
两种旱生灌木种子萌发与幼苗复活对模拟干湿处理的响应
2023-11-22 09:38曲文杰赵文智王磊屈建军杨新国
草业学报 2023年11期
曲文杰,赵文智,王磊,屈建军,杨新国*
(1. 中国科学院西北生态环境资源研究院,甘肃 兰州 730000;2. 宁夏大学生态环境学院,宁夏 银川 750021;3. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021;4. 中国科学院大学,北京 100049)
植被更新涉及种子、幼苗、成熟植株整个生命周期,而种子萌发和幼苗初期阶段是其最脆弱时期,是植被更新中不可或缺的过程[1-2]。幼苗的低复活率,尤其当干旱加剧时种子萌发率和幼苗复活率更低,已被证明强烈影响种群动态、物种多样性、群落组成和生态系统功能等关键生态过程[3]。干旱地区降水量少且时空分布不均,并且降水很大程度上决定了干旱地区的生态过程[4-6]。因此,降水格局是荒漠植物种子萌发及幼苗早期成活的主要限制性因子之一。
已有研究表明,干旱区降水频次、强度和分布格局都可以显著影响种子萌发和幼苗生长过程[5,7-10]。干旱区降水少、蒸发量大、降水间隔期不一,植物经常面对不同周期的干湿变化和不同程度的干旱胁迫。种子经历干旱后再吸水的过程一般可以启动萌发机制[11],并影响到种子的寿命[12-13],种子萌发率[14-17],甚至包括种子萌发的速度[18]。植物可忍耐一定程度的脱水,复水后仍能恢复生长[15]。当固定根长的幼苗干旱达到某一时间时,复水后幼苗无法恢复,这个时间点被称为“不归点”[19]。一些耐旱植物的幼苗在生长早期就能耐受干燥,比如根长5 cm 的钝稃野大麦(Hordeum spontaneum)幼苗脱水21 d 后复水仍可恢复生长[20];根长为0.1 cm 的赖草(Leymus secalinus)幼苗的干旱不归点是60 d[15]。
柠条(Caragana korshinskii)、花棒(Corethrodendron scoparium)是腾格里沙漠东南缘防风固沙体系的典型多年生灌木,其根系发达,固沙能力强、耐风蚀、沙埋、干旱,是荒漠地区优良的防风固沙植物[21]。花棒是本地物种,柠条是建立防护体系时的引入物种。目前已有一些关于柠条、花棒种子对干旱胁迫、沙埋等响应的研究[22-26],但是关于它们萌发过程,以及干旱复水后早期幼苗的响应研究报道较少。因此,本研究的主要目标就是通过室内模拟的方法,分析不同湿润-干旱处理对柠条、花棒种子的萌发率,以及干旱复水处理对早期幼苗复活的影响,揭示柠条、花棒种子萌发和幼苗复活的环境适应策略,为研究区植被更新管理提供支持。
