
放牧条件下萨福克与河谷型藏绵羊杂交F1 代不同部位肌肉营养品质分析
2023-11-22 09:38刘孟君任越
草业学报 2023年11期
刘孟君,任越*
(1. 西藏自治区农牧科学院畜牧兽医研究所,西藏 拉萨 850000;2. 农业农村部青藏高原畜禽遗传育种重点实验室,西藏 拉萨 850000)
畜牧业是西藏的主导产业,以西藏羊为基础的衍生产业是西藏发展畜牧业关注的重点[1],也是发展农牧业经济、增加收入的关键。随着市场的调整,西藏羊因自身存在的缺点(如体格小、产肉率低、生长速率慢、饲养周期长),已无法完全满足当前消费者的需要[2]。针对这一现象,目前以萨福克羊改良河谷型藏绵羊的商品羊的养殖模式在年楚河中下游区域取得了一定成效。河谷型藏绵羊在西藏境内主要分布于雅鲁藏布江、拉萨河、年楚河中下游的农牧结合区域[3],其羊肉肉质鲜美、蛋白含量高、无膻味。得益于独特的高原环境,以“绿色”为品牌打造的羊肉产品引起了市场的广泛关注。当前对藏绵羊肉品质的研究主要集中在整体营养价值的评定方面[4],对于不同部位肉品质的研究相对较少,羊肉作为西藏重要优质肉类来源,相比于内地较发达的牧区,在分档研究方面的工作还处于初步阶段。传统整只售卖的方式,降低了养殖户利润,限制了消费者选择的空间,面对西藏羊肉产业长期缺乏系统的胴体分割技术的问题,依据羊胴体不同部位肉品质的特性进行分割,有助于提高藏羊的附加值及种质资源利用价值,推进西藏肉羊产业的发展进度。然而,分割分档的方式依据胴体不同部位营养组成及品质不尽相同,欧慧敏等[5]研究发现,呼伦贝尔羔羊的肉品质和营养组成,前肩、前肢部位优于背最长肌、后肢、颈部;罗玉龙等[6]研究指出小尾寒羊股二头肌、背最长肌、臂三头肌3 个部位的脂肪含量、熟肉率差异不显著,肉色、嫩度、pH 差异显著,最后以饱和脂肪酸与不饱和脂肪酸比值为评价指标,认为臂三头肌部位的肉质较好;周雨[7]研究发现苏格绵羊后腿、前腿、背最长肌存在差异,后腿部位肌肉氨基酸总量最高,背最长肌的总脂肪酸、饱和脂肪酸、不饱和脂肪酸含量最高且肉色最佳;刘孟君等[8]研究发现,河谷型藏绵羊的股二头肌的口感与鲜嫩程度优于臂三头肌、背最长肌,从风味物质、氨基酸及脂肪酸组成上看,臂三头肌、股二头肌比背最长肌更符合人体对营养的需求。从上述试验结果不难看出,不同部位肉品质和营养组成确实存有差异。
西藏具有大规模的河谷型藏绵羊群体数量,利用萨福克羊开展一元经济杂交所产后代的不同部位的肉品质和营养组成有何差异尚不清楚。为此,以放牧条件下饲养的萨福克与河谷型藏绵羊杂交F1代为研究对象,测定其基本营养成分、微量元素、维生素、氨基酸、脂肪酸,比较分析杂交后F1代的背最长肌、臂三头肌、股二头肌的肉质营养特性,从而为杂交F1代羊肉的分级分割及标准化生产提供理论依据。
1 材料与方法
1.1 试验时间与地点
试验于2019 年3 月至2020 年10 月在山南市隆子县热荣乡沃塘村(28°30'37.32'' N,92°05'52.26'' E)进行,放牧地海拔4651 m,属于高原温带大陆性季风气候。放牧区域为高山草原草地、高山草甸类型,植被组成以莎草-禾本科为主,常见植物有高山嵩草(Kobresia pygmaea)、羊茅(Festuca ovina)、藏北嵩草(Kobresia littledalei)、青藏薹草(Carex moorcroftii)、低株披碱草(Elymus jacquemontii)、尼泊尔大丁草(Leibnitzia nepalense)、钉柱委陵菜(Potentilla saundersiana)、无茎黄鹌草(Youngia simulatrix)、藏布红景天(Rhodiola sangpotibetana)、禾叶点地梅(Androsace graminiflia)、弯茎还阳参(Crepis flexuosa)等。
1.2 试验动物与饲养管理
随机选取50 只体型相近[体重:(11.64±2.34)kg]的3 月龄断奶萨福克与河谷型藏绵羊杂交F1代公羔(图1),打耳标,白天随羊群自由放牧,晚上归牧后不进行补饲。试验羔羊连续放牧饲养15 个月,直至1.5 岁,体重达到(35.62±6.46) kg,放牧场可食牧草主要有高山嵩草、藏北嵩草、青藏薹草、羊茅,其营养成分见表1。

