
短期养分添加对西藏沼泽化高寒草甸地上生物量、植物多样性和功能性状的影响
2023-11-22 09:38周娟娟刘云飞王敬龙魏巍
草业学报 2023年11期
周娟娟,刘云飞,王敬龙,魏巍*
(1. 省部共建青稞和牦牛种质资源与遗传改良国家重点实验室,西藏 拉萨 850000;2. 西藏自治区农牧科学院草业科学研究所,西藏 拉萨 850000)
长期以来,受人为干扰和气候变化等因素的驱动,青藏高原高寒草地正遭受着生物多样性降低、生产力下降和土壤养分流失等现象的困扰,其生态服务功能难以维持,草地生态系统正面临不同程度的退化[1]。外源性养分添加作为改善土壤肥力条件、增加草地生产力和恢复退化草地的重要途径被广泛应用[2]。
氮(N)、磷(P)是植物结构组成、生长发育和物质代谢的必需营养元素,N、P 养分添加对草地植物和土壤的影响研究不可枚举。研究普遍显示,N、P 是高寒草甸土壤主要限制养分[3-4],添加N、P 养分具有增加高寒草甸生产力、改善土壤理化性质、调节土壤微生物等优势作用[5],同时过量的N 添加也会引起土壤酸化,导致群落物种多样性的降低[6]。适量 P 添加可以通过提高土壤 P 有效性,调节土壤 P 供给,从而缓解 N 添加引起的植物群落多样性降低[7]。然而,由于不同植物群落对营养元素的利用与需求各异,有关N、P 添加的养分效应,不同研究者也给出不尽相同的结论,在功能群水平上,N 添加提高了高山嵩草(Kobresia pygmaea)高寒草甸禾草地上生物量,而N、P 同时添加则有利于莎草科(Cyperaceae)植物的生长[8]。群落水平上,牛玉斌等[7]研究温带荒漠草原在相同N(5 g·m-2)添加的基础上,当P 添加量增加到16.0 g·m-2时,显著降低了豆科植物的重要值和Pielou 均匀度指数、增加了Simpson 优势度指数。戚智彦等[9]关于典型草原研究结果为N、P 养分添加没有降低物种多样性。以往的研究均能够关注到养分添加对草地生产力和物种多样性的影响,但养分添加还可能同时改变群落中关键物种个体的功能性状,在养分添加的条件下群落水平上的功能性状变化比物种多样性变化更为敏感[10]。
群落水平的功能性状是个体水平植物功能性状与群落物种多度结合计算获得。近年来,基于群落水平功能性状对环境响应研究备受关注[11],并被证实群落水平的功能性状能更好地反映植物适应环境变化的策略。向响等[12]研究显示群落水平的功能性状受海拔决定下的热量和土壤养分的影响。张义等[13]对长期封育的典型草原研究表明,群落水平功能性状对草地群落地上生产力的贡献远高于物种多样性。因此,通过研究养分添加过程中草地地上生物量、物种多样性、群落水平的功能性状变化及其关系,可深入理解植物多样性、功能性状与生态系统功能的关系以及养分调控机制,为草地恢复和生态系统经营提供科学依据。
青藏高原沼泽化高寒草甸是高寒草甸类的亚类之一,净面积达1.85×106hm2,占高寒草甸的7.65%[14],其对青藏高原气候调节、水源涵养、生物多样性保护和碳汇等生态系统服务功能的维持发挥着重要作用[15],也是牦牛和藏羊主要的冬季草场。但由于长期放牧和刈割利用,面积出现萎缩,植株高度及盖度均有很大程度的降低,严重地限制了该草地生态系统的生产力维持。因此,通过外源养分提高草地生产力、研究养分添加对草甸植物的作用显得至关重要。本研究以当雄县羊八井镇甲多村长期围封刈割的藏北嵩草(Kobresia littledalei)沼泽化草甸为研究对象,开展短期N、P 肥料添加控制试验,分析不同养分添加条件下,西藏沼泽化高寒草甸群落生物量、植物多样性和群落水平功能性状的变化规律,明确植物多样性和群落水平功能性状对地上生物量变化的解释度,试图给出刈割沼泽化高寒草甸的养分补给策略,为西藏沼泽化高寒草甸的可持续利用管理提供依据。