1 材料与方法
1.1 试验材料
2020 年7-11 月在腾格里沙漠东南缘的沙坡头沙漠试验研究站内(105°02′ E, 37°32′ N),采集成熟柠条和花棒种子,挑选饱满均一的种子进行模拟降水事件变化对种子萌发率和幼苗早期复活影响的试验。每个物种挑选100 粒种子通过种子分析仪(plant seed analyzer, OPTOMACHINES, OPTO-Agri, 荷兰)对其主要形态指标进行测定,结果见表1。

表1 柠条、花棒种子形态指标Table 1 Seed morphological indexes of C. korshinskii and C. scoparium (mean±SD)
1.2 湿润-干旱处理对种子萌发的影响
为探索不同湿润-干旱处理时间对柠条、花棒种子萌发的影响,设置了12 个湿润-干旱处理(即4h1d、4h2d、4h4d、4h8d、12h1d、12h2d、12h4d、12h8d、24h1d、24h2d、24h4d、24h8d)和对照处理(CK,未进行湿润-干旱处理,直接进行萌发试验),其中4h1d 表示湿润4 h 和干燥1 d,4h2d 表示湿润4 h 和干燥2 d,依此类推。两个物种共26 个处理,每个处理30 粒种子,重复5 次。将柠条、花棒种子用1%高锰酸钾溶液消毒15 min,用蒸馏水反复清洗5~7次后整齐摆放在直径9 cm 且垫有2 张whatman 1 号滤纸的一次性培养皿中,加入5 mL 蒸馏水,放入24 h 光照、25 ℃恒温、75%湿度条件的人工气候室进行湿润处理,然后转入室温实验室进行干旱处理。结束湿润-干旱处理的培养皿中加入3 mL 蒸馏水,置于光暗交替(12 h 光照)、25 ℃恒温、75%湿度条件的人工气候室进行种子萌发试验。每2 d 加入3 mL 蒸馏水保持湿润。萌发的判断依据为胚根出芽长度为1 mm。每24 h 统计一次并去除萌发的种子。连续5 d 不萌发视为试验结束。萌发率(%)=萌发种子数/供试种子数×100。
1.3 早期幼苗的干旱复水试验
