
荒漠草原人工柠条引入后土壤团聚体几何分布及拓扑结构演变特征
2023-11-22 09:38朱志昊孟晨王兴宋乃平王丽徐苗苗杜灵通
草业学报 2023年11期
朱志昊,孟晨*,王兴,宋乃平,王丽,徐苗苗,杜灵通
(1. 宁夏大学西北土地退化与生态系统恢复国家重点实验室培养基地,宁夏 银川 750021;2. 西北退化生态系统恢复与重建教育部重点实验室,宁夏银川 750021;3. 宁夏大学生态环境学院,宁夏 银川 750021;4. 宁夏大学农学院,宁夏 银川 750021)
土壤团聚体是土壤的重要组成部分,是土壤结构的基本单位[1-2],其结构、分布及稳定性对植被的恢复与生长有着重要影响,对区域生态恢复和防止表土沙化具有重要意义[3]。量化异质性土壤中团聚体结构是评价土壤和植被管理措施时必须考虑的一个重要因素[4],大量研究证明了团聚体结构对水土保持、土壤功能、植物生长的重要作用[5-10],尤其是团聚体拓扑几何结构及分布,其会对孔隙网络产生重要影响,从而影响水分、养分等资源的储存与运移[11-12]。然而,以往对土壤团聚体的研究局限于对水稳性团聚体数量及大小的量化分析[13-22],而对异质性原状土壤中团聚体几何分布情况及拓扑结构特征的认识不够,阻碍了对土壤团聚体形成过程、机制及其生态功能的充分认识。
团聚体的形成是一个自然过程,但其会受到土地利用、耕作干扰、施肥和种植制度等人类活动的影响[23],同时,土壤管理策略、土壤性质和植物根系性状复杂的相互作用共同决定着土壤团聚体的稳定性[13]。人工植被的引入及持续恢复,会导致团聚体稳定性发生变化[14],植物生长也可以改变土壤的力学以及水文特征,以促进土壤团聚体的形成和稳定[15,24],并最终增加水稳性团聚体的含量[25]。例如,>2 mm 粒级团聚体含量会随着杉木(Cunninghumia lanceolata)人工林林龄的增长呈先增后减趋势[16];>0.25 mm 粒径团聚体会随着玉米(Zea mays)种植年限的增加呈持续稳定增加趋势[17];而杉木人工林的中龄林和成熟林土壤团聚体结构优于幼龄林[18]。通过以往研究也可发现,不同植物种在引入、恢复中其土壤团聚体演变过程具有差异性[26-27],因此需要针对典型植物开展研究。柠条(Caragana korshinskii)生态恢复功能强,是我国西北荒漠地区的主要人工营造树种[28]。但是,在柠条生长至一定年限后土壤结构性质会产生再退化现象,影响表层土壤根系生物量,从而影响表层土壤结构发育及演变[29]。例如,随着柠条林龄的增长,土壤逐渐干化形成干层,且干层厚度及深度均呈增加趋势[30],降低了水土保持、生物多样性,影响了生态恢复的可持续性。因此,有必要关注柠条引入后表层土壤中团聚体结构的演变过程。本研究将利用计算机断层扫描(computed tomography,下文称CT 扫描)[31-34]结合图像处理分析,对荒漠草原不同林龄柠条林地土壤团聚体几何分布及拓扑结构特征进行量化分析,以期为明晰荒漠草原区柠条林在不同生长阶段的水土资源利用策略、维持柠条林生态效益提供重要的科学依据,并为今后土壤团聚体形态与功能等方面的深入研究提供技术支撑。
1 材料与方法
1.1 研究区概况
