
雌穗膨大多行数玉米突变遗传性状鉴定评价
2023-11-15 10:31文仁来田树云黄爱花邹成林莫润秀翟瑞宁黄开健韦新兴谭业琴何雪银
种子 2023年8期
杨 萌, 文仁来, 田树云, 黄爱花, 邹成林, 莫润秀,翟瑞宁, 黄开健, 韦新兴, 谭业琴, 何雪银
(1.广西壮族自治区农业科学院玉米研究所, 南宁 530007;2.广西农业职业技术大学, 南宁 530007)
雌穗是玉米产量的直接载体,穗长、穗粗、穗重、穗行数、行粒数和轴粗、轴重等是重要的产量组成部分[1]。在相同种植密度下,玉米单位面积产量由雌穗数和单穗籽粒产量决定,而单穗籽粒产量由每穗粒数和百粒重决定,每穗粒数由穗行数和行粒数决定[2]。通常情况下雌穗体积越大,籽粒行数越多,单株产量越大。但热带、亚热带玉米材料相较于温带玉米材料雌穗偏小、穗行数偏少,因此鉴定、分析从广西亚热带玉米群体改良过程中收集的雌穗膨大多行数玉米材料的表型遗传特性,对进一步解析玉米雌穗行数的内在决定因素及其遗传规律,定向改良广西玉米育种群体的穗行数性状具有重要意义。
玉米生殖生长开始后,茎顶端分生组织转变成花序分生组织(Inflorescence meristem,IM),IM形成排列整齐的成对小穗分生组织(Spikelet pair meristem,SPM),1个SPM产生2个小穗分生组织(Spikelet meristem,SM),每个SM再产生2个小花分生组织(Flower meristem,FM)[3],共经历生长锥伸长、小穗分化、小花分化和性器官形成4个时期成为雌穗[4]。整个过程由多个基因共同调控,主效基因较少,微效QTL较多[5-6],包括直接调控花序分生组织生长发育的基因[7-12]和相关转录因子[13-16]、植物激素[17-19]及micro RNA[20-21]等调节因子。其中,雌穗行数(即在垂直穗轴方向上由SPM最终发育成的FM数目)是玉米产量高低的决定因素之一[22-23]。复合区间作图和玉米SNP10/50芯片等技术是挖掘基因位点的常用方法,多年来研究人员利用该方法在不同染色体上鉴定出多个穗行数相关QTL[24-26]。同时利用多份不同的自然群体材料进行表型-基因型关联分析,也是挖掘穗行数相关位点的有效方法。Liu等[27]在5个种植环境下对513份自然群体的玉米材料的雌穗性状进行最佳线性无偏预测分析(Best linear unbiased prediction,BLUP),在10条染色体上检测到17个穗行数QTL,其中24个单核苷酸多态性(Single nucleotide polymorphism,SNP)变异可解释大于5%的表型变异。Xiao等[1]利用包含14个亲本遗传背景的10个重组自交系(Recombined inbred lines,RIL)复合群体,获得控制4个穗部性状的107个QTL,其中控制穗行数的QTL多为加性或部分显性效应。Chen等[28]利用序列基因分型(Genotyping by sequence,GBS)测序分析方法绘制高密度染色体图谱得到一个重要穗行数位点qKRN5,但基因的定位区间长达4.8 Mb,其他多个穗部性状QTL最小定位区间也大于700 K。因此,精细定位并克隆更多穗行数相关QTL进行玉米雌穗性状的分子改良仍然任重道远。