为探索不同干旱时间对柠条、花棒早期幼苗的影响,设置了8 个干旱处理(即2、4、7、10、14、21、30、45 d),将萌发后的种子根据胚根长度分为4 个等级,即0~0.2 cm、0.2~0.5 cm、0.5~1.0 cm、1.0~1.5 cm。每个胚根等级都进行8 个干旱处理,两个物种共64 个处理,每个处理15 个早期幼苗,重复5 次。首先将柠条、花棒种子用1%高锰酸钾溶液消毒15 min,用蒸馏水反复清洗3~5 次后整齐摆放在直径9 cm 且垫有2 张whatman 1 号滤纸的一次性培养皿中,加入5 mL 蒸馏水,放入12 h 光照、25 ℃恒温、75%湿度条件的人工气候室对种子进行萌发获得早期幼苗。其次,将早期幼苗根据上述胚根长度进行分类后,整齐摆放在直径9 cm 且垫有2 张whatman 1 号滤纸的一次性培养皿中,放入室温实验室进行干旱处理。干旱处理结束后,加入5 mL 蒸馏水,放入12 h 光照、25 ℃恒温、75%湿度条件的人工气候室进行复活试验。每2 d 加入3 mL 蒸馏水保持湿润。如果早期幼苗的根进一步拉长,被认为有生命力,记录每天的复活数量。早期幼苗复活率(%)=复活幼苗个数/供试幼苗个数×100。
1.4 数据分析
采用R 4.2.0 软件完成试验数据分析。通过线性回归方法对比分析湿润-干旱处理对柠条和花棒种子萌发率的影响,以及对柠条和花棒种子萌发过程的影响,采用R 中sjPlot 包绘图。采用线性回归方法分析不同干旱时间、不同根长对干旱复水的早期幼苗复活率的影响,采用R 中visreg 包绘图。通过One-way ANOVA 在95%的置信水平上,用Tukey HSD 方法检验不同等级根长和不同干旱处理的早期幼苗复活天数之间的差异性,采用Origin 绘图。
2 结果与分析
2.1 湿润-干旱处理对柠条、花棒种子萌发的影响
如图1 所示,湿润-干旱处理显著提高了柠条种子的总体萌发率(8.31%,P<0.05),而对花棒种子没有显著影响(1.24%,P>0.05)。在全部处理中,只有12h1d 处理下两个物种的种子萌发率都出现了降低的趋势,但是依然不显著(P>0.05)。

图1 湿润-干旱处理对柠条和花棒种子萌发率的影响Fig.1 Effects of wet-dry treatment on seed germination of C. korshinskii and C. scoparium
2.2 湿润-干旱处理对柠条、花棒种子萌发过程的影响
如图2 所示,湿润-干旱处理显著提高了柠条和花棒种子的早期萌发率(P<0.05),但是两者的萌发过程和响应格局并不完全一致。湿润-干旱处理下柠条前3 d 萌发率依次为29.34%,19.40%和5.06%,相对CK 呈现出先提高后降低的响应格局。但是,4h1d、4h8d、12h4d 处理没有显著提高柠条种子的早期萌发率(第1 天,P>0.05)。