研究区位于宁夏回族自治区吴忠市盐池县内(37°57′23″-37°83′47″ N, 106°77′99″-107°50′70″ E),平均海拔1600 m,主要的土壤类型是灰钙土和风沙土。研究区域属于温带大陆性气候,年平均气温为8.1 ℃,无霜期为150 d。年均降水量为280 mm,80%以上的降水量集中在5-9 月,其中全年60%以上的降水又集中在7-9月,年均蒸发量为2710 mm。西风和西北风为全年主要风向,主要发生在春季。研究区植被多以牛枝子(Lespedeza potaninii)、老瓜头(Cynanchum komarovii)、草木樨状黄耆(Astragalus melilotoides)、猪毛蒿(Artemisia scoparia)、狗尾草(Setaria viridis)、短花针茅(Stipa breviflora)、苦豆子(Sophora alopecuroides)、绵蓬(Corispermum hyssopifolium)、阿尔泰狗娃花(Heteropappus altaicus)等植被为主。
1.2 样地选取与取样方法
试验于2021 年5 月开始,共设置5 个样地,根据柠条不同种植年份选取4 个试验样地,另选取未种植柠条的草本样地为对照组。柠条林地未引入柠条前均为放牧草地,样地基本情况见表1。

表1 样地基本信息Table 1 Plot fundamental information
在4 种柠条引入(9、14、24、35 年生)样地中,每种选取2 个样地重复,每个样地选取2 个样点重复,草地样地选取2 个样点重复,在每个样点的0~10 cm、10~20 cm 土层利用自制超薄钢材环刀(高65 mm,内径50 mm,厚度小于0.5 mm,为了保障根系构型分析精度,单个扫描样品高度控制在65 mm)取样后利用nanoVoxel-5000 系列双射线源CT 系统(三英精密有限公司)扫描分析根-土原状结构,即有(4 种林龄样地×2 个样地重复×2 个样点重复+1 个草地×2 个样点重复)×2 层土壤共计36 个环刀样用于CT 扫描。采样过程中,首先去除土壤表面的枯落物层,再将环刀底部平放在土壤,利用反铲小心地铲开环刀外围的土壤,并同时令环刀缓慢地垂直深入。为了避免土壤被破坏,在每根环刀的上下两端各固定一块铁板,为了防止土壤移动和渗漏,在铁板外用保鲜膜包裹,在固定铁板的同时,减少了土壤水分的蒸发。土柱采集完毕立即送往实验室进行CT 扫描,运输的过程中需要将土柱固定,避免运输途中颠簸造成的土体破坏,减少运输过程导致的误差。最后利用铝盒、自封袋、普通环刀取得土壤样品(各3 个重复)带回实验室进行测定。样地土壤基本性质见表2。
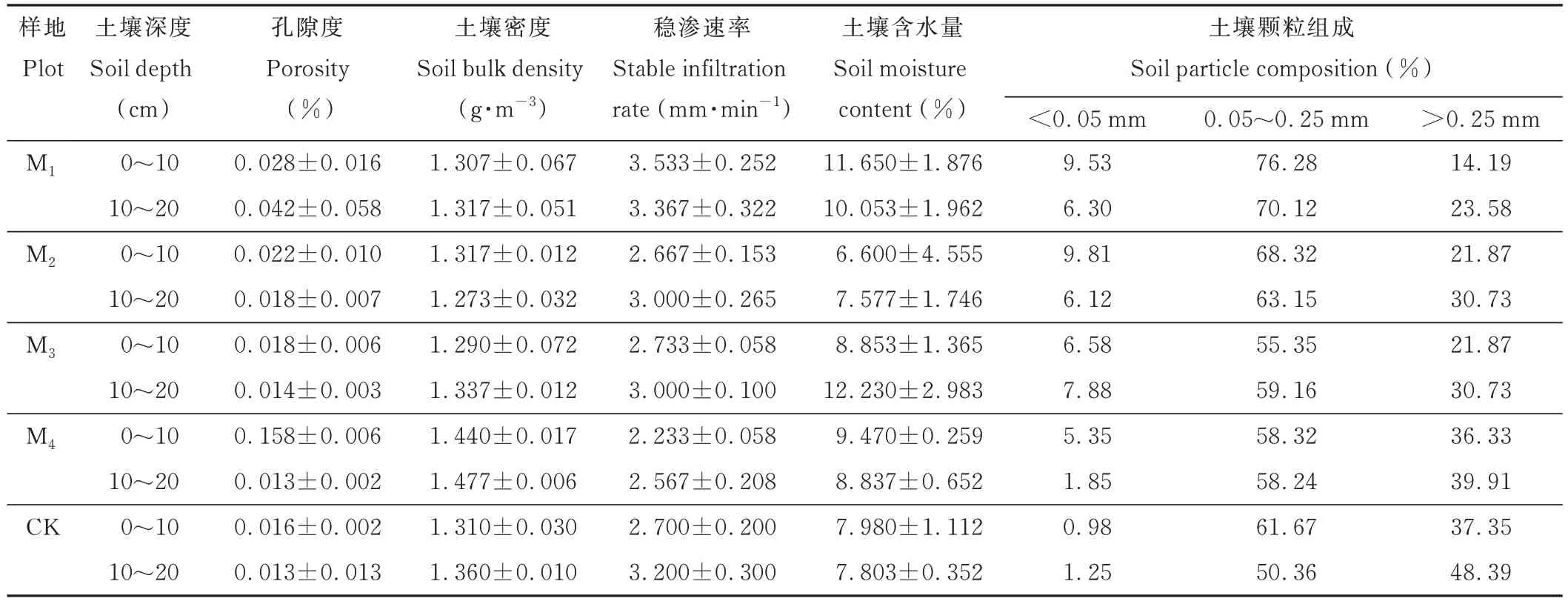
表2 样地土壤基本性质Table 2 Basic properties of the soil in the plot
1.3 数据分析