表1 主要采食牧草的营养水平Table 1 Nutrient levels of the main forage grasses (dry matter basis)

图1 萨福克与河谷型藏绵羊杂交F1代Fig. 1 F1 generation of Suffolk crossed with river valley type Tibetan sheep
1.3 样品采集与处理
于2020 年10 月中旬随机选取接近组内平均体重的10 只体况、体型基本一致的杂交F1代,屠宰前12 h禁食,宰前2 h 禁水,屠宰后取背最长肌、臂三头肌、股二头肌各0.5 kg,4 ℃冷藏后真空封装于保鲜袋,置于-18 ℃冰箱冷冻保存,待检。
1.4 指标检测与方法
1.4.1 肉品质 pH 测定:屠宰45 min 后,用便携式pH 计(BJ-260,INESA 科学仪器,上海,中国)在样品处用探针插入2 cm 深度连续测定3 次pH 值,取平均值作为计算结果记为pH45min;4 ℃静置排酸24 h 后在背最长肌、臂三头肌、股二头肌处测定pH,记为pH24h;肉色测定:将色彩色度计(CR-10,KONICA MINOLTA,日本)调至L*(亮度)、a*(红度)、b*(黄度)色域系统,对样品随机选择5 个点分别测定亮度(L*)、红度(a*)和黄度(b*),取平均值;熟肉率测定:取肉样,将其切割为2 cm×2 cm×3 cm=12 cm3肉块称重记为重量M1,放入85 ℃恒温水浴锅,蒸煮30 min,待肉块冷却至室温,用纸吸干表面水分称重记为M2,设置3 个平行,M2/M1×100%=熟肉率。
1.4.2 常规营养成分 肌肉水分含量测定:参照 GB 5009.3-2016《食品中水分的测定》[9]的冷冻干燥法。粗蛋白含量测定:参照 GB 5009.5-2016《食品中蛋白质的测定》[10]的凯氏定氮法。粗脂肪含量测定:参照GB 5009.6-2016《食品中脂肪的测定》[11]的索氏提取法。粗灰分含量测定:参照GB 5009.4-2016《食品中灰分的测定》[12]的总灰分测定法。
1.4.3 维生素A、维生素E、维生素B1、维生素B2、胆固醇 维生素A、E 含量测定:参照GB 5009.82-2016《食品中维生素A、D、E 的测定》[13]的反相高效液相色谱法。维生素B1 含量测定:参照GB 5009.84-2016《食品中维生素B1 的测定》[14]的高效液相色谱法。维生素B2 含量测定:参照GB 5009.85-2016《食品中维生素B2 的测定》[15]的高效液相色谱法。胆固醇含量测定:参照GB 5009.128-2016《食品中胆固醇的测定》[16]的高效液相色谱法。
1.4.4 肌肉微量元素 肌肉内钙、铁、锌、硒元素含量采用电感耦合等离子体发射光谱法测定。采用微波消解法[17],取冷冻干燥的肉样粉末1 g,置于消解内罐中,加入5 mL 硝酸,放置1 h,再放入150~170 ℃恒温干燥箱内消解4 h,在超声水浴箱中脱气2~5 min,用水定容至25 mL,留待备用。将标准工作液注入电感耦合等离子体发射光谱仪中,绘制出标准曲线,将空白溶液和试样溶液分别注入电感耦合等离子体发射光谱仪中,根据标准曲线得到消解液中待测元素的浓度。
1.4.5 肌肉脂肪酸含量 取冷冻干燥肉样粉末0.5 g,置于10 mL 玻璃离心管中,用氯仿-甲醇(体积比为2∶1)混合溶剂提取脂肪,分别用酸和碱对提取的试样进行甲基化。在脂肪酸甲基化之前,用1 mL 正癸酸甲酯(11∶0)作为内标(1 mg·mL-1)生成脂肪酸甲酯。经配备火焰离子化检测器和熔融石英毛细管柱的气相色谱分析,内标法[18]定量测定脂肪酸甲酯含量,依据各种脂肪酸甲酯含量和转换系数计算出总脂肪酸、饱和脂肪酸、单不饱和脂肪酸、多不饱和脂肪酸含量。气相色谱的柱温箱升温程序为初始温度120 ℃,保持5 min,以3 ℃·min-1的速度增加到230 ℃,保持3 min;以1.5 ℃·min-1的速度增加到240 ℃并保持13 min;运行时间48 min。使用氮气作为载气,流速为1 mL·min-1,进样体积为1.0 μL,进样口温度为260 ℃。