1 材料与方法
1.1 研究区概况
试验地选在西藏拉萨市当雄县格达乡甲多村(29°59.31′ N、90°18.44′ E,平均海拔4460 m),该地属典型的高寒半干旱气候,冬季西风和印度夏季季风占主导。夏季多夜雨,冬季寒冷干燥,年平均气温-0.6 ℃,最冷月(1月)和最热月(7 月)气温分别为-9.0 和10.0 ℃。年降水量415 mm,70%集中在生长季(6-9 月),蒸发量约是降水量的4 倍[16]。
该区域植被分异明显,较高海拔(4600~5200 m)广泛分布着以高山嵩草为建群种的高寒草甸,植被覆盖度65%~80%,草层高度2~4 cm;低海拔(4300~4600 m)以大花嵩草(Kobresia macrantha)和丝颖针茅(Stipa capillacea)为建群种的高寒草原,植被覆盖度30%~40%;河漫滩区域生长着金露梅(Dosiphora fruticosa)灌丛,灌丛盖度30%~40%;水分富集区域发育着以藏北嵩草为建群种的沼泽化草甸,植被覆盖度75%~95%,草层高度15~30 cm。草原植物5 月初至5 月中旬返青,地上生物量通常在7 月下旬至8 月中旬达到高峰。土壤质地多为风化早期阶段的砂质和粉质土[17]。
1.2 试验设计
试验于2019 年4 月布设。为了减少自然地形和微环境条件的差异,挑选匀质性较高、物种组成一致,地下水位较低的藏北嵩草沼泽化高寒草甸进行围栏,围栏面积50 m×100 m,作为肥料短期添加的试验样地,传统的利用方式为刈割打草+冬季放牧。试验施肥量参考Niu 等[18]的标准设置,设置两种肥料添加,所施N 素为含46%纯N 的尿素,所施P 素为含46%P2O5的磷酸二铵。试验处理共计5 个,分别为:无养分添加(CK)、低氮(NL,N 素15 g·m-2·a-1)、高氮(NH,N 素30 g·m-2·a-1)、低氮+磷(NLP, N 素15 g·m-2·a-1+P 素20 g·m-2·a-1)和高氮+磷(NHP, N 素30 g·m-2·a-1+P 素20 g·m-2·a-1)。每个处理5 次重复,共计25 个小区。小区布设采用随机区组试验设计。小区4 个角用木桩固定,用铁丝围成面积2 m×2 m 的处理小区,小区之间相邻设置2 m 的缓冲隔离带。肥料添加时间为2019 年5 月中旬和7 月初,2020 年5 月中旬和7 月初,每次添加量占总量的1/2,添加时用等量的水溶解,均匀地喷施到小区内,试验地基础养分见表1。
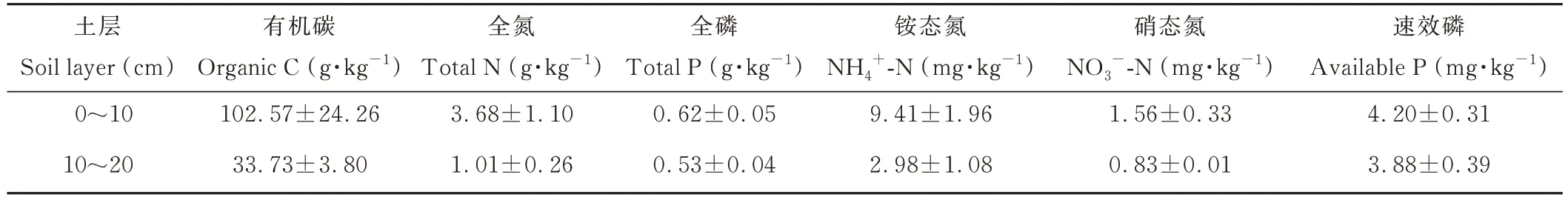
表1 试验地土壤养分概况Table 1 Soil nutrition of the test plots
1.3 野外取样和测定方法
地上生物量和植被群落调查选择在2019 和2020 年8 月中旬进行,此时地上生物量达到峰值。在不同处理样地内选择生长均匀的区域进行调查取样,样方大小为50 cm×50 cm,调查内容包括植物组成、物种数、植株高度、盖度、密度。随后齐地剪掉地上部分,按物种分装在牛皮纸袋中,带回实验室烘干至恒重(烘箱温度65 ℃)并称其干重,计算地上生物量。同时,将每种植物的叶与茎分开,测定叶片干重和地上总干重,计算单物种的叶比重。利用2020 年8 月实地测定的群落调查数据计算物种重要值和多样性指数。