1.3.1 CT 扫描与图像处理 利用nanoVoxel-5000 系列双射线源CT 系统(图1,三英精密有限公司生产,像素细节分辨能力200 nm,空间分辨率200 nm,可测样品尺寸直径×高度不高于600 mm×600 mm)对36 个原状土样进行扫描。经CT 扫描后,每个原状土柱将得到2094 张横向切层图像。首先利用ImageJ软件对所有横切图进行预处理,再使用图像处理软件Avizo 2019.1(图2)对每个土柱的所有横切图像进行三维重构,得到各原状土样的几何分布图像。三维重建方法如下:首先导入二值化图像,依次进行整合处理、阈值分割,再通过剪裁功能修改单位,使重构模型接近真实试件,利用Volume-edit 模块截取圆柱并生成mask,最终得到体素为0.137 mm/pixel(X 方向)×0.137 mm/pixel(Y 方向)×0.2 mm/pixel(Z 方向)的三维立体图像。

图1 nanoVoxel-5000 系列双射线源CT 系统Fig.1 nanoVoxel-5000 series dual ray source CT system

图2 图像处理软件Avizo 2019.1 使用界面Fig.2 Image processing software Avizo 2019.1
1.3.2 根-土结构参数分析 在几何分布图像重建后,由于取样时土管边缘会反向给予土壤压力,从而使得边缘处土壤结构变化,因此需要去除这种边缘效应带来的误差。经过观察及分析,决定去除圆周1 mm、上下两端各1 mm 的数据,利用Avizo 2019.1 图形软件的ROI 工具将边缘1 mm 及上下两端1 mm 的图像数据去除。最终得到X、Y 轴-24~24 mm,Z 轴-32~32 mm 的土柱三维立体图像数据,并进行下一步的计算。
利用Avizo 2019.1 进一步对图像进行解析,构建土壤团聚体结构分布特征。提取所有粒径>0.25 mm 的团聚体,并计算相关参数及其在图像中的位置。最终将土壤团聚体数据整合成为Excel 文件格式,准备进行下一步计算与处理。土壤团聚体的阈值选取方法及各特征参数的具体计算方法如下:首先预制一个水稳性团聚体,同时结合土柱拆解后肉眼识别的方法,将确定为团聚体阈值范围内的土团确定为团聚体。勾画出土柱三维结构中的团聚体,使得土柱团聚体三维可视化,利用Avizo 2019.1 软件分析得到每个团聚体的长度、体积、表面积、等效直径、球度,并统计团聚体数量。统计得到各土柱中每1 mm 土层的团聚体数量密度、长度密度、体积密度、表面积密度、平均等效直径及平均球度。
1.4 统计分析
采用Excel 2016 对试验数据进行整理,采用SPSS 26 对数据进行方差齐性检验,数据不符合方差齐性,选择韦尔奇方差分析法(Welch’s anova)分析柠条种植年限对团聚体的影响,并用Games-Howell 法进行多重比较(α=0.05);采用R 4.1.2 中“vegan”包的“cor”函数进行Pearson 相关性分析,使用“vegan”包的“rda”函数进行主成分分析(principal component analysis, PCA);采用Origin 2021 和R 4.1.2 制图,结果中数值采用“平均值±标准误”表示。
2 结果与分析
2.1 不同林龄柠条林地土壤团聚体几何分布特征
土壤团聚体几何分布结果见图3,不同林龄下的团聚体几何分布特征差异明显:相较之下,35 年生柠条林地及草地对照样地土壤团聚体分布更为均匀,且分布密度高,而9 和14 年生林地土壤中团聚体分布的均匀性较差,土壤团聚体集中于部分区域,而在部分区域的分布密度较低。