1.4.6 肌肉氨基酸含量 氨基酸含量测定:参照GB 5009.124-2016《食品中氨基酸的测定》[19]测定肉样的16种氨基酸及采用高效液相色谱仪测量半胱氨酸含量[20]。
1.4.7 氨基酸评分计算方法 根据联合国粮农组织/世界卫生组织(Food and Agriculture Organization of the United Nations/World Health Organization, FAO/WHO)1973 年建议的氨基酸评分标准模式和全鸡蛋蛋白质的氨基酸模式,按下式计算氨基酸评分(amino acid score, AAS)、化学评分(chemical score, CS)和必需氨基酸指数(essential amino acid index, EAAI):
式中:n为比较的必需氨基酸个数;A,B, …,H为待评价样品蛋白质中某种必需氨基酸含量;a,b, …,h为全鸡蛋蛋白质中同种必需氨基酸含量。
1.5 数据统计与分析
采用 SPSS 21.0 统计软件对试验数据进行单因素方差分析(One-way ANOVA),采用Duncan 氏法进行多重比较,结果用平均值±标准差(mean±SD)表示,P<0.05 表示差异显著。
2 结果与分析
2.1 杂交F1代不同部位肌肉品质
杂交F1代不同部位肉的食用品质差异见表2,背最长肌、臂三头肌的肉色L*值显著高于股二头肌(P<0.05),背最长肌、股二头肌的肉色a*值显著高于臂三头肌(P<0.05),背最长肌的肉色b*值显著高于臂三头肌、股二头肌(P<0.05)。3 个部位肌肉的熟肉率无显著差异(P>0.05)。臂三头肌pH45min最高,为5.85,显著高于背最长肌、股二头肌(P<0.05);3 个部位肌肉pH24h无显著差异(P>0.05)。

表2 杂交F1代不同部位肌肉品质Table 2 Muscle quality of different parts of the F1 generation of the hybrid progeny
2.2 杂交F1代不同部位肌肉基础营养成分
杂交F1代不同部位肌肉的基本营养成分见表3,背最长肌的粗蛋白含量显著高于臂三头肌(P<0.05),股二头肌的粗蛋白含量与背最长肌、臂三头肌无显著差异(P>0.05);3 个部位肌肉之间的水分、粗脂肪、灰分含量无显著差异(P>0.05)。
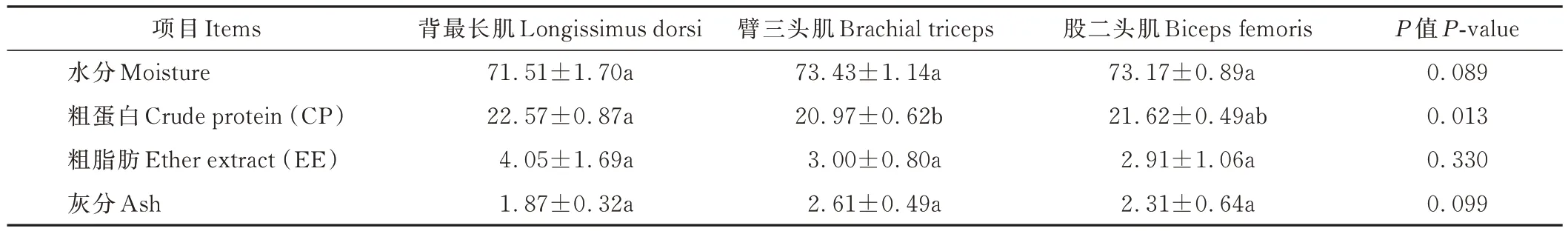
表3 杂交F1代不同部位肌肉常规营养水平Table 3 Conventional nutrient composition of different parts of muscle in the F1 generation of the hybrid progeny (%)
2.3 杂交F1代不同部位肌肉钙、铁、锌、硒元素含量
杂交F1代不同部位肌肉的钙、铁、锌、硒元素含量见表4,臂三头肌锌元素含量显著高于背最长肌、股二头肌(P<0.05), 背最长肌、臂三头肌、股二头肌在钙、铁、硒元素含量上均无显著性差异(P>0.05)。

表4 杂交F1代不同部位肌肉钙、铁、锌、硒元素含量Table 4 Content of calcium, iron, zinc and selenium in the muscles of different parts of the F1 generation of the hybrid progeny(dry matter basis, mg·kg-1)
2.4 杂交F1代不同部位肌肉胆固醇、VA、VE、VB1、VB2含量
杂交F1代不同部位肌肉的胆固醇、维生素A、维生素E、维生素B1、维生素B2 含量见表5,背最长肌、股二头肌、臂三头肌在胆固醇、维生素A、维生素E、维生素B1、维生素B2 含量上均无显著性差异(P>0.05)。
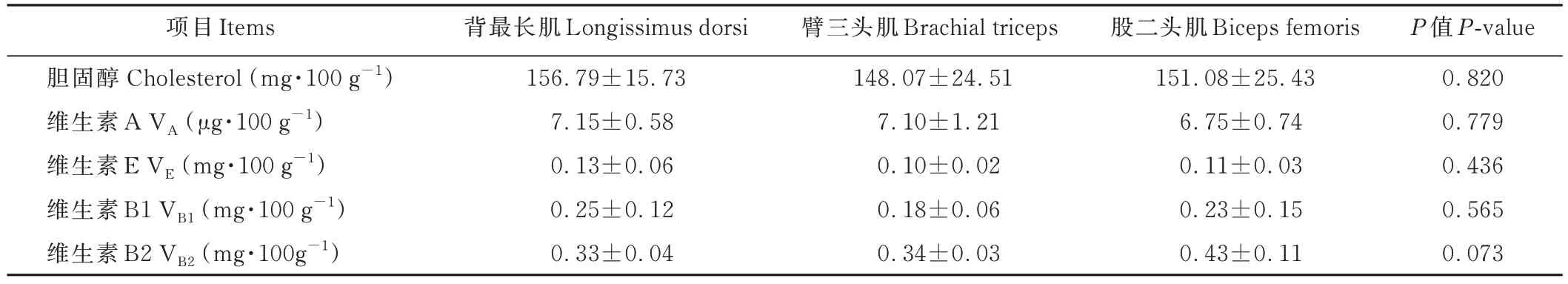
