
宰后贮藏期间氧化磷酸化对秦川牛肉色泽的影响
2023-12-05 08:54:06张萌李亚蕾
中国食品学报 2023年10期
张萌,李亚蕾
(宁夏大学食品与葡萄酒学院 银川 750021)
肉色及其稳定性是影响消费者购买肉品的重要因素之一[1]。宰后生鲜肉肉色的变化机制以及护色工艺一直是肉类科学的研究热点。当动物宰后充分放血时,肉品色泽主要由肌红蛋白含量和所呈现的化学状态决定[2]。随着机体宰后供氧的切断,肉品色泽受到肌肉耗氧量的影响。氧合肌红蛋白(Oxymyoglobin,OxyMb)随着机体耗氧量的增加转化成脱氧肌红蛋白(Deoxymyoglobin,DeoMb),而脱氧肌红蛋白更易被氧化成导致肉色褐变的高铁肌红蛋白(Metmyoglobin,MetMb),高铁肌红蛋白的积累是造成肉色劣变的主要原因[3]。蛋白质水平的变化会对肉色产生影响。传统的肉色研究方法不能反映内部蛋白质的变化,而采用蛋白质组学可以从蛋白变化角度探究肉色形成机制和外界因素影响肉色的途径。
蛋白质组学通过大规模筛选技术,对组织或细胞中所有蛋白进行定量、定性,确定其结构以及生物化学和细胞功能,可从整体上揭示蛋白质功能及蛋白质间的相互作用。目前,蛋白质组学已被应用于肉品科学领域,在肉品品质研究方面也取得许多进展[4]。罗辉等[5]通过蛋白质组学发现随着牛肉贮藏时间的延长,不同差异表达蛋白质对能量代谢存在不同程度的调控作用,且能量物质的变化对肉品质产生影响。Hughes 等[6-7]采用蛋白质组学研究不同牛种、不同肌肉部位的肌原纤维,结果表明结构蛋白丰度的变化影响光线的吸收和散射,从而改变肉色。Joseph 等[8]采用蛋白质组学分析肉色稳定性不同的牛背最长肌和腰大肌中的肌浆蛋白组差异,结果发现肉色稳定的背最长肌中氧化还原酶和伴侣蛋白的含量比肉色不稳定的腰大肌高。现有研究多集中于不同贮藏期影响肉色稳定性的因素,宰后贮藏期间肉色褐变相关生物标志物鉴定等,然而,贮藏期间能量关键代谢通路——氧化磷酸化在肉色形成过程中的代谢作用尚未见报道。本研究以25 月龄秦川牛肉背最长肌为研究对象,测定4 ℃贮藏0,4,8 d 时其pH 值、肉色和肌红蛋白相对含量的变化。基于4D-非标记蛋白质组学技术研究不同贮藏过程中氧化磷酸化代谢通路关键蛋白质表达量的变化情况,确定其在肉色形成过程中的作用,为宰后牛肉品质控制技术以及延长肉品货架期提供理论参考。
1 材料与方法
1.1 材料与试剂
牛肉样品,采集于宁夏尚农生物科技有限公司。
磷酸三钠、尿素、磷酸(85%)、氢氧化钠均为分析纯级,国药集团化学试剂有限公司;无水氯化镁、抗坏血酸钠、盐酸胍、考马斯亮蓝R250、2-硝基苯甲酸(DTNB)、乙二醇双(2-氨基乙基)醚(EGTA)均为分析纯级,上海麦克林生化科技有限公司;2,4-二硝基苯肼(2,4-DNPH)分析纯级,北京索莱宝科技有限公司;乙醇(95%)、氯化钠、十二烷基硫酸钠、氯化钾、乙二胺四乙酸(EDTA)均为分析纯级,西陇化工股份有限公司。
1.2 仪器与设备
SJ-3F 便携式pH 计,上海德图仪器国际贸易有限公司;CR-400 便携式色差仪,日本柯尼卡美能达公司;UV-1200 紫外分光光度计,上海美谱达仪器有限公司;TG16W 型离心机,湖南湘仪实验室仪器开发有限公司;LC-11L 拍打式无菌均质器,上海净信事业发展有限公司;JXFSTPRP-CL全自动样品冷冻研磨仪,上海净信事业发展有限公司;ImageScanner 扫描仪,美国GE Healthcare公司。