图3 土壤团聚体几何分布特征Fig.3 Geometric distribution characteristics of soil aggregates
2.2 柠条林土壤团聚体数量及长度分布演变特征
对柠条灌丛引入后的土壤团聚体数量及长度特征演变规律进行分析(表3),可见,柠条种植后,除了14 年生柠条林地,其他林龄土壤团聚体数量密度整体上显著高于草地(P<0.05);0~10 cm 土层土壤团聚体数量密度表现为9 年生>24 年生>35 年生>14 年生>CK,10~20 cm 土层土壤团聚体数量密度随着林龄增加呈先增后减再增的趋势,总体上,0~20 cm 土层土壤团聚体数量密度表现为9 年生>35 年生>24 年生>14 年生。林龄与团聚体数量密度的相关系数为0.262(P<0.001)(图4),可见团聚体数量密度整体上与林龄成正比。通过图5a 可见,在地下0~10 cm,随着土壤深入,9 年生团聚体数量密度总体呈先增加后减少的趋势,其他组团聚体数量密度总体呈平缓的趋势;在地下10~20 cm,随着土壤深入,9、24、35 年生团聚体数量密度总体呈先增加后减少的趋势,草地对照团聚体数量密度总体呈先减少后增加的趋势,14 年生团聚体数量密度总体呈平缓增加的趋势。

图4 柠条林龄与土壤团聚体拓扑结构特征相关关系Fig.4 Correlation between different years of C. korshinskii and topological characteristics of soil aggregates
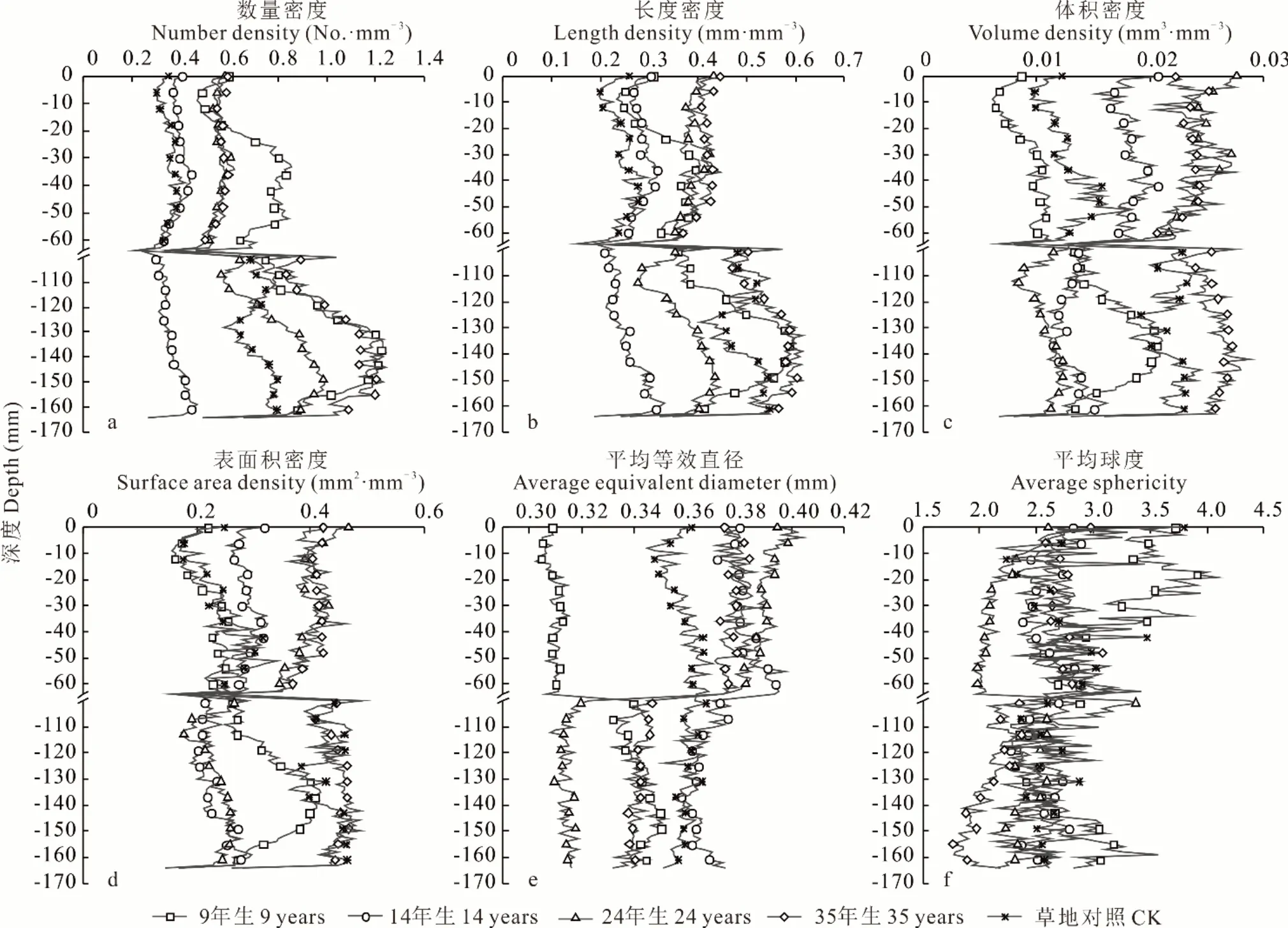
图5 不同林龄柠条林地土壤团聚体拓扑结构特征随土壤深度变化情况Fig.5 Topological characteristics of soil aggregates in C. korshinskii shrubland of different years vary with soil depth

表3 不同林龄柠条林地不同深度土层土壤团聚体拓扑结构特征Table 3 Topological characteristics of soil aggregates in soil layers at different depths in C. korshinskii shrubland of different years
由表3 可见,9 年生至35 年生土壤团聚体长度密度呈先减后增的趋势;在地下0~10 cm,团聚体长度密度整体上显著高于草地(P<0.05)。林龄与团聚体长度密度的相关系数为0.309(P<0.001)(图4),可见团聚体长度密度整体上与林龄成正比。由图5b 可以看出,在0~10 cm 土层 ,随着土壤深入,团聚体长度密度总体呈先增加后减少的趋势;在地下10~20 cm,随着土壤深入,9、24、35 年生团聚体长度密度总体呈先增加后减少的趋势,草地对照团聚体长度密度总体呈先减少后增加的趋势,14 年生团聚体长度密度总体呈平缓增加的趋势。