表5 杂交F1代不同部位肌肉胆固醇、VA、VE、VB1、VB2含量Table 5 Cholesterol, vitamin A, vitamin E, vitamin B1 and vitamin B2 content of different parts of muscle in the F1 generation of the hybrid progeny (dry matter basis)
2.5 杂交F1代不同部位肌肉氨基酸组成及含量
如表6 所示,在杂交F1代的背最长肌、臂三头肌、股二头肌3 个部位均检测出了7 种必需氨基酸(Thr、Val、Met、Ile、Leu、Phe、Lys)、10 种非必需氨基酸(Asp、Ser、Glu、Pro、Gly、Ala、Cys、Tyr、His、Arg)。其中臂三头肌的谷氨酸含量显著高于背最长肌(P<0.05),背最长肌的组氨酸含量显著高于股二头肌、臂三头肌(P<0.05),除此之外,检测出的其余氨基酸含量及必需氨基酸、非必需氨基酸总量均无显著性差异(P>0.05)。3 个部位检测出来的风味氨基酸有鲜味类氨基酸(天冬氨酸、谷氨酸),苦味类氨基酸(异亮氨酸、络氨酸、缬氨酸、亮氨酸、苯丙氨酸),甜味类氨基酸(丙氨酸、甘氨酸、丝氨酸、脯氨酸),芳香类氨基酸(络氨酸、苯丙氨酸),鲜味类、苦味类、甜味类、芳香类氨基酸的含量在背最长肌、股二头肌、臂三头肌之间有差异,但不显著;就鲜味类、苦味类、甜味类、芳香类氨基酸含量总量而言,臂三头肌>股二头肌>背最长肌。

表6 杂交F1代不同部位肌肉的氨基酸组成及含量Table 6 Amino acid composition and content of muscles in different parts of the F1 generation of the hybrid progeny(dry matter basis, %)
如表7 所示,对杂交F1代的背最长肌、股二头肌、臂三头肌进行必需氨基酸评分(AAS)、化学评分(CS)时发现蛋氨酸的必需氨基酸评分和化学评分最低,缬氨酸次之,故各部位肌肉第一限制性氨基酸均为蛋氨酸(Met),第二限制性氨基酸为缬氨酸(Val),从必需氨基酸化学评分来看臂三头肌、股二头肌、背最长肌接近,分别为60.68、61.96、60.07,与标准蛋白接近。从氨基酸角度评价来看,杂交F1代的背最长肌、股二头肌、臂三头肌的营养成分差异不大。
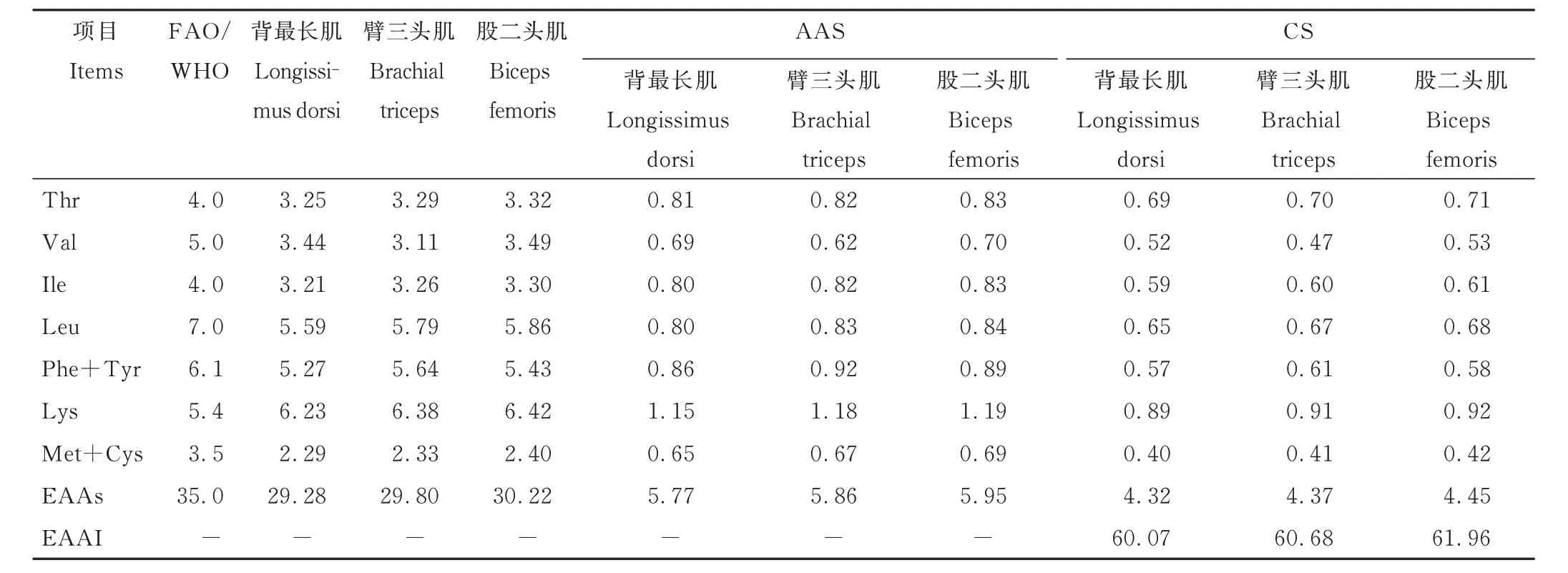
表7 杂交F1代不同部位肌肉的必需氨基酸评分Table 7 Essential amino acid scores of different parts of muscle in the F1 generation of the hybrid progeny
2.6 杂交F1代不同部位肌肉脂肪酸组成及含量
如表8 所示,除棕榈酸(C16:0)、花生四烯酸(C20:4n6)外,杂交F1代的背最长肌、臂三头肌、股二头肌的其余种类脂肪酸均无显著性差异(P>0.05);背最长肌、股二头肌的棕榈酸含量显著高于臂三头肌(P<0.05),臂三头肌的花生四烯酸含量显著高于背最长肌(P<0.05)。背最长肌、股二头肌、臂三头肌脂肪酸组成规律一致,为∑SFA>∑MUFA>∑PUFA,饱和脂肪酸(saturated fatty acid, SFA)、单不饱和脂肪酸 (monounsaturated fatty acids, MUFA)、多不饱和脂肪酸(polyunsaturated fatty acids, PUFA)含量在3 个部位间差异不显著(P>0.05),其中饱和脂肪酸主要由棕榈酸、硬脂酸组成,单不饱和脂肪酸主要由油酸组成,多不饱和脂肪酸主要由亚油酸、α-亚麻酸、花生四烯酸组成。
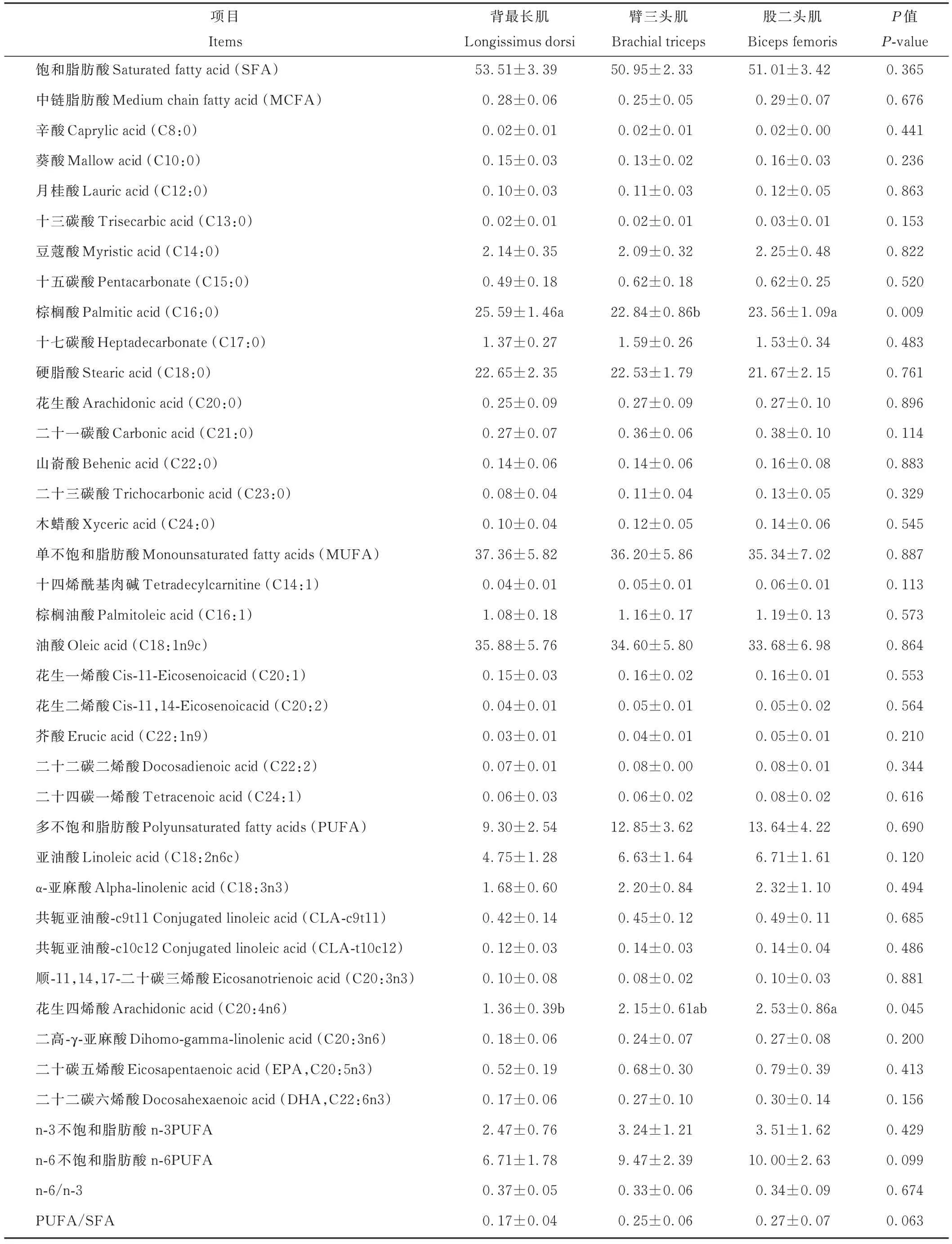
表8 杂交F1代不同部位肌肉的脂肪酸组成及含量Table 8 Fatty acid composition and content of muscles in different parts of the F1 generation of the hybrid progeny (dry matte basis, mg·g-1)