1.3 方法
1.3.1 样品采集 选取3 头体质量相近的25 月龄秦川公牛,屠宰后用灭菌刀取下右侧背最长肌,每个背最长肌样品平均分成3 份,共计9 个样本。编号后置于聚乙稀薄膜内,储存于风速3 m/s,相对湿度80%,4 ℃条件下。采集贮藏0,4,8 d 的样品约200 g 用于测定pH 值、肉色及肌红蛋白相对含量,采集不同贮藏时间的样品各200 mg,迅速置于液氮中放置2 h 后转移至-80 ℃冰箱中保存,以备蛋白质组学分析。
1.3.2 蛋白质组学分析 采用张杏亚等[9]的方法,从牛肉中提取总蛋白,取上清液测定蛋白含量,绘制标准曲线。4D-LFQ 蛋白质组学数据分析委托杭州景杰生物科技有限公司,运用MaxQuant(杭州景杰生物科技有限公司)软件系统获得蛋白质组学数据。基因本体注释(Gene ontology,GO)采用InterProScan(v.5.14-53.0)软件系统获得。京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通过KEGG 网站通路层级分类方法将这些通路进行分类。InterPro(对蛋白序列的家族分类、结构域和特殊位点的预测等功能分析提供资源)数据库用于分析差异表达蛋白的功能结构域的富集情况。
1.3.3 指标的测定
1.3.3.1 pH 测定 分别在贮藏期第0,4,8 天用便携式pH 计直接测量每块牛排的pH 值,共测定3 次,结果取平均值。
1.3.3.2 肉色的测定 冷却秦川牛肉贮藏第0,4,8 天,用便携式色差仪测定L*(亮度值)、a*(红度值)与b*(黄度值),仪器在测量前用白盘进行校准,提供3 个生物副本2.5 cm×2.5 cm×2.5 cm,每个副本在肉表面的6 个位置测量,取平均值。
1.3.3.3 肌红蛋白相对含量的测定 参考Tang等[10]的方法并适当修改。首先,制备磷酸盐缓冲液(0.04 mol/L,pH 6.8)和待测肉样(5 g),样品加入25 mL 缓冲液中,混合均匀;其次,在室内使用拍打式无菌均质器(12 次/s)均质10 min,所得溶液在冰箱中储存1 h,离心15 min(4 ℃,6 000×g),过滤上清液;最后,在503,525,557 nm 和582 nm 波长处读取滤液的吸光度,磷酸盐缓冲液作空白。
3 种肌红蛋白含量的测定均采用紫外分光光度法。在582 nm 波长处测定脱氧肌红蛋白的吸光值;分别在557,503 nm 波长处测定氧合肌红蛋白、高铁肌红蛋白的吸光值;在525 nm 波长处测定肌红蛋白总量的吸光值。3 种肌红蛋白相对含量计算公式:
式中,R1=A582/A525,R2=A557/A525,R3=A503/A525。
1.4 数据处理
所有试验指标均重复测定3 次,结果以“平均值±标准差”表示。采用Excel 软件统计数据,采用SPSS 软件对数据进行方差分析及相关性分析,采用Origin 软件作图。
2 结果与分析
2.1 肌肉的pH 值、肉色及肌红蛋白相对含量