综上,不同林龄柠条林地土壤团聚体数量密度及长度密度存在显著差异,结合图6 可得,总体表现为随着林龄的增加,团聚体数量密度及长度密度随之增加;相比于表层土壤,深层土壤具有更大的团聚体数量密度及长度密度。

图6 柠条林龄与土壤团聚体拓扑结构特征主成分分析Fig. 6 Principal component analysis of topological characteristics of different years of C. korshinskii and soil aggregates
2.3 柠条林土壤团聚体体积及表面积分布演变特征
分析柠条灌丛引入后的土壤团聚体体积及表面积特征演变规律(表3)可知,草地至9 年生柠条林的团聚体体积密度下降;在地下0~10 cm,团聚体体积密度在9 年生至24 年生增加,而到35 年生减少;在地下10~20 cm,团聚体体积密度在9 年生至24 年生减少,而到35 年生增加。林龄与团聚体体积密度的相关系数为0.482(P<0.001)(图4),可知团聚体体积密度整体上与林龄呈正相关关系。由图5c 可以看出,在地下0~10 cm,随着土壤深入,9 年生团聚体体积密度总体呈增加的趋势,其他组团聚体体积密度总体呈先增加后减少的趋势;在地下10~20 cm,随着土壤深入,9、35 年生团聚体体积密度总体呈先增加后减少的趋势,14 年生和草地对照团聚体体积密度总体呈先减少后增加的趋势,24 年生团聚体体积密度总体呈增加的趋势。
由表3 可得,草地至9 年生柠条林的土壤团聚体表面积密度下降;在0~10 cm 土层,14、24、35 年生和草地对照团聚体表面积密度分别是9 年生的1.31、1.80、1.84、1.10 倍,9 年生至35 年生呈增加趋势;在10~20 cm 土层,9、24、35 年生和草地对照团聚体表面积密度分别是14 年生的1.46、1.02、1.92、1.88 倍,14年生至35 年生呈增加趋势。林龄与团聚体表面积密度的相关系数为0.368(P<0.001)(图4),可知团聚体表面积密度整体上与林龄呈正相关关系。团聚体体积密度与表面积密度相关性极强,相关系数达到0.948(P<0.001) (图4)。由图5d 可以看出,在地下0~10 cm,随着土壤深入,团聚体表面积密度总体呈先增加后减少的趋势;在地下10~20 cm,随着土壤深入,9、24、35 年生团聚体表面积密度总体呈先增加后减少的趋势,草地对照团聚体表面积密度总体呈先减少后增加的趋势,14 年生团聚体表面积密度总体呈增加的趋势。
综上,不同林龄柠条林地土壤团聚体体积密度及表面积密度存在显著差异,总体表现为随着林龄的增加,团聚体体积密度及表面积密度随之增加;相比于表层土壤,深层土壤具有更大的团聚体体积密度及表面积密度。
2.4 柠条林土壤团聚体平均等效直径及平均球度分布演变特征