在调控雌穗轴发育的单基因突变中,研究较为深入的是FEA2基因,该基因编码一个受体蛋白,通过CLAVATA基因信号通路调节分生组织发育,其突变体雌穗分生组织过度增殖导致雌穗过度膨大[29]。Peter等[10]进一步研究发现,一份FEA2基因的弱等位突变型,在引起穗轴扁化和穗行数增加的同时不导致果穗过度变形,比野生型植株增产13%。Byoung等[30]报道了与FEA2表型相似的突变体FEA3,其雌穗顶端出现多个生长点,穗轴扁化,研究表明,FEA3通过响应CLAVATA3/CLE在器官原基中的表达进而调控雌穗分生组织增殖,其弱等位突变型同样能提高杂交玉米产量。
本研究对广西亚热带玉米群体进行改良的过程中收集到一份雌穗膨大多行数材料,但对于该材料的种质背景、变异特征、细胞组织特征以及受单一基因还是多个QTL位点调控等方面并不清楚,难以开发利用,因此将其与穗轴较细、穗行数12行的优良育种材料桂18421杂交再自交构建遗传分析群体,进行性状鉴定和遗传背景分析,以明确该材料表型性状的遗传分离规律,并利用40对SSR标记和23份广西育种中使用的骨干自交系,对该多行数玉米材料自交过程中保留的2个姐妹系进行遗传背景分析,为利用多行玉米材料进行玉米群体改良、新品种选育、QTL挖掘及分子标记辅助育种奠定基础。
1 材料与方法
1.1 材 料
本试验材料为广西基础育种群体(7239与CML285杂交建立的选系群体,7239为早年间玉米品种隆玉2号的母本之一,CML285来自国际玉米小麦改良中心,该群体曾选育出广西优良杂交品种桂单0810的母本桂39722)和对新材料培育中发现的雌穗膨大变异植株(经多代自交纯化形成的稳定变异材料),该变异株穗行数22行,初步命名为fcb(flat corncob),在自交纯化过程中保留了2个姐妹系,分别命名为fcb1和fcb2(图1B、C),其雌穗外形与已报道的FEA2基因弱等位型突变体具有一定相似性(图1D~G)[10]。试验对照材料为穗行数12行的广西自选材料桂18421(图1A)。

注:A为对照桂18421,B~C为fcb,D~G为FEA2突变体及等位突变表型。
1.2 方 法
1.2.1 田间种植
将fcb1(P1)与桂18421(P2)杂交,并连续自交2代,构建F2∶3群体,进行农艺表型、细胞形态和遗传分离分析,P1、P2、F1、F2和F3各世代分别调查10株、10株、10株、300株和3 000株,田间种植管理与大田农业生产相同。与fcb两个姐妹系(fcb1和fcb2)以及其他23份材料一起进行遗传背景分析(表1)。
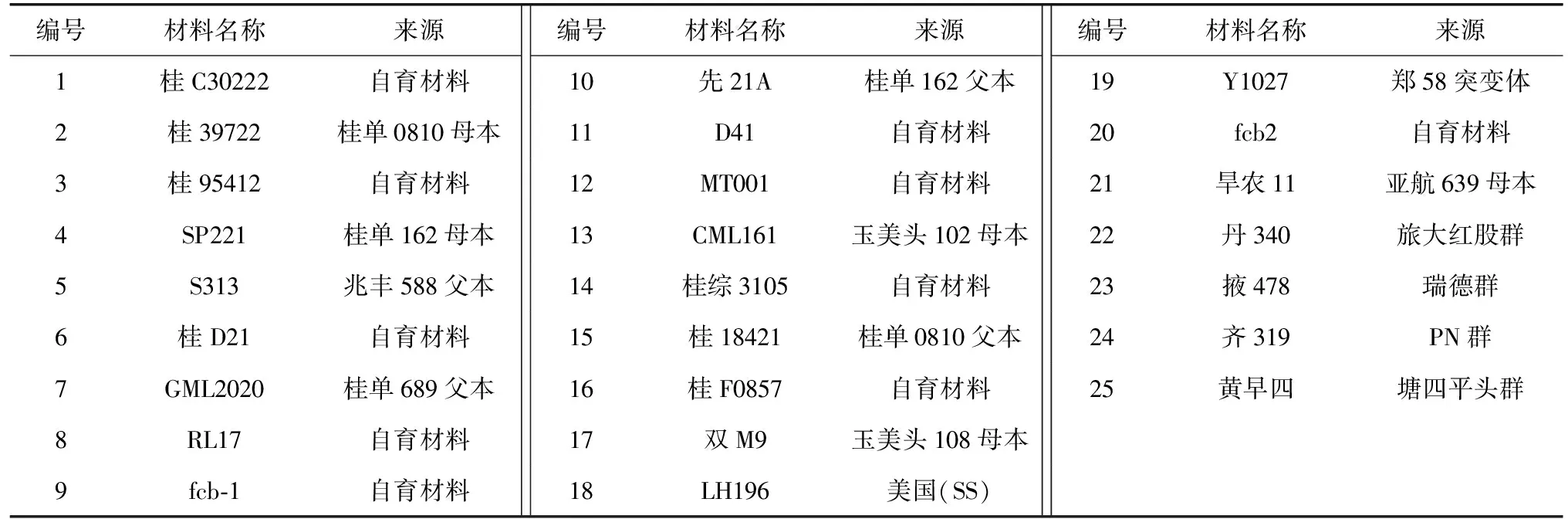
表1 用于遗传背景分析的玉米材料名称及其来源
1.2.2农艺性状测定与分析
进行表型测量的P1、P2、F1和F2各代植株全部自交授粉,待果穗收获晒干后考察穗粗、轴粗、穗行数以及穗轴是否扁化,穗粗和轴粗分别用游标卡尺测量横切面,扁穗横切面表现出椭圆形,分别测量其(穗、轴)长、短直径,并以平均值表示该果穗的穗粗或轴粗,以长轴/短轴表示穗或轴的扁化程度,比值等于1表示未发生扁化。P1、P2和F1的各项表型数据分别为10株的平均值,F2∶3家系的穗行数测定为F2单株结合相应F3后代数据,其300份F2雌穗分别单穗考种,300行F3每行测定10枚雌穗取平均值与对应上代F2进行比较,以增加F2表型数据的准确性。