由表1 可知,宰后0~8 d,秦川牛背最长肌pH值呈先下降后上升的趋势(P〈0.05),在第4 天达到最低点5.43。宰后成熟0~8 d 内牛肉的pH 呈先下降后上升的趋势,这一变化趋势与刘佳东等[11]对黄牛肉的研究结果一致。国内外诸多学者研究表明家畜屠宰后初期整体下降幅度大于上升幅度,可能是由于宰后机体氧气供应切断,机体通过无氧糖酵解产能的同时产生乳酸;此外宰后ATP水解产生磷酸,这些酸性物质长期积累无法分解与转运,使得肌肉组织的pH 值迅速下降[12]。而pH值在4~8 d 升高可能是由于分解代谢消耗了肌肉中所积累的酸性物质[13]。

表1 牛肉宰后贮藏过程中肌肉pH、肉色的变化Table 1 The changes of muscle pH and meat color during postmortem storage of beef
肉色是评价肉品质最直观的指标之一,决定肉品的经济价值,其主要由肌红蛋白3 种衍生态的相对含量和理化状态决定[14]。贮藏过程中秦川牛背最长肌色泽变化如表1 所示,L*值表示牛肉的亮度值,0~8 d L*值呈先升高后降低的趋势(P〈0.05),初期的上升可能是由于贮藏背最长肌汁液渗出较多,表面水分增加光的反射率导致L*值升高[15];而4~8 d 肉色的下降是由于肌红蛋白被不可逆地转化成高铁肌红蛋白致肉色变暗[16]。本研究结果与卢骁[17]的一致。a*表示牛肉的红度值,是评价肉色变化的重要指标。贮藏过程中a*值呈下降趋势(P〈0.05),这可能是由于长时间贮藏使牛肉的pH 值升高,细胞环境的改变,也使氧合肌红蛋白难以形成;且ATP 的降低导致还原反应无法发生,使脱氧肌红蛋白被不可逆地氧化成暗褐色的高铁肌红蛋白,因此红度值下降。本研究结果与Wu 等[18]一致。b*值表示牛肉的黄度值,如表1所示,随着贮藏时间的延长,牛肉的b*值呈上升趋势(P〈0.05)。这可能是由于长时间贮藏使肌肉与脂肪充分接触,肌肉中的脂肪被不断氧化,从而使b*值增加。本研究结果与刘佳东等[11]的一致。
本研究中贮藏8 d 内肌红蛋白相对含量的变化如图1 所示。牛肉中氧合肌红蛋白显著下降(P〈0.05),高铁肌红蛋白相对含量显著上升(P〈0.05),脱氧肌红蛋白变化不显著。活体内,脱氧肌红蛋白与氧合肌红蛋白、高铁肌红蛋白可相互转化,脱氧肌红蛋白在有氧条件下转化为氧合肌红蛋白,氧合肌红蛋白在有氧条件下转化为高铁肌红蛋白,而高铁肌红蛋白还原为氧合肌红蛋白和脱氧肌红蛋白是需要一定条件的,比如能量的维系以及细胞环境的稳定。本研究中高铁肌红蛋白相对含量显著上升,这可能是由于宰后成熟时间延长,pH降低导致细胞环境发生变化,同时肌肉中氧含量降低,ATP 大量的消耗无法维持还原反应的发生。罗辉等[19]研究表明在0~8 d 贮藏过程中ATP 不可逆地降解。高铁肌红蛋白不能被还原,导致其含量持续上升。这些因素综合影响肌红蛋白氧化-还原反应过程,最终导致肉色变差。
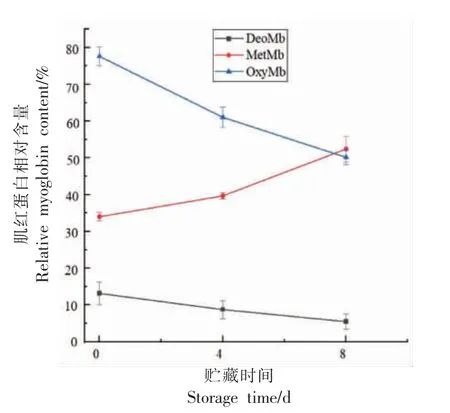
图1 冷鲜牛肉贮藏过程中3 种形式肌红蛋白的相对含量Fig.1 Relative contents of three types of myoglobin in chilled beef during storage
2.2 蛋白质组学数据分析