3 讨论
3.1 杂交F1代不同部位肌肉的肉品质评价
肉色是判断肉胴体表观的重要指标,直观反映出肉产品的质量状况,受年龄、饲料、取样部位、肌纤维类型、肌红蛋白含量、肌内脂肪含量等多种因素影响[21],其中a*与肌内肌红蛋白、血红蛋白含量有关[22],b*与脂肪沉积、肌内肌红蛋白含量、饲料色素有关[22],L*与肌内水分含量、肌内纤维类型有关[23]。杂交F1代背最长肌的a*、b*、L*值最高,臂三头肌的a*、b*最低,股二头肌的L*最低,与周雨[7]对苏格绵羊、罗玉龙等[6]对小尾寒羊、柰如嘎[24]对阿拉善绵羊的研究结论相似。研究表明[21-22],肉色主要取决于肌内纤维中的肌红蛋白含量,其含量可以决定肉色鲜红程度的80%~90%,氧化型肌纤维富含肌红蛋白色泽好,酵解型肌纤维肌红蛋白含量少色泽显苍白,受年龄、环境、运动、饲养管理等因素影响,肌纤维会由氧化型向酵解型转化,本研究中背最长肌的a*、b*、L*值均高于臂三头肌、股二头肌,放牧条件下,羊采食一直处于攀爬状态,背部肌肉的运动量小于后腿、前腿[7],背部肌肉纤维转化为酵解型纤维的程度低于后腿、前腿。熟肉率是衡量肉品系水力的重要指标,与肉的多汁性、嫩度、香味、外观有关,通常熟肉率越高,肉产品的出品率越高,加工品质就越好[6],本试验中,萨藏杂交F1代3 个不同部位的熟肉率无明显差异,与罗玉龙等[6]、金艳梅等[25]的报道相近,但明显高于李旺平等[26]开展不同品种杂交肉羊的肉品质研究的结果:澳洲白公羊×湖羊母羊[(54.25±2.86)%]、杜泊公羊×湖羊母羊[(53.19±2.63)%]、澳洲白公羊×湖寒母羊[(54.26±2.45)%]、杜泊公羊×湖寒母羊[(54.38±2.95)%]、澳洲白公羊×杜湖母羊[(55.21±3.01)%]、杜泊公羊×杜湖母羊[(55.09±2.46)%],推测引起差异的主要原因可能是屠宰年龄不同[27]。胴体pH是评定肉品质的重要指标之一,动物屠宰后,在肌肉向肉转化过程中,血液循环停止,糖酵解最终产物乳酸不断在肌肉中积累,直至停止[28],影响肉品的嫩度、肉色、系水性能,主要与不同类型肌纤维的糖酵解潜力有关[21]。本试验中,整体来看,24 h 背最长肌、臂三头肌、股二头肌的pH 相比于45 min 有所下降,其中,pH45min时臂三头肌的pH显著高于背最长肌、股二头肌,pH24h3 个部位无明显差异,可见臂三头肌的糖酵解潜力大于背最长肌、股二头肌,马晓冰等[29]报道指出巴美肉羊的背最长肌与股二头肌的糖酵解潜力无差异,但显著高于臂三头肌;小尾寒羊的股二头肌与臂三头肌的糖酵解潜力无差异,但显著低于背最长肌;苏尼特羊的背最长肌的糖酵解潜力显著高于股二头肌、臂三头肌,股二头肌糖酵解潜力又显著高于臂三头肌,本研究得出的结论与其具有差异。林在琼等[30]、苏琳等[31]研究报道指出背最长肌的酵解型肌纤维含量显著高于股二头肌、臂三头肌,富含糖原,代谢类型以无氧酵解为主,Choe 等[32]研究发现肌肉纤维类型组成直接影响屠宰后肌肉的能量代谢,快速酵解型肌肉的纤维含量与肌肉的糖酵解潜力呈正相关,而富含氧化型纤维肌肉的糖酵解潜力较低,由此可知,杂交F1代在高寒高海拔牧区放牧饲养模式下,背最长肌的糖酵解潜力相比于前腿、后腿较高,可能是由于运动量不一致。
3.2 杂交F1代不同部位肌肉的营养成分分析
肉产品的基本营养成分不仅能大致反映出其食用价值,而且还对肉的口感、风味、嫩度有重要参考作用,水分含量与肉持水性能、熟肉率相关,肌内脂肪(intramuscular fat, IMF)含量则与嫩度、多汁性、食用品质相关,有研究表明IMF 的增加可明显改善肉品多汁性,使肉在烹饪中保留更多水分[33],IMF 对于肌肉嫩度的改善主要是存在于肌纤维束之间的肌束膜结缔组织中的肌内脂肪,降低了肌纤维密度,使肉咀嚼时更易破碎[34]。