1.2.3细胞组织观察
在授粉后5~10 d,取发育中的幼嫩雌穗,在雌穗中上部垂直穗轴方向横切取样,制作石蜡切片和番红固绿染色,光学显微镜观察细胞组织结构形态,主要步骤如下:
材料脱水:将新鲜雌穗组织用FAA固定液固定24 h,然后取出放于脱水盒内,再放进脱水机内依次用不同浓度梯度乙醇进行脱水:75%乙醇4 h,85%乙醇2 h,90%乙醇2 h,95%乙醇、1 h,无水乙醇2次,各30 min,醇苯1次10 min,二甲苯2次,各10 min。
石蜡包埋:65 ℃融化石蜡1 h,将浸好蜡的细胞组织置于包埋机内并贴上对应的标签,再置于-20 ℃冻台冷却,蜡凝固后将蜡块从包埋框中取出并修整蜡块。
切片:将修整好的石蜡块置于石蜡切片机切片(厚4 μm),切片漂浮于摊片机40 ℃温水上将组织展平,载玻片将组织捞起,60 ℃烘箱内烤片。
脱蜡:依次将切片放入二甲苯脱蜡2次,各20 min,无水乙醇脱蜡2次,各5 min,75%乙醇1次5 min,蒸馏水清洗。
染色:将切片放入植物番红染色液中染色2 h,自来水清洗去除多余染料,将切片依次放入50%,70%和80%梯度乙醇中各5 s,之后将切片放入植物固绿染色液中染色约15 s,无水乙醇脱水3次。
封片观察:将切片置于二甲苯中透明5 min,中性树胶封片,显微镜镜检,进行图像采集。
1.2.4遗传背景分析
采用CTAB小量提取法制备玉米基因组总DNA[31],从玉米基因组数据库(www.maizegdb.org)中查找合成SSR标记,选择其中带型清晰、分布均匀,多态性明显的40对(表2),采用10 μL PCR体系扩增,经7%聚丙烯酰胺凝胶电泳,0.1%硝酸银染色显影,以Excel2016软件记录带型统计结果,将相同迁移率位置上有条带的记1,无条带的记0。

表2 40对SSR标记
1.3 统计分析
试验数据采用Excel2016软件记录测量数据,利用SPASS19.0软件进行统计学分析;采用NTsys2.10e软件进行聚类分析。
2 结果与分析
2.1 多穗行玉米雌穗表型性状分析
雌穗膨大主要表现为穗行数增加、果穗变粗、穗轴变粗、穗扁化和轴扁化5种特征,这些特征在F2代发生不同程度的分离。如图2所示,在F2代分离群体中,F2代穗行数具有12,14,16,18,20,22行共6种分离表型,相关性分析结果(表3)显示,雌穗行数与穗粗、轴粗及穗扁化度均呈极显著性相关,与轴扁化度不相关,说明该变异材料雌穗膨大性状的不同特征可能受多个数量性状位点(QTL)分别控制,且不同位点之间有累加或协同效应。结果(表4)表明,不同雌穗行数对穗粗有极显著性影响,对轴粗存在显著性影响,同时由穗粗、轴粗与行数的相关性度量分析结果(表5)可知,随着雌穗行数的增多,穗粗增加的正向效应大于轴粗,说明其QTL增加穗行数带来的增产潜力大于其负面影响。在F2分离群体中,6种穗行数的玉米分离比为1∶9.6∶12.6∶7.9∶1.8∶0.6,呈连续正态分布,说明有2个QTL调控影响穗行数性状,基因间具有累加效应。

图2 F2群体不同穗行数频率分布情况