对不同贮藏期秦川牛背最长肌蛋白质表达模式进行样品相关性分析,以验证蛋白质组学样本采集及试验设置的合理性,结果如图2 所示。贮藏时间可将样本很好地区分开。同一贮藏时间样本间差异不明显,不同贮藏时间样本间差异明显。这一结果充分表明本试验样品采集合理,蛋白质组学数据可靠,可满足后续分析要求。

图2 不同贮藏期样本蛋白质组学结果的组间相关性分析Fig.2 Intergroup correlation analysis of proteomic results of samples with different storage periods
2.2.1 差异蛋白质筛选 以变化倍数(Fold Change,FC)〉1.5 或〈1/1.5,且P〈0.05 为标准筛选差异蛋白质,结果发现4 d 对比0 d 组共鉴定出15 个显著差异蛋白质,其中6 个为显著上调表达的差异蛋白质,9 个为显著下调表达的差异蛋白质(图3a)。8 d 对比4 d 组共鉴定出15 个显著差异蛋白质,其中6 个为显著上调表达的差异蛋白质,9 个为显著下调表达的差异蛋白质(图3b)。

图3 不同贮藏期牛肉差异蛋白质的火山图Fig.3 Volcano gram of differential proteins in beef at different storage periods
2.2.2 差异蛋白质生物信息学分析
2.2.2.1 Gene Ontology(GO)功能注释富集分析通过GO 注释对所筛选的差异蛋白质进行生物信息分析,从生物过程(Biological Process,BP)、分子功能(Molecular Function,MF)、细胞组分(Cellular Components,CC)3 个方面解析其功能,结果表明:4 d 对比0 d 组显著富集到137 个BP,63 个CC,35 个MF;8 d 对 比4 d 组显著 富集到83 个BP,58 个CC,49 个MF。
由图4a 可知,宰后贮藏0~4 d,差异蛋白质主要涉及的生物过程包括Rho-依赖蛋白负调控,高尔基间池囊泡介导的转运,Rho-依赖蛋白丝氨酸/苏氨酸的调控,微管正端调节,ATP 生物合成过程,蛋白丝氨酸/苏氨酸激酶的调节等。细胞组成主要包括交界肌浆网膜,线粒体质子转运ATP 合酶复合物,高尔基膜的外部成分,大分子复合物,细胞内部成分等。分子功能主要包括蛋白质结合,钙黏蛋白结合,细胞黏附分子结合,硫氧还蛋白过氧化物酶活性,果糖二磷酸醛缩酶活性,寡糖转移酶活性等。以上结果表明,牛肉屠宰后初期构成肌肉组织的交界肌浆网膜、连接膜复合体、膜结合囊泡等发生变化。这些变化可能引起细胞膜通透性发生改变,导致细胞内氧含量降低,细胞的主动运输发生障碍,细胞内能量无法正常交换,能量代谢发生紊乱,使褐色的高铁肌红蛋白还原反应无法进行,从而引起肉品色泽的变化。
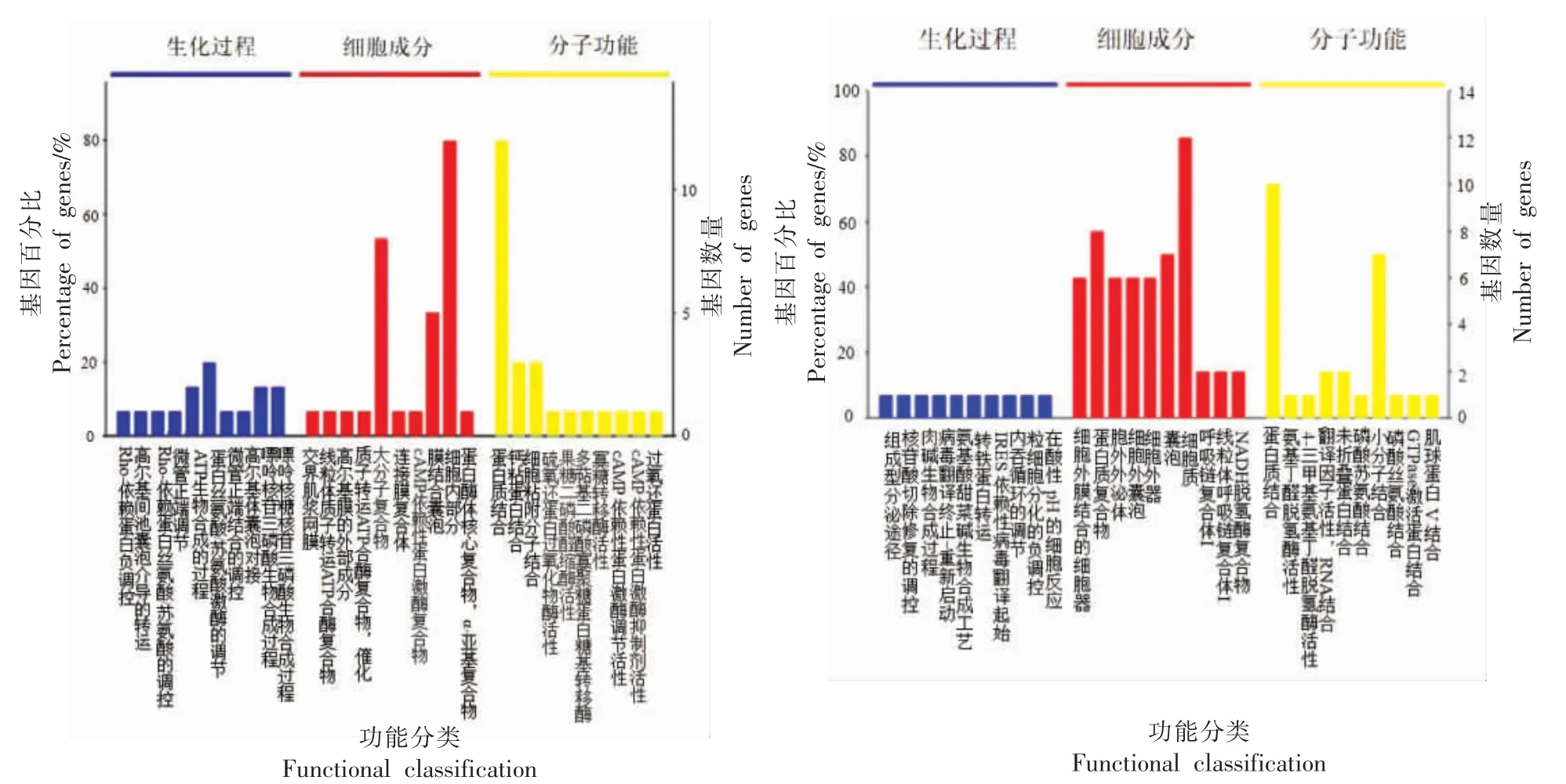
图4 不同贮藏期牛肉差异蛋白质的GO 注释(TOP 10)Fig.4 Gene ontology annotation of differential proteins of beef at different storage periods(TOP 10)