2020 年8 月在每个试验小区内选择6 种[藏北嵩草、喜马拉雅嵩草(Kobresia royleana)、华扁穗草(Blysmus sinocompressus)、早熟禾(Poasp.)、鹅绒委陵菜(Potentilla anserine)、西藏报春(Primula tibetica)]常见的代表性物种(6 种植物占群落总盖度的90%以上),每个物种采集5~10 片完整叶片,分别装入密封袋中,标记好试验处理和重复数后迅速带回实验室进行植物性状的测定,在室内实验中,首先将每种植物的完整叶片进行扫描,并用ImageJ 软件获得叶片面积;之后将叶片浸水至饱和后擦干称质量获得饱和鲜质量,最后将叶片置于65 ℃烘箱烘干后称量,获得叶片干质量。将烘干后的叶片粉碎,用于叶片碳、氮、磷含量的测定。
植株高度(plant height, H)用卷尺测定每种植物的基部到冠层顶端的长度。叶比重(leaf mass per unit area,LA)是叶片干重与地上总干重的比值。比叶面积(specific leaf area, SLA)是叶片面积与叶片烘干质量的比值。叶干物质含量(leaf dry matter content, LDMC)是叶片的干质量与饱和鲜质量的比值。叶片碳含量(leaf carbon content, LCC)采用重铬酸钾容量法-外加热法,叶片氮含量(leaf nitrogen content, LNC)采用硫酸消煮-凯氏定氮仪(ATN-1100,上海)法,叶片磷含量(leaf phosphorus content, LPC)采用酸溶-钼锑抗比色法[19]。
1.4 数据分析
单个植物的相对高度(relative height,RHi)、相对密度(relative density,RDi)、相对盖度(relative coverage,RCi)、相对生物量(relative biomass,RBi)、相对重要值(relative importance value,Pi)计算公式如下:
本研究选取4 种常见的多样性指数分别为物种丰富度(S)、Shannon-Wiener 多样性指数(H)、Simpson 指数(D)和Pielou 均匀度指数(EP)。
式中:N为每一样方中的物种总数,Pi为相对重要值[20-21]。
植物群落水平的功能性状值(community-weighted mean trait values,CWM)的计算以相对密度(RDi)为权重[13],计算公式如下:
式中:Sc为每一施肥处理中测定性状物种数,RDi为物种i的相对密度,Traiti为物种i的性状值。
1.5 数据处理
数据处理和作图采用Origin 2018 软件,采用SPSS 16.0 中的One-way ANOVA 进行地上生物量、物种多样性及植物功能性状在不同养分添加的差异显著性分析(显著水平为P<0.05)。采用线性回归分析和 Pearson 相关系数分析地上生物量和植物群落水平功能性状、植物多样性指数之间的关系。采用R 语言Partial Least Squares Path modeling 模块分析氮、磷添加植物群落水平的功能性状、植物多样性指数对地上生物量的影响大小。方差分解利用“vegan”包的“varpart”功能来完成,数值采用“均值±标准差”表示。