本研究中,检测出的粗脂肪含量在3 个部位间差异不显著,李晓亚等[35]研究指出外源性脂肪酸的摄取及机体内自身合成的脂肪酸在不同组织间的沉积具有主次性,如背部皮下脂肪受日粮的影响较其他身体部位更大,Rouse 等[36]研究指出组织成熟的顺序为骨、肌肉、脂肪,据实际观测,杂交F1代在屠宰时还处于生长发育阶段,且在放牧条件下采食量及营养均衡均受到限制,肌肉并未到脂肪沉积的最佳时间点,同时Tateo 等[37]研究发现IMF 沉积与运动呈负相关,臂三头肌、股二头肌略低于背最长肌,原因可能是羊在高海拔放牧生境下采食时,臂三头肌、股二头肌作为主要运动部位之一,运动量过大,可见屠宰年龄、营养水平、饲养模式可能是3 个部位无差异的主要原因。Mottram 等[38]、Sañudo 等[39]研究指出IMF 作为肉品风味形成的重要前体参与物质,其含量在3.5%~4.5%时肉品口感最佳,本试验中,3 个部位的IMF 含量均在此范围内,其中背最长肌相比于臂三头肌、股二头肌的粗蛋白含量更高。
3.3 杂交F1代不同部位肌肉的钙、铁、锌、硒元素含量分析
矿物元素是机体组织及正常生理活动的重要参与者,不能由机体本身合成,只能从外界获取[21],机体内矿物元素的营养状况直接影响生长发育、营养物质代谢、酶的活性、繁殖、免疫应答,羊肉中微量元素组成及含量作为评价羊肉营养价值的重要组成部分,不仅是人体微量元素的重要来源,也是影响肉品质如肉色、抗氧化特性的主要营养素。本研究检测的钙、铁、锌、硒元素,除臂三头肌的锌元素显著高于背最长肌、股二头肌外,其余3 种元素无显著性差异。刘孟君等[8]对河谷型藏绵羊不同部位肉品质评价的报道中,钙、锌元素在臂三头肌、股二头肌、背最长肌无差异,臂三头肌的铁、锌元素的含量高于背最长肌。本研究与其既有相似也有差异,可见在引进萨福克羊对西藏河谷型藏绵羊进行杂交改良时,某些矿物元素在不同肌肉部位的分布也会受到不同程度影响[40],周艳等[41]研究指出肌肉中微量元素含量与品种、饲养方式、肌肉组织类型、微量元素种类变化有关,并表明不同品种肉羊对微量元素消化吸收富集效果不一样,可知,影响羊肉中微量元素分布规律的因素很多,需要进一步研究探讨。
3.4 杂交F1代不同部位肌肉的胆固醇及维生素含量分析
胆固醇来源途径为外源性摄入、内源性合成,机体内所需的胆固醇大部分由机体自身合成,额外的外源性摄入几乎全来自动物性食物[42],对于机体自身的胆固醇含量主要受遗传和环境因素的协同影响,因胆固醇可提高人体内低密度脂蛋白的含量,过量摄入胆固醇可引发心血管疾病,近年来低胆固醇的肉类食品备受青睐。本研究中背最长肌、臂三头肌、股二头肌的胆固醇含量并无显著差异,刘璐等[43]研究指出苏尼特羊肌肉组织中胆固醇含量变化规律不一致,具有组织特异性,如羯羔的臀肌低于背最长肌、股二头肌、臂三头肌,母羔的背最长肌低于股二头肌、臂三头肌、臀肌,本研究中3 个部位的胆固醇含量(干物质基础)低于王金文等[44]对5 月龄的鲁西黑头羊(59.19)、小尾寒羊(63.72)鲜样测量的报道,高于杨开伦等[45]对陶赛特与小尾寒羊杂交后代鲜样测量的结果,与张灿等[46]对自然放牧方式下欧拉羊鲜样测量结果相似,可见肌肉内胆固醇含量与品种、屠宰年龄、饲养方式均有关,就本研究而言,萨藏杂交F1的3 个不同部位肌肉的胆固醇含量较低。维生素作为维持动物健康不可或缺的脂溶性物质,其对肝脏、胰脏、肌肉、脂肪组织中的碳水化合物、脂质、蛋白质的代谢起作用[47],如维生素E 作为天然抗氧化剂,通过保护细胞膜的流动及稳定性,维持细胞膜完整性,提高肌肉抗氧化能力、阻止肉质氧化酸败、增加熟肉率、改善肉品质[48];维生素A 只存在于动物体中,作为碳水化合物、蛋白质和脂质代谢的重要调节剂[49],在细胞发育、调节、增殖和分化中起作用[50];维生素B1 主要对新陈代谢具有调节和控制作用,有研究表明脂肪可降低机体对维生素B1 的依赖性,而能量会增加机体对维生素B1 的需要量[51],同时,维生素B1 具有较高的遗传性,当母体缺乏会直接影响子代的生长发育[52];维生素B2 是维持机体健康、正常生长发育必需的维生素,通过其形成的辅酶,是氧化酶系统必要的组成部分,主要参与机体氧化过程中氢的传递,促进核酸、脂肪、蛋白质、糖类的代谢[53],广泛存在于各类食品,通常动物性食品含量高于植物性食物。对于反刍动物来说,维生素E、维生素A 的来源主要为采食的牧草,B 族维生素通过瘤胃微生物合成及食物获取,本试验中杂交F1代的背最长肌、股二头肌、臂三头肌在胆固醇、维生素A、维生素E、维生素B1、维生素B2 含量上均无显著性差异,与刘孟君等[8]报道一致,再次佐证了——肌肉中维生素含量与饲粮有关,饲粮来源及维生素组成一致,肌肉中维生素含量也大致相同[21]。