与柠条不同,湿润-干旱处理显著提高了花棒种子第1 天(7.89%,P<0.05)和第2 天的萌发率(46.90%,P<0.05),却显著降低了第3 天的萌发率(22.40%,P<0.05)。第1 天,与CK 相比,仅有24h1d、24h2d、24h4d 显著提高了花棒种子的萌发率(P<0.05);第2 天,除4h4d、24h2d 和24h4d 外,其余处理都显著提高了萌发率(P<0.05);而第3 天所有处理都降低了花棒种子的萌发率,其中4h8d、12h2d、24h1d、24h2d、24h4d、24h8d 显著降低(P<0.05)。
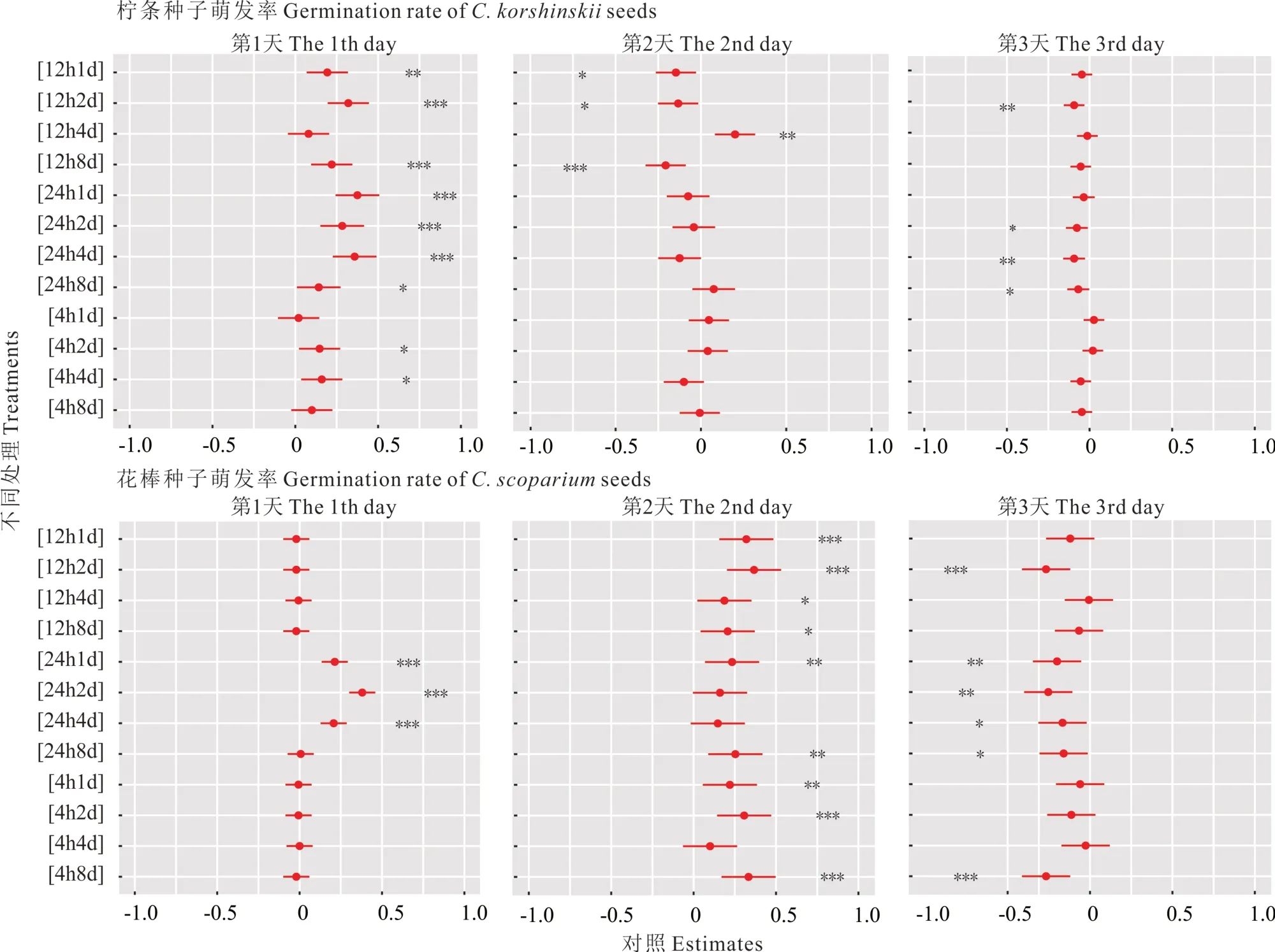
图2 湿润-干旱处理对柠条、花棒种子萌发过程的影响Fig.2 Effects of wet-dry treatments on seed germination process of C. korshinskii and C. scoparium
2.3 干旱复水处理对柠条和花棒幼苗早期复活率的影响
以根长为幼苗生长阶段(状态)的划分依据,研究发现随着干旱处理天数的增长和幼苗根长的增加,早期幼苗复水后的复活率也随之降低,但是花棒早期幼苗整体复活能力明显强于柠条(图3)。相比较其他等级的根长,根长为0~0.2 cm 的柠条和花棒早期幼苗复水后复活率最高,分别为77.20%和83.17%,而根长为1.0~1.5 cm 时复活率最低,分别为7.61%和21.17%。根长为0.5~1.0 cm 的柠条和花棒早期幼苗的干旱不归点分别是21 和45 d,而根长为1.0~1.5 cm 时干旱不归点分别是7 和14 d。花棒幼苗对干旱处理的总体响应格局与柠条相似,但是基本上花棒早期幼苗每个根长等级下复活率明显高于柠条,表明花棒早期幼苗具有很强的干旱耐受性。
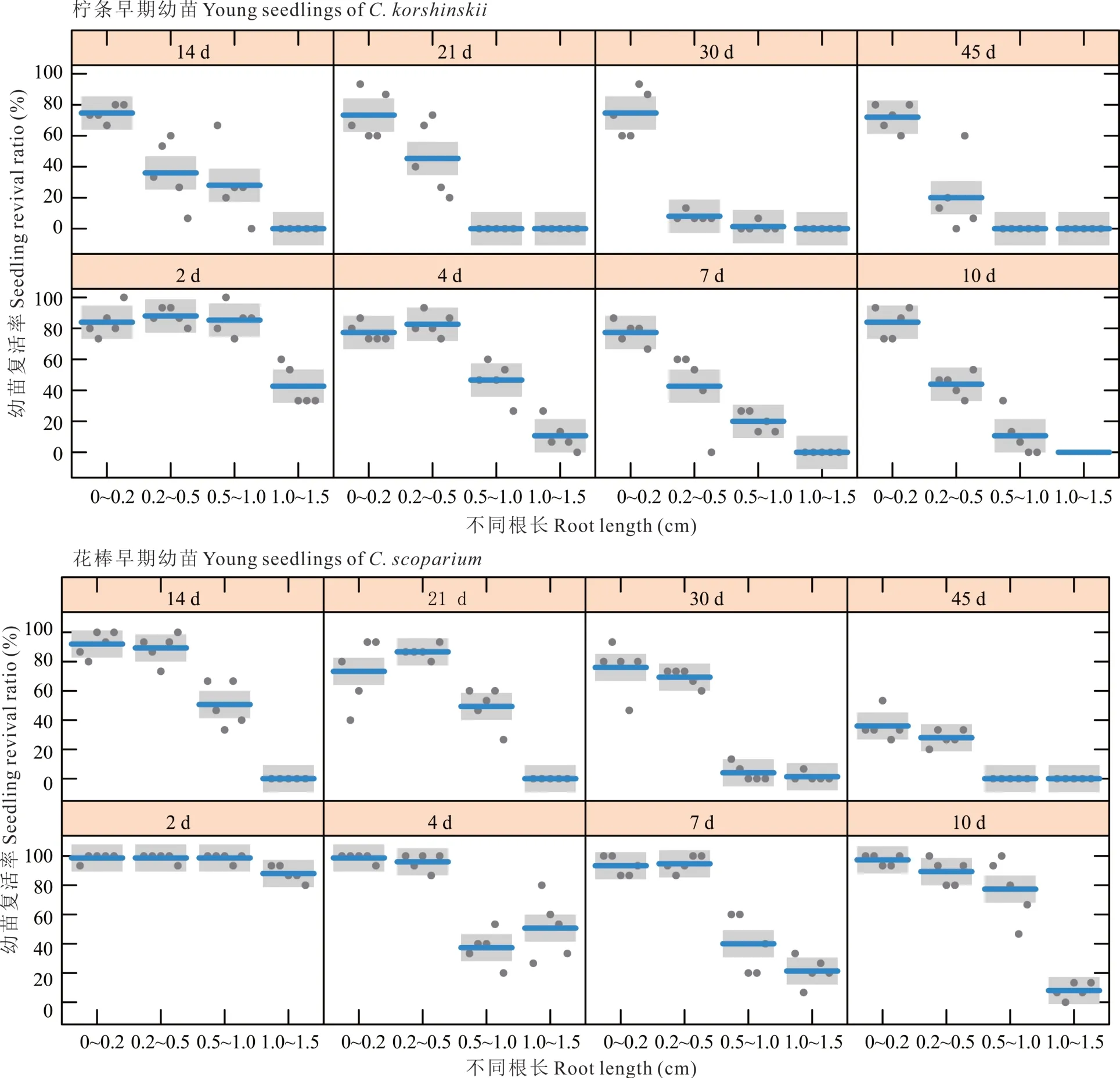
图3 不同干旱时间对早期幼苗(不同根长)复活率的影响Fig.3 Effects of different durations of drought on the survival rate of young seedlings with different root lengths
2.4 干旱复水处理对柠条和花棒幼苗早期复活时间的影响