对不同林龄柠条林地土壤团聚体平均等效直径及平均球度特征进行分析(表3)可知,草地至9 年生柠条林的团聚体平均等效直径下降;在地下0~10 cm,9年生至24 年生团聚体平均等效直径呈增加趋势;在地下10~20 cm,9 年生至35 年生团聚体平均等效直径总体呈先增后减再增趋势。林龄与团聚体平均等效直径的相关系数为0.108(P<0.01)(图4),可见整体上,团聚体平均等效直径随林龄增加而增加。由图5e 可以看出,在地下0~10 cm,随着土壤深入,14 年生和草地对照团聚体平均等效直径总体呈增加的趋势,24 年生团聚体平均等效直径总体呈减少的趋势,其他组团聚体平均等效直径总体呈平缓的趋势;在地下10~20 cm,随着土壤深入,9 年生团聚体平均等效直径总体呈先增加后减少的趋势,14 年生团聚体平均等效直径总体呈先减少后增加的趋势,其他组团聚体平均等效直径总体呈平缓的趋势。由图7 可以看出,不论是不同林龄的柠条林地,还是草地对照,平均等效直径在0.25~0.30 mm 的团聚体最多,并且随着团聚体平均等效直径增大,团聚体的占比逐渐变少,在0.35~0.40 mm 急剧减少。

图7 不同林龄土壤团聚体平均等效直径分布Fig. 7 Average equivalent diameter distribution of soil aggregates of different years
土壤团聚体球度越大,代表团聚体越规则,稳定性越强[35]。草地至9 年生柠条林的土壤团聚体平均球度增加(表3);在地下0~10 cm,9 年生至35 年生团聚体平均球度呈先减后增的趋势;在地下10~20 cm,9 年生至35 年生团聚体平均球度总体呈减少趋势。林龄与团聚体平均球度的相关系数为-0.384(P<0.001)(图4),可见整体上,林龄对团聚体平均球度有重要的负相关影响。由图5f 可以看出,在地下0~10 cm,随着土壤深入,14 年生团聚体平均球度总体呈先减少后增加的趋势,草地对照团聚体平均球度总体呈增加的趋势,35 年生团聚体平均球度总体呈平缓的趋势,其他组团聚体平均球度总体呈减少的趋势;在地下10~20 cm,随着土壤深入,9 年生团聚体平均球度总体呈增加的趋势,14 年生和草地对照团聚体平均球度总体呈增加的趋势,其他组团聚体平均球度总体呈减少的趋势。
综上,不同林龄柠条林地土壤团聚体平均等效直径及平均球度存在显著差异,总体趋势表现为随着林龄的增加,团聚体平均等效直径有所增加,团聚体平均球度有所减少;相比于深层土壤,浅层土壤具有更大的团聚体平均等效直径及平均球度。
3 讨论
相比以往研究中对土壤水稳性团聚体含量、团聚体平均质量直径(mean weight diameter, MWD)、几何平均直径(geometric mean diameter, GMD)和质量分形维数(fractal dimension, D)等指标的测量分析[13-22],本研究利用CT 扫描结合图像处理解析方法,获取了荒漠草原不同林龄柠条林地土壤团聚体的几何分布特征,并量化了原状土壤中团聚体的数量、长度、体积、表面积、平均等效直径、平均球度等拓扑结构特征参数,这有利于进一步认识土壤团聚体的发育过程、机制及其生态功能。同时,利用CT 扫描及图像处理技术获取土壤团聚体拓扑结构指标,可为今后土壤团聚体水文效应[36]、根系与土壤团聚体互作关系[37]等方面的研究提供技术支撑。