由图4b 可知,宰后贮藏4~8 d,差异蛋白质主要涉及的生物过程包括组成型分泌途径、核苷酸切除修复的调控、肉碱生物合成过程、转铁蛋白转运、内吞循环的调节等。在细胞组成中主要包括细胞外膜结合的细胞器、蛋白质复合物、胞外外泌体、细胞外囊泡、细胞外器等。在分子功能中主要包括蛋白质结合、氨基丁醛脱氢酶活性、4-三甲基氨基丁醛脱氢酶活性、未折叠蛋白结合、磷酸苏氨酸结合等。以上结果表明,牛肉贮藏4 d 后,除细胞膜通透性发生变化,差异表达蛋白质还涉及内吞循环调节和在酸性pH 的细胞反应等代谢过程。这些代谢过程的变化反映细胞内金属离子稳定性受到破坏和细胞环境的改变使细胞正常的能量代谢受到影响,其可能直接影响肌红蛋白及其衍生物的占比,使高铁肌红蛋白逐渐累积,肉色发生褐变。
2.2.2.2 京都基因与基因组百科全书(KEGG)途径分析 通过KEGG pathway 对所筛选的差异蛋白质进行通路注释分析,结果表明,4 d 对比0 d组显著富集到3 个KEGG 通路,8 d 对比4 d 组显著富集到17 个KEGG 通路。由图5a 可知,宰后贮藏0~4 d 差异蛋白质主要参与的KEGG 通路包括氧化磷酸化、N-聚糖的生物合成和胰岛素信号通路。由图5b 可知,宰后贮藏4~8 d 差异蛋白质主要参与的KEGG 通路包括抗坏血酸和醛糖酸盐代谢、丙酮酸代谢、氧化磷酸化、脂肪酸代谢、组氨酸代谢等。这些代谢过程可能通过影响宰后能量代谢方式,使高铁肌红蛋白还原反应的发生无法进行,间接影响肉品的色泽变化。同时,氧化磷酸化为0~4 d 和4~8 d 共同富集到的显著差异代谢通路,并且它对能量代谢有显著影响,而能量代谢在宰后成熟过程中对肉品质变化具有重要的调控作用。本研究以氧化磷酸化代谢通路为重点,探究氧化磷酸化代谢通路中关键蛋白在肉色形成中的重要作用。

图5 不同贮藏期牛肉差异蛋白质的KEGG 通路注释Fig.5 KEGG pathway annotation of differential proteins in beef at different storage periods
2.3 氧化磷酸化代谢通路关键蛋白质的变化
为明确氧化磷酸化代谢通路对宰后肉色的作用,通过蛋白质组学分析鉴定氧化磷酸化通路中关键蛋白质的表达情况,结果显示:贮藏4 d 与0 d 对比,鉴定出1 个显著差异蛋白质富集于氧化磷酸化代谢通路,为上调表达差异蛋白质。8 d 与4 d 对比,鉴定出2 个显著差异蛋白质富集于氧化磷酸化代谢通路,均为下调表达差异蛋白质(表2)。

表2 不同贮藏期氧化磷酸化代谢通路中富集的差异表达蛋白质Table 2 Differentially expressed proteins enriched in oxidative phosphorylation signaling pathways at different storage stages
ATP5F1D 即ATP 合酶亚基δ,它是线粒体ATP 合成酶的一个亚基,在质子转运和ATP 合成的偶联过程中起着重要的作用[20]。ATP 合酶参与ATP 的合成与消耗,因此其直接影响机体细胞的供能和耗能,也反映细胞中能量需求情况和三羧酸循环的产能情况[21]。本试验中ATP5F1D 在0~4 d 处于上调表达,8 d 与4 d 比较其表达量变化不显著,表明直至宰后8 d,ATP5F1D 表达量仍处于较高水平,这可能反映宰后短期内组织/细胞的自我保护机制。