表3 F2群体各表型性状指标的相关分析结果
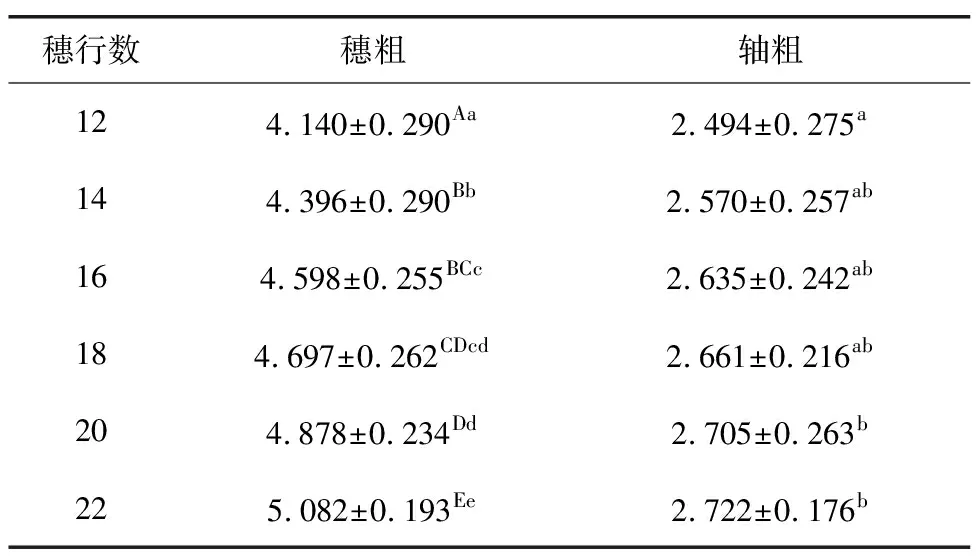
表4 F2群体不同行数对穗粗与轴粗的影响

表5 F2群体穗粗、轴粗与行数的相关度量分析结果
2.2 多穗行玉米雌穗膨大的细胞学成因
通过对幼嫩雌穗做横截面石蜡切片,经番红固绿染色后进行光学显微观察,发现穗轴变异特征表现出分生组织体积变大,髓腔及其外围维管束数目和层数变多,在正常维管束层外围和小穗着生部位出现较多散乱分布的形态完整度不一的维管组织细胞,如图3所示,该特征与FEA2弱等位突变型植株的雌穗表型变化较类似[15],主要为花序分生组织细胞过度分裂分化,穗轴各心皮在生长过程中过度发育相互挤压,导致穗轴变形。
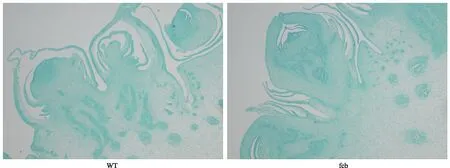
图3 fcb雌穗横切面小花形态
2.3 多穗行玉米的杂种优势群划分
利用40对SSR标记在25份样品中共检测出162个电泳带型清晰稳定、多态性良好的等位基因,每对标记可检测3~5个,平均4.1个。以遗传相似度系数0.77为标准,可将25份玉米材料划分为3个杂种优势群,少量遗传距离较远的种质材料无法有效分群。
第Ⅰ类群包含6份材料,是以泰国Swan(late)C4群体为基础选育的热带、亚热带玉米材料及其衍生种质,包含品种桂单0810的父本桂兆18421。第Ⅱ类群包含10份材料,以遗传相似度系数0.79为标准,可进一步分为2个亚群,第Ⅱ-1亚群以桂39722为核心,包含掖478,具有一定亚热带种质血缘的改良瑞德群体;第Ⅱ-2亚群包括4份广西本土自交系材料。第Ⅲ类群包含3份材料,均为广西本土自交系。此外有3份种质材料(先21A、LH196、W8555)与其他材料遗传距离较远,未能有效分群。本研究中使用的玉米材料齐319、丹340和黄早4为国内玉米杂种优势群常用测验种,聚类分析结果表明,本研究未包含与其背景较为相近的种质材料。由此可知,fcb与目前广西本土主栽的桂单0810、母本桂39722、广西国审品种桂单203、母本桂F0857、玉米新组合689、父本GML2020及掖478等划归为一类,属于具有温-热种质背景的改良瑞德群体,而其自交过程中保留的两个姐妹系在40对SSR标记中未检测到差异。
3 结论与讨论
3.1 玉米雌穗膨大材料为一份新的基因变异
在已公开报道的参与调控雌穗发育的相关基因中,2013年报道的玉米基因FEA2发生隐性突变后可导致雌穗的膨大变形,最终引起穗行数增加而促进增产[29],本研究材料与2016年报道的FEA2弱等位突变表型较相似,但又具有明显不同的特点:FEA2基因调控雌穗分生组织发育,基因突变后表型变化比较剧烈的等位型会出现分叉和不规则的雌穗,弱等位突变型会导致雌穗扁化、穗轴变粗、尖端月牙形、穗行数增加和穗行不整齐、穗中部籽粒不饱满等表型,且这些变化在后代不会发生分离[10]。本研究材料具有的此类特征会在F2代等后代群体不同单株间发生一定分离,表明其雌穗体积增大、变化及多行数等特征可能由多个QTL控制,这点与FEA2基因的突变体不同。2022年报道的控制玉米穗行数的KRN2基因来自具有野生玉米背景穗行数6行的特异玉米材料,其野生等位型负调控玉米雌穗行数,利用基因编辑技术创制的KRN2基因功能丧失材料可提高约10%的玉米产量[32]。可知,已克隆的FEA2和KRN2基因介导的性状变化与本研究材料fcb明显不同,因此fcb可能为新的未知基因位点变异,后期进一步将fcb增加穗行数的基因位点单独分离进行精细定位和基因克隆,具有较高的基础理论研究和育种应用价值。