由图4 可知,不同干旱处理早期幼苗复水后平均复活时间为7.38 d,达到最大复活率平均时间为3.48 d。干旱处理4 d 柠条早期幼苗复活所需天数最长,45 d 干旱处理下复活天数最短;干旱处理2 和45 d 时达到最大复活率所需天数较少。花棒早期幼苗经历不同干旱时间复水后所需复活天数和达到最大复活率的天数之间都没有显著差异(P>0.05),而柠条早期幼苗的显著差异主要表现在4 和2 d 以及4 和45 d 干旱处理之间,其他处理间没有显著差异(P>0.05)。根长为0~0.2 cm 时花棒早期幼苗完成复活天数最短[(5.84±0.46) d],根长1.0~1.5 cm时所需天数最长[(10.00±0.77) d]。比较而言,柠条早期幼苗复活所需天数[(6.39±0.37) d]短于花棒[(8.28±0.36) d],且达到最大复活率的天数[(3.05±0.22) d]短于花棒[(3.83±0.20) d]。

图4 不同等级根长和不同干旱处理幼苗复活天数Fig.4 Seedling revival days at different root length and different drying treatments
3 讨论
干旱地区生长季降水不可预测、且雨量不稳定,因此土壤表面种子经常遭受反复的湿润-干旱过程。植物通过对降水的适应可以形成不同的萌发策略[10,17-18,24]。本研究中,模拟湿润-干旱处理显著提高了柠条种子的萌发率,但是对花棒种子的萌发率无显著影响,表明花棒对环境变化相对不敏感,选择的是一种耐受型的萌发策略。但是,类似方法对加拿大的翼蓟(Cirsium vulgare, 二年生草本)处理后,发现对其萌发率有抑制作用[27],砂蓝刺头(Echinops gmelinii, 一年生草本)、赖草(L. secalinus, 多年生草本)也是抑制作用为主[15,28]。鉴于研究区植被构成中以柠条、花棒等多年生灌木和砂蓝刺头等一年生草本为主,灌草植被的种子对湿润-干旱的响应所存在的明显差异,反映了旱区高度水分竞争环境下群落内部物种间的反向适应对策选择,从而达成对潜在水分竞争的最大避免。
湿润-干旱处理显著提高了柠条种子的萌发率,表明湿润-干旱处理是柠条种子萌发的一种启动机制[11]。研究区的柠条一般在6 月底落种,当地7 月进入雨季,种子则集中在8-9 月萌发;表明经历多次湿润-干旱过程后,大多数种子的萌发机制已被启动,进入8 月伴随温度下降和浅层土壤水分含量的提高,柠条种子具备了集中萌发的充分条件[29]。但是,相对而言,花棒种子野外并未发生上述现象。这依然与两者对湿润-干旱的不同响应对策有关。