组织/细胞通过提高ATP5F1D 表达量,满足其正常代谢所需能量水平,然而,呼吸作用及血液循环的停止导致细胞环境发生改变,因此,组织/细胞无法通过提高蛋白质的表达量促进三羧酸循环,以达到正常产能的目的。其上调可能引起宰后能量代谢的转换。同时,ATP5F1D 表达量升高可能引起细胞内离子稳态失衡。张同刚等[22]研究表明,动物屠宰后其机体细胞氧气供能被切断,细胞内氧含量逐渐降低,会引起肌细胞内离子稳态的失衡,导致线粒体损伤,从而使后续pH 持续下降以及ATP 持续降解,高铁肌红蛋白被不可逆地转换,肉色变差。
NDUFA4 是细胞色素c 氧化酶(COX,complex IV)的一个副亚基,它在NADH 和呼吸链间的电子传递中起着重要的生理作用,维持和促进细胞内生物氧化的进行,并最终推动ATP 合成以维持正常生命活动[23]。而细胞色素c 氧化酶具有质子泵的作用,可将H+由基质抽提到膜间隙,同时可通过血红素中铁原子的氧化还原变化,传递给还原的氧,形成水。Purslow 等[24]研究表明宰后大量的H+会引起金属离子超载,从而激活蛋白水解酶,由此引起肌肉结构被水解,肌肉小片化程度加剧,可能会导致光在肌肉结构中的反射与折射发生变化,从而影响肉色。本试验中NDUFA4 在4~8 d 呈下调表达的趋势,这可能从侧面说明细胞环境和所需能量条件发生改变,导致细胞增殖减弱,细胞凋亡进入后期阶段[25]。
NDUFB5 是线粒体膜呼吸链NADH 脱氢酶I的副亚基,参与线粒体内膜上的呼吸作用,对电子从NADH 传递给辅酶Q 有催化作用[26]。调控NADH 脱氢酶的活性,NADH 脱氢酶可促使稳定肉色的甲基血红蛋白、高铁肌红蛋白、细胞色素b5 和细胞色素c 含量降低[27]。4~8 d 其表达量下调,可能是因为血液循环终止后,细胞内氧含量降低,有氧产能过程发生改变,使能量供给方式转变为糖酵解的过程,不利于维持肉色稳定。不同贮藏期氧化磷酸化代谢通路关键蛋白质的变化表明,屠宰后初期牛肉肌细胞内可能因H+跨膜交换而引起离子稳态的失衡,激活水解肌肉结构的内源酶,引起肌肉纤维小片化,导致光在结构中的反射与折射发生变化[24]。同时,能量代谢方式的改变引起pH 值降低,氧化还原反应不能逆向发生,褐色的高铁肌红蛋白无法被还原,因此,随着贮藏时间的延长,肉色变差。
2.4 贮藏期间肌红蛋白表达量的变化
肌红蛋白(Mb)是存在于肌肉纤维肌浆中的一种蛋白质,决定宰后肉的颜色状态[28]。秦川牛肉宰后0~4 ℃贮藏过程中肌红蛋白表达量变化如表3 所示。贮藏0~4 d 时,肌红蛋白表达量为上调表达,可能是因为氧气含量下降,使肌红蛋白在细胞水平通过提高表达量适应周边环境[29]。4~8 d 贮藏中,肌红蛋白表达量为下调表达。有研究表明肌红蛋白表达水平受氧气水平和线粒体钙稳态调控,因此贮藏后期的下调可能是由于含氧量降低使肌红蛋白的初级结构发生改变;同时,随着贮藏时间的延长,氧含量降低也会使肌细胞内离子稳态失衡,造成线粒体功能受损,ATP 持续发生降解,最终导致肌红蛋白表达量下降[3,30]。

表3 秦川牛肉贮藏过程中肌红蛋白表达量变化Table 3 Changes in myoglobin expression in meat of Qinchuan cattle during postmortem storage
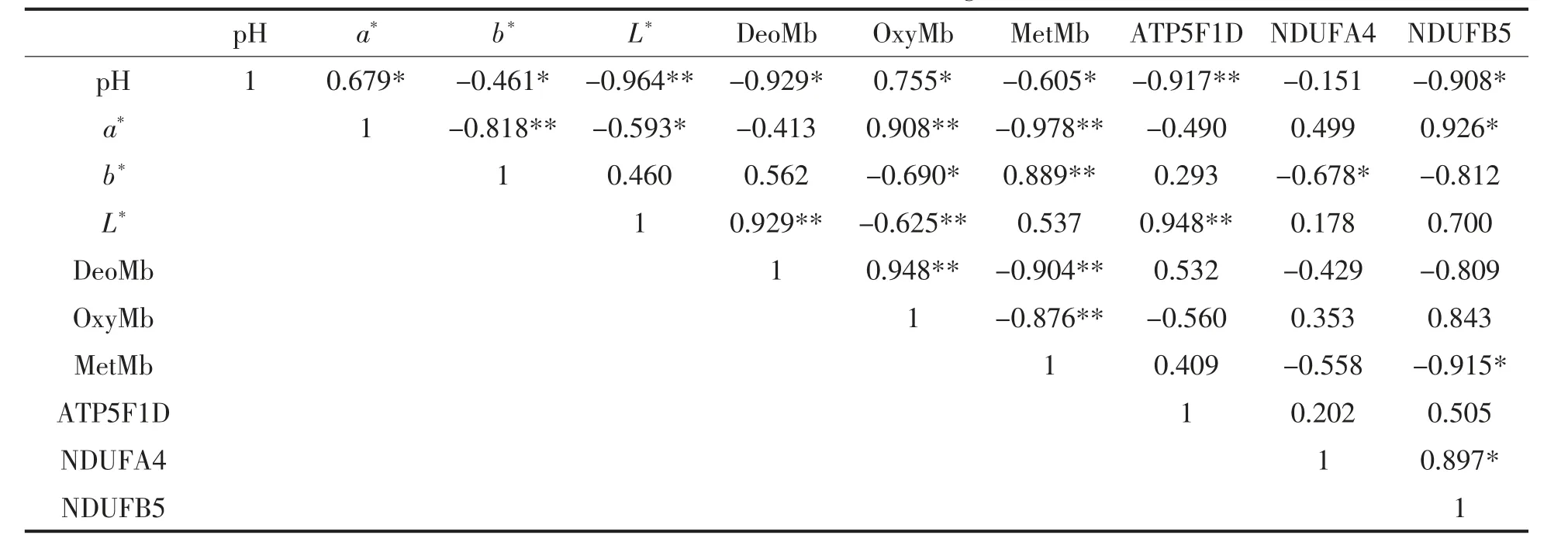
