这7 株毒株属于中等毒力。裴育等[14]从北京、天津、山东鸽群分离的4 株鸽NDV F 蛋白切割位点序列是典型强毒特征,但对鸡的致病性显示出中等毒力。因此,进行鸽新城疫致病性分析时应结合其他毒力判定指标进行综合判定。
目前,鉴于无鸽源新城疫疫苗的现状,鸡新城疫疫苗被广泛应用于鸽新城疫预防。调查数据显示,近年和田地区规模化养鸽场主要应用鸡新城疫、禽流感(H9亚型)二联灭活疫苗(LaSota株+SZ株)、鸡新城疫活疫苗(LaSota株)、鸡新城疫活疫苗(CS2 株)、新城疫灭活疫苗、重组新城疫病毒灭活疫苗(A-Ⅶ株)防控鸽新城疫,鸡源新城疫疫苗在控制鸽新城疫暴发流行中发挥了重要作用。本研究中,基于F基因的核苷酸同源性分析结果显示,6 株新疆鸽NDV流行株与基因Ⅱ型的疫苗株核苷酸同源性为83.5%~84.7%,与基因Ⅲ型Mukteswarz株同源性为84.9%~85.7%。结果表明,基因型不匹配的新城疫疫苗对鸽子的临床保护力有限,不能解决新城疫病毒脱落的缺点;在这种情况下,田间病毒仍可在鸽群中传播并引起鸽新城疫呈点状散发的流行特点。有研究报道,并非所有鸽子来源的病毒对鸡的毒力均很低,鸽子病毒在家禽中的持续传播可导致产生对家禽具有毒性的毒株,分离自孟加拉国的鸽子来源的NDV基因型ⅩⅪ.1.2对鸡的致死率很高[15]。因此,规模化养鸽场需要定期进行血清学与病原学监测并结合监测结果制定相应的免疫程序,同时还需加强养鸽场饲养管理及消毒管理,以达到有效防控鸽新城疫的目标。
4 结论
本研究结果显示,和田地区鸽NDV 基因型与我国鸽群中主要流行的NDV基因型Ⅵ的特征相符,6株F蛋白主要功能区均发生了点突变,且裂解位点序列具备强毒特征,但这些突变是否会影响病毒的致病性还需进一步研究加以证实。综上所述,加强鸽新城疫流行病学调查,掌握鸽新城疫病毒分子特征,对养鸽业及养鸡业的健康发展具有积极意义。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
民族古籍研究(2018年1期)2018-05-21
湖南畜牧兽医(2016年1期)2016-06-05
兽医导刊(2015年7期)2016-01-04
癌变·畸变·突变(2014年2期)2014-03-01
当代畜禽养殖业(2014年7期)2014-02-27
当代畜禽养殖业(2014年6期)2014-02-27
双语教育研究(2014年3期)2014-02-27