2 结果与分析
2.1 沼泽化高寒草甸生产力变化
2019-2020 年,不同养分添加藏北嵩草沼泽化高寒草甸地上生物量变化如图1 所示。2019 年对照(CK)地上生物量为375.1 g·m-2,养分添加与CK 处理相比地上生物量均有不同程度的增加,NL、NH、NLP和NHP 处理增加量分别为106.4、87.7、157.7 和200.0 g·m-2,NLP 和NHP 处理与CK 相比差异显著(P<0.05)。通过分析2020 年的地上生物量变化情况得知,CK 处理地上生物量为416.4 g·m-2;经过2 年的养分添加,NL、NH、NLP 和NHP 处理的地上生物量较CK 分别增加163.5、323.0、411.7 和461.4 g·m-2。N添加和N+P 添加,地上生物量增加了39.3% 和98.9%。

图1 不同养分添加对沼泽化高寒草甸地上生物量的影响(2019-2020 年)Fig. 1 Effects of different nutrient additions on aboveground biomass of swampy alpine meadow (2019-2020)
2.2 沼泽化高寒草甸主要植物及功能群地上生物量变化
3 种莎草科植物(藏北嵩草、喜马拉雅嵩草、华扁穗草)、禾草类功能群和杂类草功能群的生物量分析见表2。结果显示,不同养分添加处理显著提高建群植物藏北嵩草的地上生物量,不同植物地上生物量变化对N、P 添加的响应各异。群落中,生物量组成仍以莎草科植物为主。藏北嵩草地上生物量在群落总生物量中所占比例最大,为93.8%以上;藏北嵩草在群落总生物量中占比随N 添加量的增加而增加。喜马拉雅嵩草地上生物量为6.44~32.73 g·m-2;NLP 处理显著高于CK、NL和NH处理(P<0.05)。华扁穗草地上生物量为0.31~ 10.35 g·m-2,NLP处理华扁穗草地上生物量在群落总生物量中所占比例最大,为1.43%。禾草类群地上生物量在NLP 和NHP 处理下均增加。杂类草类群的变化与禾草类群变化一致,NLP 和NHP 处理显著高于NL处理、NH处理和CK(P<0.05),NLP 处理杂类草类群在群落总生物量中所占比例最高,为1.19%。综上,N+P 添加与单独N 添加相比,显著增加了喜马拉雅嵩草、华扁穗草、禾草类群和杂类草类群的地上生物量(P<0.05)。

表2 主要植物及功能群地上生物量随养分添加的变化Table 2 Changes of aboveground biomass of main plants and functional groups with the nutrient additions
2.3 群落植物多样性分析
藏北嵩草沼泽化高寒草甸群落样地内共包含11 种植物(表3),隶属6 科9 属,其中,莎草科种类最多,有5 种。通过对不同养分添加处理间植物的地上生物量、植株高度、密度和盖度进行综合分析,计算重要值(表3)。沼泽化高寒草甸主要的优势植物为藏北嵩草,且藏北嵩草重要值在66.24%以上,伴生种为喜马拉雅嵩草、华扁穗草等,重要值分别为5.99%~10.35%、4.48%~5.49%。

表3 不同养分添加对草本植物重要值的影响Table 3 Effects of different nutrient additions on herbaceous importance values