表4 不同贮藏时间牛肉pH、肉色、肌红蛋白相对含量与ATP5F1D、NDUFA4 及NDUFB5 的相关性分析Table 4 Correlation analysis of pH,meat color,myoglobin relative content and ATP5F1D,NDUFA4 and NDUFB5 in beef at different storage time
2.5 不同贮藏时间pH、肉色、肌红蛋白相对含量以及氧化磷酸化代谢通路关键蛋白的相关性分析
通过Pearson 相关性分析确定氧化磷酸化代谢通路关键蛋白质与秦川牛背最长肌色泽的关系。牛肉背最长肌pH 与肉色、肌红蛋白相对含量均呈显著相关性,与ATP5F1D、NDUFB5 呈负相关。a*值与NDUFB5 呈显著正相关(P〈0.05),b*值与NDUFB4 呈显著负相关(P 〈 0.05),L*值与DeoMb 相对含量呈极显著正相关(P 〈 0.01),与OxyMb 相对含量呈极显著负相关(P 〈 0.01),与ATP5F1D 呈极显著正相关(P〈0.01)。MetMb 相对含量与NDUFB5 呈显著负相关(P〈0.05)。相关性分析表明,氧化磷酸化代谢通路关键蛋白ATP5F1D、NDUFB5 和NDUFA4 通过引起宰后能量代谢方式转变对肉色产生显著影响。
相关性分析表明,氧化磷酸化代谢通路关键蛋白对pH、肉色以及肌红蛋白相对含量均有不同程度的影响。牛肉背最长肌pH 与肉色及其3 种肌红蛋白相对含量均呈显著相关,可能是因为宰后供氧被切断,细胞内环境发生改变,酸性物质长期积累而无法代谢。有研究表明pH 是影响宰后成熟过程中肉色形成的重要指标之一[2]。而ATP5F1D、NDUFB5 与pH 呈负相关,可能是因为能量代谢方式的转变,ATP5F1D、NDUFB 通过影响肌肉pH 值,间接影响肌红蛋白氧化还原状态。a*值与NDUFB5 呈显著正相关;b*值与NDUFB4呈显著负相关,可能是因为离子稳态失衡,激活了水解肌肉结构的内源酶,引起肌肉纤维小片化,导致光在结构中的反射与折射发生变化[31]。L*值与DeoMb 呈极显著正相关,与OxyMb 呈极显著负相关,表明宰后随着机体耗氧量的增加,细胞环境以及能量代谢方式的转变,MetMb 无法被还原,使得肉色变差,L*值也增大[32]。MetMb 与NDUFB5 呈显著负相关,这可能是因为能量代谢方式的改变引起pH 降低,氧化还原反应不能逆向发生,且随着贮藏时间的延长,肉色变差。综上,宰后成熟过程中肉色的变化由多个因素共同决定。Wu 等[33]研究表明氧化磷酸化在宰后贮藏过程中降低了MetMb的稳定性,导致深切牛肉色泽变差。杨啸吟等[34]采用蛋白质组学研究高氧气调包装下牛排肉色稳定性,结果表明宰后贮藏期间共15 个酶与肉色相关,且随贮藏时间的延长,表达量的下降,烟酰胺腺嘌呤二核苷酸和还原型烟酰胺腺嘌呤二核苷酸磷酸减少,从而使肉色稳定性发生变化[35]。氧化磷酸化代谢通路中的关键蛋白可能通过引起宰后能量代谢方式转变、加剧金属离子超载以及调控离子稳态平衡等方式最终导致3 种肌红蛋白占比发生改变,高铁肌红蛋白无法还原。
3 讨论