3.5 杂交F1代不同部位肌肉的氨基酸组成及含量分析
肌肉蛋白质分解与合成代谢过程均与肉品质的形成、风味物质的结合、肉色的稳定性有关[54],风味是评判肉类制品独特的、重要的感官属性,其吸附的主要载体则是蛋白质,Chen 等[55]研究发现赋予肉品的风味物质主要是由蛋白质降解过程产生的游离氨基酸、小分子肽,可见氨基酸的组成与含量不仅可以评价肌肉的蛋白质营养价值,还可以评价肉品的风味。本研究中,3 个部位均检测出7 种EAA、10 种NEAA,氨基酸种类组成较齐全,所检测出的EAA 中,亮氨酸、赖氨酸的含量相对较高,检测出的NEAA 中,谷氨酸、天冬氨酸的含量相对较高。谷氨酸、天冬氨酸是形成肉品香味所必需的前体氨基酸,肉品的鲜美程度取决于这类氨基酸含量高低,通常这类氨基酸含量越高,肉品鲜美程度越好,本研究中鲜味氨基酸的报道结果与张灿等[46]对欧拉羊报道结果较为一致,高于欧慧敏等[5]对呼伦贝尔羔羊、曾勇庆等[56]对小尾寒羊、吴法平等[57]对茶卡羯羊、吴铁梅等[58]对绒山羊研究的报道。进一步研究发现,臂三头肌的谷氨酸含量显著高于背最长肌,与欧慧敏等[5]对呼伦贝尔羔羊得出的研究结论一致,罗玉龙等[59]研究指出不同部位AMPD1、ADSL、ATIC基因表达量不同,运动量大的部位3 种基因表达量相对较高,可进一步调控形成鲜味物质的酶来提高羊肉的鲜味物质含量,本研究中鲜味氨基酸(Glu、Asp)的含量相较于其他报道的结果偏高,一方面可能与高寒高海拔的牧草营养成分有关,羊采食高寒高海拔的优质牧草富含抗氧化剂,肌肉组织的细胞完整性能最大程度保留,减少了风味物质的损耗[59];另一方面运动加大了肌细胞ATP 合成速率,提高了风味物质合成能力。就整体检测的结果来说,除Glu、His 含量有所差异外,3 个部位肌肉的其余种类氨基酸含量、EAA、NEAA、EAA/NEAA、EAA/TAA、鲜味类氨基酸、苦味类氨基酸、甜味类氨基酸、芳香类氨基酸的含量在统计学上差异不显著,与现有一些报道的结果不一致,如曾勇庆等[56]、欧慧敏等[5]、邱翔等[60]、原振清等[61]研究报道均指出了不同部位的各种氨基酸含量具有差异性,本研究中3 个部位的氨基酸含量较为一致,是由于放牧饲养模式下屠宰年龄未到引起还是由于高原适应引起,还需要进一步研究探讨。但就鲜味类、苦味类、甜味类、芳香类氨基酸总体含量而言,臂三头肌>股二头肌>背最长肌。由表7 可知,各部位肌肉的第一、第二限制性氨基酸均为蛋氨酸、缬氨酸,与闫忠心等[62]、刘孟君等[8]的研究结果一致。背最长肌、臂三头肌、股二头肌3 个部位的必需氨基酸指数分别为60.07、60.68、61.96,与标准蛋白接近,必需氨基酸营养价值差异不大。综合可见,放牧条件饲养模式下,3 个部位蛋白营养价值差异不显著,杂交F1代臂三头肌的鲜味氨基酸含量略高于股二头肌、背最长肌。
3.6 杂交F1代不同部位肌肉的脂肪酸组成及含量分析
肌肉中脂肪酸的沉积受年龄、品种、性别、饲养管理、营养调控等多种因素影响,其组成与含量不仅影响肉品的价值,还决定肉品风味,且关乎人类的膳食健康问题[63],脂肪降解形成的短链脂肪酸更是不同种属间特殊风味差异的主要物质[64]。现有的研究表明棕榈油酸(C16:1)、油酸甘油三酯(C18:1)、亚油酸(C18:2)对肉风味作用较大,而硬脂酸(C18:0)、十八碳三烯酸(C18:3)、葵酸(C10:0)、丁酸(C4:0)与肉品异味呈显著正相关[34]。本研究中3 个部位的葵酸(C10:0)、硬脂酸(C18:0)、棕榈油酸(C16:1)无显著性差异,与欧慧敏等[5](背最长肌、前肢的葵酸含量高于后肢,后肢的硬脂酸含量高于前肢,前肢的棕榈油酸含量高于后肢)、罗玉龙等[6]的研究结果(背最长肌的葵酸含量高于臂三头肌,股二头肌的硬脂酸高于臂三头肌)相比有差异。