3.2 雌穗行数与玉米产量密切相关
本研究根据广西种子管理局网站(http://www.gxseed.com.cn)公布的信息,对2005年、2014—2021年年度所有玉米品种的相关数据进行了归纳统计,显示最近十多年间审定广西玉米品种的穗行数平均增幅约7%,穗粗平均增幅约6%,千粒重平均增幅约10%,而穗长降幅约5%,行粒数降幅约9.5%,穗粒重和单位株数基本保持不变,表明玉米籽粒的容重和穗行数是近年来广西玉米品种的重要增产点,与邹成林等[33]和梁晓玲等[34]的研究结果一致。一般情况下,穗行数增加易伴随穗轴变粗和籽粒变小、千粒重降低等不利性状,统计结果表明,广西玉米品种的选育在提升穗行数的同时较好地将穗粗增幅控制在较低水平,同时提高了千粒重,而行粒数略微降低源自籽粒厚度增加,实际有助于增加籽粒品质和千粒重。
3.3 调控穗行数的QTL的开发利用
本研究中,利用40对SSR标记进行的遗传背景分析,将fcb划归为属于具有温-热混合血缘背景的改良瑞德群体。因此,一方面可以通过常规杂交育种的方式,利用fcb对同杂种优势群的优良材料及衍生品系进行穗行数性状改良,或者与其他杂种优势群的材料进行杂交组配,选育新品种[35-36];另一方面,利用分子生物学技术挖掘其与穗行数相关的基因位点,通过分子标记辅助进行单基因导入,实现对穗行数的定向改良,可以克服杂种优势群的限制。此外,可以进一步将fcb与其他控制雌穗发育的优良基因,例如玉米基因ZmKL9(能显著提高玉米粒重、穗长和穗粒数[37])、ZmEXPB15基因(调控玉米籽粒大小和粒重[38])等进行基因聚合,以更有效地在增加穗行数的同时避免穗轴变粗、容重降低、籽粒变小、千粒重减小、雌穗变短以及配合力降低等负面影响[39]。因此,在后续研究中,可利用筛选出的行数多和行数少的F2∶3家系,分别提取F2叶片DNA,构建两个极端DNA混池,在全基因组利用分子标记进行基因初步定位,并利用高世代回交群体进一步精细定位,开发与穗行数相连锁的分子标记。
综上所述,从广西玉米基础群体中收集的雌穗膨大多行数玉米材料为一份新的未知基因突变材料,具有温-热混合血缘背景,其多穗行性状符合广西玉米生产需求,可用于群体性状改良或利用温-热×热杂种优势配对模式对其进行杂交组配,选育新品种。同时,可进一步挖掘其控制穗行数的QTL,进行分子标记辅助育种及解析玉米雌穗发育的分子遗传机理。
猜你喜欢
中国农业大学学报(2022年4期)2022-05-26
中国种业(2022年4期)2022-04-19
农业工程学报(2021年14期)2021-10-12
作物学报(2021年7期)2021-04-27
读天下(2020年4期)2020-04-14
中国种业(2019年7期)2019-07-24
中国种业(2018年2期)2018-03-06
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