种子萌发后的幼苗生长阶段是限制植被更新的另外一个关键过程。干旱区植物种子萌发后缺乏降水或整株幼苗暴露在空气中,都可能导致幼苗脱水[15]。本研究发现,不管经历了2 d 还是45 d 的干旱,根长为0~0.2 cm 的柠条和花棒早期幼苗复水后,平均复活率分别达到77.20%和83.17%;但是随着根长的增长,幼苗干旱的不归点越来越早。这与以往的研究中幼苗脱水后的恢复取决于干燥时根系的长度和干燥时间长度的结果完全一致[20-21,30]。造成这种现象的机制是,胚乳作为种子贮藏营养部位,早期幼苗根系越长则种子中保留胚乳越少,意味着根越长复水后供幼苗再生的能量越少,复活率降低,不归点从而提前[15]。同时,比较可以发现,根长度相同时,花棒早期幼苗比柠条更耐旱,说明干旱胁迫下,花棒幼苗受到的伤害程度小于柠条,这与丁龙等[31]对柠条锦鸡儿、花棒实生苗对干旱胁迫的研究结果一致。总体而言,花棒一旦萌发后,其复活能力明显强于柠条。
花棒是腾格里沙漠东南缘地带性植被,至今在人工植被区和流沙区依然广泛地存在。而柠条主要是包兰铁路防护体系建设时的引入植被,仅存在于人工植被区。如上所述,柠条和花棒对干旱气候的适应对策存在显著的差异,这种差异也与研究区现有植被分布格局相吻合[29,32]。综合来看,伴随研究区人工柠条的逐步退化和自然更新的困难,在后续植被建设管理中,建议可以更多采用花棒一类的具有较高干旱忍耐力的灌木,逐步替代退化人工柠条,或辅助强化已有花棒灌丛的种子传播定居能力,从而提高现有人工固沙植被系统的多样性和稳定性。
4 结论
湿润-干旱过程有利于柠条和花棒种子萌发,但是随幼苗生长,对干旱的耐受程度也随之降低;相对而言,花棒幼苗对持续干旱的忍耐力要强于柠条。柠条和花棒是研究区最为常见的两种旱生灌木,其中柠条以人工引种为主,而天然花棒在流动沙地依然分布。在长期气候变化背景下,建议研究区人工固沙植被可以更多选择花棒一类的“忍耐型”灌木,进一步替代退化的人工柠条,从而提高固沙植被的多样性及其稳定性。
猜你喜欢
现代农村科技(2021年5期)2021-12-07
小小说月刊·下半月(2021年8期)2021-09-05
中国煤炭工业(2020年1期)2020-12-28
食品安全导刊(2020年33期)2020-12-26
当代陕西(2019年7期)2019-04-25
动漫界·幼教365(大班)(2018年3期)2018-05-14
山东农业科学(2017年4期)2017-05-18
湖南大学学报·自然科学版(2016年12期)2017-05-12
江苏农业科学(2016年4期)2016-06-14
现代农村科技(2016年6期)2016-03-28