不同N、P 添加处理下,植物的重要值变化明显。与CK 相比,N 单独添加藏北嵩草在群落中的重要值增加,增加了4.97%。与CK 相比,N+P 添加,藏北嵩草的重要值降低,降低了7.11%以上;喜马拉雅嵩草的重要值增加,增加0.39%以上;禾草类群中早熟禾的重要值明显增加,增加了6.29%以上;杂类草类群中鹅绒委陵菜、西藏报春的重要值增加。
不同养分添加处理群落物种多样性存在差异(表4)。N 添加降低了群落的物种丰富度、Shannon-Wiener 指数、Simpson 指数和Pielou 均匀度指数,NL、NH处理与CK 相比物种丰富度、Simpson 指数和均匀度指数差异不显著(P>0.05);NL处理Shannon-Wiener 指数显著低于CK(P<0.05);N+P 添加,显著提高了物种丰富度、Shannon-Wiener 指数和Simpson 指数(P<0.05),对Pielou 均匀度指数的影响不显著(P>0.05)。

表4 不同养分添加群落植物多样性变化Table 4 The response of species diversity characteristics to different nutrient additions
2.4 植物群落水平功能性状的变化
在短期养分添加过程中,植物群落水平功能性状单因素方差分析结果(图2)表明,不同养分添加的比叶面积(SLA)没有显著性变化,其余功能性状均有显著差异。养分添加处理植株高度(H)显著增加(P<0.05)。NHP 处理的叶比重(LA)显著高于CK(P<0.05)。NLP、NHP 处理的叶干物质含量(LDMC)与CK 相比显著降低(P<0.05)。叶片碳含量(LNC)随着单独N 添加量的增加显著增加(P<0.05)。不同养分添加处理叶片氮含量(LNC)显著增加(P<0.05),其中NL和NH处理下的LNC 含量显著高于NLP 和NHP 处理。叶片磷含量(LPC)随着N 添加量的增加先升高后降低,NLP、NHP 处理的LPC 含量显著高于NL、NH和CK;单独N 添加显著增加了叶片N/P(P<0.05),其值大于14;N+P 添加叶片N/P 与CK 相比差异不显著(P>0.05)。
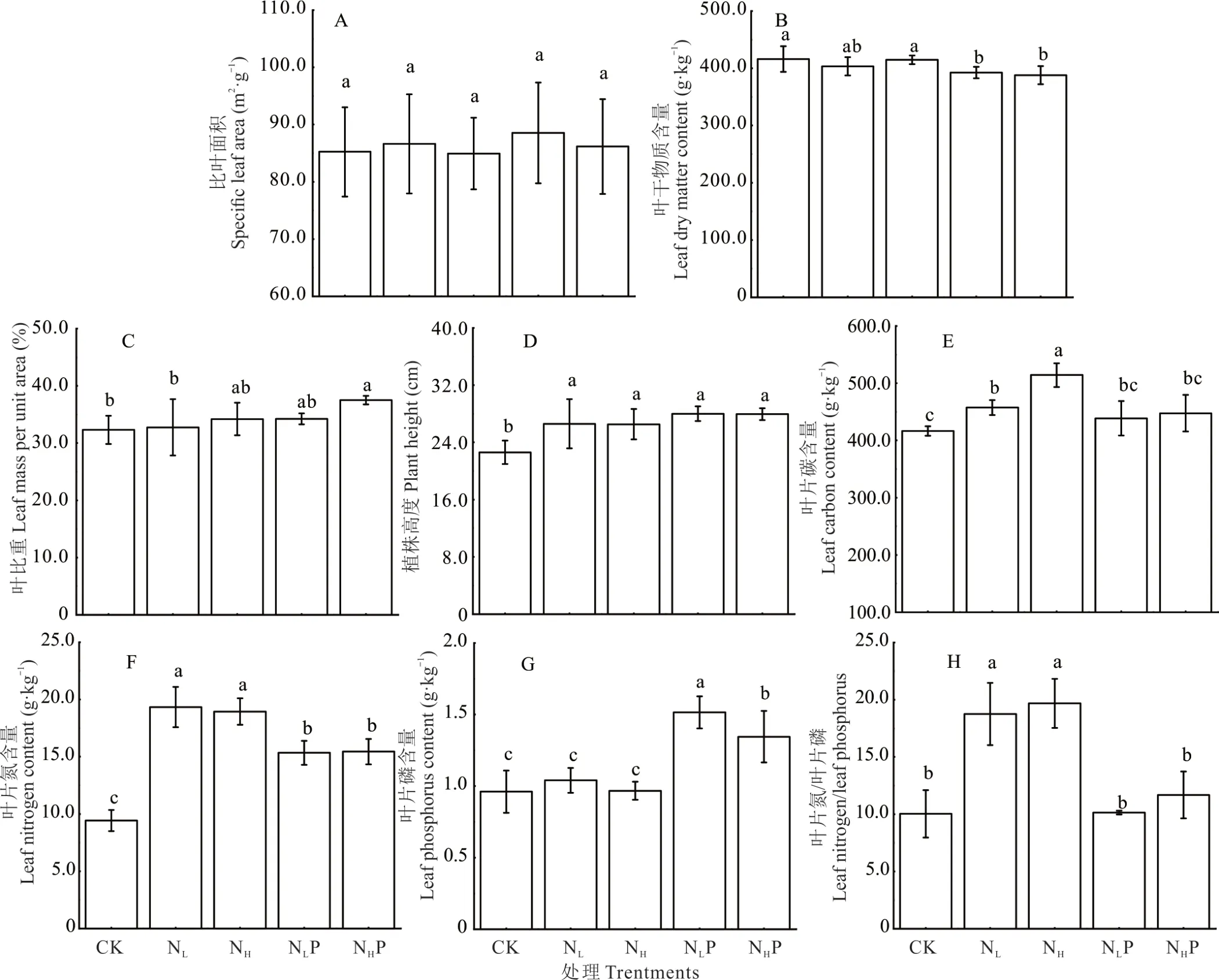
图2 不同养分添加植物群落水平功能性状的变化Fig.2 The response of community weighted mean functional traits to different nutrient additions
2.5 地上生物量与功能性状、物种多样性之间的关系
采用线性回归和 Pearson 相关系数分析地上生物量与植物群落水平功能性状的关系(图3)。结果显示,群落地上生物量与植物群落水平功能性状中植株高度(H)、叶干物质含量(LDMC)、叶片氮含量(LNC)、叶片磷含量(LPC)有相关性,与植株高度(H)、叶片磷含量(LPC)极显著正相关(P<0.01),线性回归拟合R2系数为0.5550和0.4389。与叶干物质含量(LDMC)显著负相关(P<0.05);与比叶面积(SLA)、叶比重(LA)、叶片碳含量(LNC)和叶片N/P 不存在线性回归关系(P>0.05)。
线性回归分析结果显示(图4),地上生物量与物种Pielou 均匀度指数相关性不显著,与物种丰富度、Shannon-Wiener 指数和Simpson 指数均有显著的正相关关系(P<0.05),其中物种丰富度、Shannon-Wiener 指数与地上生物量呈极显著正相关(P<0.01),线性回归拟合系数R2为0.3493 和0.2352。