柠条林地土壤团聚体数量密度表现为9 年生>35 年生>24 年生>14 年生,可见,刚种植柠条的一定年限内,土壤团聚体含量会随着林龄增加先减少后增加。以往研究发现土壤大团聚体(>0.25 mm)含量总体表现为35年生柠条>25 年生>15 年生[19],而>0.25 mm 水稳性团聚体数量随着柠条种植年限增加出现先减小后增大的现象[20],本研究结果与其基本一致。在0~10 cm 土层,团聚体体积密度在林龄为14 年生时开始增加,而在10~20 cm 土层,体积密度在35 年生时才开始增加,可见,虽然大团聚体在深层含量多于浅层,但大团聚体在表层形成快于深层,柠条在该地区的表层恢复效果强于深层。以往研究发现植被恢复可显著改善土壤理化性质,但随着土层深度增加,土壤物理性质改善效果会减弱[38],土壤酶活性、土壤养分和土壤微生物多样性均呈降低趋势[39],本研究结果与其基本一致。本研究中土壤团聚体平均等效直径表现为CK>9 年生>35 年生>24 年生,可见大林龄柠条林地土壤团聚体平均等效直径要低于未种植柠条的草地,这与以往研究结果一致[21]。可见人工造林会导致土壤团聚体平均直径降低,可能是因为柠条的引入提高了地下生态系统功能,促进了小直径土壤团聚体的形成,导致微团聚体数量增多,从而使得柠条林地土壤团聚体平均直径降低[40]。
对比近35 年柠条林地土壤团聚体几何分布特征发现,随着柠条林龄增加,土壤团聚体密度逐渐增大且分布逐渐均匀,土壤结构改善效果良好,可见柠条的种植可以很好地改善土壤的结构,这与以往的研究结果基本一致[22,41-42]。虽然林地表层土壤团聚体数量、体积少于深层土壤,但表层团聚体的平均球度大、平均等效直径大,因此可以认为表层土壤的团聚体稳定性更强,这与以往的研究结果一致[43-44],因此,在对土地资源利用及柠条林管理时不能仅考虑团聚体数量、大小而忽略土壤团聚体分布及拓扑结构改变带来的重要影响。此外,今后研究中需要充分考虑人工林生长过程中林地土壤团聚体几何分布与拓扑结构特征演变引起的生态功能改变,从而进行针对性、精细化管理,以提高人工林生态恢复的高效性及可持续性。
4 结论
1)利用CT 扫描及图像处理技术可以量化土壤团聚体几何分布及拓扑结构特征,可为今后土壤团聚体水文效应、根系与土壤团聚体互作关系等方面的研究提供技术支撑。
2)人工柠条引入及林龄增加会导致土壤团聚体数量增加、表层大团聚体形成、拓扑结构优化,需要关注人工林生长过程中林地土壤团聚体几何分布与拓扑结构特征演变引起的生态功能改变。
3)在人工柠条林地,虽然0~10 cm 土层土壤团聚体数量、体积小于10~20 cm 土层土壤,但0~10 cm 土层土壤团聚体平均粒径及球度更大,可见林地表层土壤具有更少但更稳定的团聚体,在今后相关研究中,不仅需要分析团聚体数量及大小,同时不能忽略可表征团聚体稳定性的拓扑结构参数的特征与变化。
猜你喜欢
现代农村科技(2021年5期)2021-12-07
中学生数理化·高一版(2021年2期)2021-03-19
中国煤炭工业(2020年1期)2020-12-28
防护林科技(2020年10期)2020-12-22
今日农业(2020年23期)2020-12-15
中国外汇(2019年6期)2019-07-13
当代陕西(2019年7期)2019-04-25
中学生数理化·高一版(2017年2期)2017-04-25
防护林科技(2016年9期)2016-09-27
防护林科技(2016年5期)2016-09-05