同时将3 个部位的葵酸、硬脂酸、棕榈油酸含量与现有的一些研究报道进行比较发现:本研究中的葵酸含量与罗玉龙等[6]对小尾寒羊的报道较为一致,高于刘孟君等[8]对河谷型藏绵羊的报道,低于刘璐等[43]对苏尼特羊、欧慧敏等[5]对呼伦贝尔羔羊的报道;硬脂酸含量高于欧拉羊[46]、小尾寒羊[6]、河谷型藏绵羊[8]的报道,与苏尼特羊[43]的报道接近;棕榈油酸低于欧拉羊[46]、小尾寒羊[6]、苏尼特羊[43]、河谷型藏绵羊[8]的报道,与呼伦贝尔羔羊[5]的报道接近。可见,杂交F1代的肉品风味、异味相比于河谷型藏绵羊有一定变化,但从比较的结果来看,变化并不明显,从大范围来看,在可接受的范围。本研究在3 个部位中均检测出了31 种脂肪酸,与小尾寒羊[6]、欧拉羊[46]、苏尼特羊[43]、呼伦贝尔羔羊[5]的肌肉中分别含有20、24、30、21 种脂肪酸相比,脂肪酸种类较为丰富,与河谷型藏绵羊[8]肌肉的脂肪酸种类(31 种脂肪酸)进行比较,发现脂肪酸种类是一致的,值得一提的是检测出的EPA、DHA,通常大量存在于海洋生物中[65],EPA、DHA 作为多不饱和脂肪酸的重要组成部分,近年研究表明它们对癌症具有抑制作用,富含EPA、DHA 的食物备受推崇[66]。进一步分析发现,背最长肌、臂三头肌、股二头肌的棕榈酸含量分别占总脂肪酸含量的25.55%、22.80%、23.52%,其次硬脂酸含量分别占总脂肪酸含量的22.61%、22.49%、21.63%;单不饱和脂肪酸由油酸(C18:1n9c)组成,背最长肌、臂三头肌、股二头肌的油酸含量分别占总脂肪酸含量的35.82%、34.54%、33.62%;多不饱和脂肪酸中亚油酸(C18:2n6c)的含量最多,背最长肌、臂三头肌、股二头肌的亚油酸含量分别占总脂肪酸含量的4.74%、6.62%、6.70%,表明3 个部位的肌肉均富含单不饱和脂肪酸,尤其是油酸的占比极大,Sañudo等[39]研究发现羊背最长肌肉的膻味与油酸、硬脂酸、亚麻酸等短链脂肪酸的含量呈正相关,与亚油酸的含量呈负相关,本研究所测的油酸、硬脂酸含量相比于欧拉羊[46]、苏尼特羊[43]、呼伦贝尔羔羊[5]的报道要低。就本研究而言,3 个部位的肌肉仅棕榈酸、花生四烯酸含量具有统计学上的差异,与前人得出的结论并不一致,Rouse 等[36]研究表明组织成熟具有优先性(骨组织成熟优于肌肉组织优于脂肪组织)、李晓亚等[35]研究指出脂肪酸在不同组织间的沉积具有主次性,受放牧饲养模式影响,杂交F1代在屠宰时还处于生长发育阶段,脂肪沉积并未到最佳观测点,这可能也是3 个不同部位肌肉没有差异的原因之一。
4 结论
本研究结果表明天然放牧饲养模式下18 月龄的萨福克与河谷型藏绵羊杂交F1代的背最长肌、臂三头肌、股二头肌的肉品质存在一定差异。背最长肌的肉色优于臂三头肌、股二头肌,臂三头肌的谷氨酸、锌元素、花生四烯酸含量高于背最长肌,背最长肌的组氨酸、棕榈酸含量高于臂三头肌,总体来看,杂交F1代背最长肌、臂三头肌、股二头肌均富含矿物元素、维生素、氨基酸和脂肪酸,3 个部位蛋白营养价值差异不显著,臂三头肌的鲜味氨基酸含量略高于股二头肌、背最长肌,且棕榈酸、硬脂酸是饱和脂肪酸主要组成部分,油酸是肉中单不饱和脂肪酸主要组成部分,亚油酸、α-亚麻酸、花生四烯酸是多不饱和脂肪酸主要组成部分。
猜你喜欢
临床骨科杂志(2021年6期)2022-01-08
解剖学杂志(2021年6期)2021-12-31
中国生殖健康(2020年5期)2021-01-18
中国生殖健康(2018年5期)2018-11-06
解剖学杂志(2018年2期)2018-02-13
中华骨与关节外科杂志(2017年1期)2017-05-17
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
广西林业科学(2016年2期)2016-03-20
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
中国洗涤用品工业(2015年8期)2015-02-28