图4 不同养分添加地上生物量与物种多样性指数的关系Fig.4 Regression relationships between aboveground biomass and species diversity under different nutrient additions
PLS-PM 偏最小二乘结构方程模型显示(图5),N 添加对植物群落水平功能性状和地上生物量有显著的正向促进作用(P<0.05),路径系数分别为:0.3012 和0.5081。P 添加对群落水平加权性状、地上生物量和植物多样性有显著的正向促进作用(P<0.05),路径系数分别为0.7595、0.7009 和0.8321。方差分解结果表明,植物群落水平功能性状和4 种植物多样性指数对养分添加下地上生物量的总解释度为81.3%,植物群落水平功能性状对地上生物量的解释度(78.5%)>植物多样性的解释度(45.5%)。
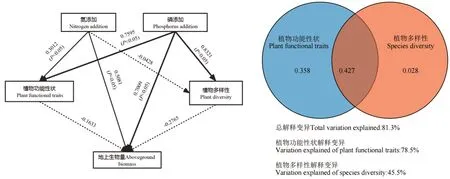
图5 养分添加时地上生物量决定因素的结构方程模型和方差分解Fig. 5 Structural equation model and variance decomposition for factors influencing of aboveground biomass under different nutrient additions
3 讨论
3.1 短期氮、磷添加对沼泽化高寒草甸地上生物量和植物多样性的影响
以往研究,由于人类的过度利用(放牧+刈割)沼泽化高寒草甸土壤养分和有机质含量持续下降,群落地上生产力呈降低的趋势[14]。通过对西藏沼泽化高寒草甸连续2 年的养分添加试验发现:群落水平上,地上生物量随着氮添加量的增加而增加。氮添加后,地上生物量较对照(无添加)至少增产39.3%;而氮、磷同时添加时,地上生物量较对照(无添加)至少增产98.9%,增加近1 倍。表明通过氮、磷添加可快速补充土壤养分,对地上生物量的促进作用非常明显,该沼泽化高寒草甸生产力受氮、磷养分的共同限制,这与Bowman 等[22]在落基山脉湿草甸的研究结果一致,氮是限制高寒草甸和高寒湿地草甸的主要养分[23],而氮、磷养分混合添加对草甸地上生物量影响更为显著。西藏沼泽化高寒草甸建群物种为莎草科植物,这也印证了宗宁等[24]的研究结果,莎草科植物生产力受氮、磷共同限制。在物种水平上,短期养分添加对地上生物量的贡献主要来自藏北嵩草生物量增加,藏北嵩草为短根茎型草本,以根茎芽为主的无性繁殖为主导,养分添加时,根茎芽大量萌发,分枝密度增加,进而导致地上生物量增加。多年生丛生禾草和杂类草主要以分蘖和分枝芽进行繁殖,属资源保守型植物[9],因此,单独氮添加对禾草类和杂类草的地上生物量影响不显著。值得注意的是,喜马拉雅嵩草地上生物量随着氮添加量增加而降低,可能的原因是不同莎草科植物对氮肥的利用策略和自身的繁殖策略不同。氮、磷同时添加,土壤氮、磷养分限制得到同时缓解,造成群落中的藏北嵩草、喜马拉雅嵩草、华扁穗草、禾草类和杂类草地上生物量均有不同程度增加。本研究中,养分添加第2 年相比第1 年各处理间地上生物量差异显著性增高,受土壤中的养分逐渐累积所影响。