肌红蛋白是肉色的主要来源,肌红蛋白的氧化还原稳定性和鲜肉的颜色受多种因素控制,其表达量以及蛋白结构的改变对氧气运输和能量代谢有显著影响,从而引起肉色变化[36]。本研究中肌红蛋白表达量在贮藏0~4 d 呈上升趋势,4~8 d 呈下降趋势。肌细胞内含氧量的持续降低,一方面使肌红蛋白的初级结构发生改变,分子间相互作用降低;另一方面使肌细胞内离子稳态失衡,从而刺激下游一系列蛋白质表达的变化,引起不同时期ATP 的生物合成、嘌呤核苷三磷酸的生物合成等发生变化,细胞内能量代谢发生紊乱,最终导致肌红蛋白稳定性下降,表达量降低。氧化磷酸化关键蛋白ATP5F1D 在0~4 d 处于上调表达,在4~8 d处于较高水平。其表达量的持续上调可能引起宰后能量代谢的转换以及离子稳态的失衡,最终导致高铁肌红蛋白无法被还原,肌红蛋白表达量持续下降,肉色变差。NDUFA4 和NDUFB5 在4~8 d呈下调表达的趋势,说明细胞凋亡进入中后期。随着能量供给方式的转变,氧化-还原反应所需的细胞环境以及能量条件都发生改变,肌红蛋白表达量也持续下降,作为色素蛋白的肌红蛋白特有功能也在减弱乃至消亡,3 种肌红蛋白衍生态占比发生转变。综上,这3 种关键蛋白表达量的变化对肌红蛋白表达量均有不同程度的影响。
4 结论
宰后0~8 d 内,随着贮藏时间的增加,秦川牛肉背最长肌pH 值呈先下降后上升的趋势。L*值呈先上升后下降的趋势,a*值逐渐降低,b*值逐渐升高。氧合肌红蛋白显著下降,高铁肌红蛋白相对含量显著上升,脱氧肌红蛋白变化不显著。肌红蛋白表达量在0~4 d 上调,4~8 d 下调,总体为下降趋势。氧化磷酸化代谢通路上的关键蛋白质ATP5F1D 贮藏初期上调,NDUFA4、NDUFB5 贮藏后期下调,且氧化磷酸化代谢通路上的关键蛋白对牛肉背最长肌pH 值、肉色均有显著影响;L*值与DeoMb 相对含量呈极显著正相关,与OxyMb 相对含量呈极显著负相关,与ATP5F1D 呈极显著正相关。NDUFA5 与高铁肌红蛋白相对含量显著负相关。研究结果表明:氧化磷酸化代谢通路中的关键蛋白表达水平的变化对氧和细胞环境的影响使其能量代谢方式发生转变,从而引起细胞内离子稳态失衡,后续pH 持续下降以及ATP 的持续降解,使高铁肌红蛋白无法还原,肌红蛋白表达量持续下降。同时,氧化磷酸化关键蛋白质的变化可能引起贮藏期间金属离子超载,从而激活水解肌肉结构的内源酶。肌肉微观结构的变化也使光的反射、散射和吸收程度受到影响,使肉品色泽发生变化。综上,探究氧化磷酸化代谢通路关键蛋白质对肉色的影响,对于宰后肉品品质形成具有重要调控作用。
猜你喜欢
花卉(2023年21期)2023-11-16 07:54:00
食品科学(2022年17期)2022-09-30 07:49:12
现代临床医学(2021年2期)2021-03-29 05:32:00
解放军医学院学报(2020年12期)2020-03-29 05:11:02
食品工业科技(2018年20期)2018-10-24 01:09:28
幼儿教育·父母孩子版(2018年12期)2018-04-20 11:35:52
恋爱婚姻家庭(2017年18期)2017-12-26 07:20:42
黑龙江八一农垦大学学报(2017年1期)2017-03-10 01:46:42
广东饲料(2016年7期)2016-12-01 03:43:36
河南科技学院学报(自然科学版)(2014年3期)2014-04-29 09:08:54