本研究中,物种多样性结果显示,单独氮添加导致物种丰富度、Shannon-Wiener 指数和Simpson 指数均降低,究其原因是氮添加快速增加藏北嵩草根茎芽生长且密度增加,从而减少养分和空间,导致其他植物的重要值降低。宗宁等[8]研究表明,氮肥过量不利于草地物种多样性的维持,有可能导致土壤酸化而出现负面效应。单独氮添加不是草甸改良的优良做法。氮、磷同时添加,导致物种丰富度、Shannon-Wiener 指数和Simpson 指数均升高,这与戚智彦等[9]的研究结果类似。有研究表明,养分添加导致物种多样性丧失[25],丧失的原因是养分添加导致植物间受光竞争影响,由于土壤养分增加造成养分利用率较高的植物快速生长,对其他低矮植物造成遮阴,进而降低植物多样性。本研究中,沼泽化高寒草甸群落以藏北嵩草为优势种,自身郁闭度较大,光竞争不是植物多样性的限制因素,植物多样性变化主要受土壤最低养分含量的影响。在氮、磷同时添加时,物种多样性与对照相比显著增多,说明磷的添加能缓解不同植物对养分需求的限制。关于养分添加量,本研究给出的氮添加量30 g·m-2配施磷素20 g·m-2可有效提高地上生物量,增加物种多样性。马玉寿等[26]研究高山嵩草草甸短期氮肥适宜添加量为30 g·m-2·a-1,而在王长庭等[27]对藏嵩草草甸连续6 年施肥研究表明,氮添加量为9.3 g·m-2·a-1对群落地上生物量影响不显著,但显著影响了土壤微生物群落的变化。二者区别在于添加量的差异。养分添加量不仅要考虑地上生物量变化,还要系统衡量地下根系、土壤养分和微生物等综合的变化。养分添加量对草地植被群落稳定性的研究仍需要开展长时间的试验观察确定。
3.2 短期氮、磷添加下植物群落水平功能性状的变化
植物功能性状是植物定植、生长发育、死亡过程的特征属性,这些属性能表征植物生存策略和对环境变化的响应[28]。本研究中,植物群落水平上功能性状受植物功能性状与物种多度共同影响。养分添加条件下植物群落水平的植株高度显著增加,氮添加30 g·m-2·a-1、磷添加20 g·m-2·a-1处理时叶比重也显著增加,主要是该类草地受氮、磷养分限制,养分添加植物通过提高植株密度、增加高度、增加叶生物量来获得更多光资源,提高生存能力。具备较低的比叶面积和较高的叶干物质含量的植物具有生长速度慢的生存策略[29],本研究中,高海拔生长的莎草科和禾本科为了适应高原寒冷环境,叶面积均较小[30],不同养分添加处理对群落水平上的比叶面积变化影响不显著。氮、磷同时添加时降低了群落水平上的叶片干物质含量,与杂类草中鹅绒委陵菜、西藏报春叶干物质含量降低有关。植物叶片碳、氮、磷含量与植物对土壤养分的吸收利用能力和植物的贮存养分的状态密切相关[31],本研究中单独氮添加显著增加了群落水平的叶片碳含量,这与游郭虹等[31]在四川红原研究结果不一致,有机碳虽然是植物结构组成物质,在植物叶片中一般稳定存在,不随土壤养分变化而变化,造成这个结果可能的原因是群落植物碳分配格局发生了改变,破坏了植物自身资源的分配稳态[32]。群落水平的叶片氮、磷含量对养分添加响应明显,随着单独氮添加,群落水平叶片氮含量显著增加,有研究显示,氮添加显著增加群落中不同植物的叶片氮含量[33]。叶片磷含量随着单独氮添加呈现先增加后降低的趋势,原因是氮元素输入量的迅速增大加速了该草地生态系统的磷循环,进而增加了叶片的磷含量,随着植物吸收磷的增加更加剧了土壤中潜在的磷限制,当植物生物量的增加产量大于植物对磷的吸收,随后叶片磷养分产生稀释,磷含量随之降低[34]。氮、磷同时添加显著增加了群落水平的叶片氮、磷含量,李冬冬[35]研究显示,叶片有机氮和有机磷与土壤可利用性氮、磷呈正相关关系。本研究表明,有些群落水平上的功能性状对单独氮添加和氮、磷混合添加的响应较为敏感,结合功能性状和物种多样性能够更好解释群落生产力的变化。
3.3 植物多样性与群落水平功能性状对地上生物量变化的解释作用
在草地自然群落试验研究中对植物多样性与生产力的关系解释各异。Hector 等[36]研究指出,生态位分化和互补效应是植物多样性导致草地生产力增加的理论基础[36],陈丽等[37]在天山高寒草地的研究表明,多种养分添加比单独养分添加对物种多样性减少作用明显,植物群落地上生物量与Shannon-Wiener 指数、Simpson 指数和均匀度指数均呈负线性回归关系。Niu 等[18]在西藏高寒草地通过3 年的养分添加得出生产力与物种多样性不相关。本研究中,养分添加2 年后,草地群落地上生物量与草地物种丰富度、Shannon-Wiener 指数和Simpson 指数正相关,并呈线性回归关系。说明生态位分化和互补效应得到支持。最小二乘法结构方程模型和方差分解结果显示,N 添加对群落水平功能性状和地上生物量有显著的正向促进作用,P 添加对植物群落水平功能性状、地上生物量和植物多样性有显著的正向促进作用,氮、磷共同决定了地上生物量的增加。植物群落水平功能性状和4 种植物多样性指数对地上生物量的总解释度为81.3%,植物群落水平功能性状对地上生物量的解释度为78.5%,4 种植物多样性指数的解释度为45.5%。原因是氮添加对植物多样性影响不显著,而氮、磷同时添加显著增加了植物丰富度和多样性,地上生物量的增加与植物丰富度和多样性指数呈正相关关系。因此植物多样性对地上生物量变化存在一定解释能力。植物群落水平的功能性状不仅综合了不同植物的属性,还可更好地表征群落动态变化和植物利用资源的能力,其比植物多样性能更好地解释草地生产力变化[13]。本研究表明,群落水平的植株高度、叶片干物质含量、叶片氮含量、叶片磷含量均与地上生物量呈线性回归关系。功能性状均可以很好地解释地上生物量变化。说明在短期氮、磷添加群落中主要植物的功能性状驱动了生物量的增加。上述结果表明植物群落水平的功能性状比植物多样性能更好地解释地上生物量对短期养分添加的响应变化。
4 结论
综上可知,在藏北嵩草沼泽化高寒草甸短期养分添加的过程中,地上生物量随着养分添加量的增加而增加,氮磷混合添加优于单独氮添加,氮添加量30 g·m-2配施磷素20 g·m-2可有效提高地上生物量,增加物种多样性。地上生物量较无添加处理提高461.4 g·m-2,增产110.8%;喜马拉雅嵩草、早熟禾、鹅绒委陵菜、西藏报春的重要值都有不同程度增加。养分添加处理显著提高了群落水平的植株高度和叶片氮含量,氮+磷混合添加显著增加叶片群落水平磷含量,氮+磷混合添加增加了叶比重,降低了叶干物质含量。群落水平植物功能性状和4 种植物多样性指数对地上生物量的总解释度为81.3%,群落水平植物功能性状对地上生物量的解释度(78.5%)>植物多样性的解释度(45.5%)。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年19期)2020-12-14
活力(2019年21期)2019-04-01
中国科技信息(2015年2期)2015-11-16
水生生物学报(2015年1期)2015-02-28
植物营养与肥料学报(2014年1期)2014-03-11
