
三叶木通种子形态、解剖及萌芽生物学特性的研究
2023-11-15 10:31赵小琴曹运梅段佳欣蔡小东
种子 2023年8期
赵小琴, 曹运梅, 段佳欣, 蔡小东
(1.长江大学园艺园林学院/香辛作物研究院, 湖北 荆州 434025;2.荆州佳之源生物科技有限公司, 湖北 荆州 434025)
三叶木通(Akebiatrifoliata)为木通科木通属多年生落叶木质藤本植物,因其果实于阴历八月纵向炸开,也被称为八月瓜、八月炸[1]。三叶木通分布广泛,野生资源常集中于热量丰富、降水充沛、湿度较高、日照时间相对较短的区域[2],在贵州、陕西、湖南、河南等省份人工种植已规模化[3]。三叶木通兼有药用、食用、观赏及园林应用价值,其果皮、果肉、种子具有多种用途,是一种开发前景广阔的特色经济植物[4-5]。三叶木通作为传统中草药已有2 000多年的历史,具有抗癌、抗炎、抗肿瘤和抗风湿等药用价值[6-7]。果实富含氨基酸、蛋白质、可溶性糖、有机酸和多种矿物质,口感细腻而甘甜,可作为药食兼用的新型水果进行开发[4-5]。种子脂肪酸含量较高,且以中油酸、亚油酸以及棕榈酸等不饱和脂肪酸为主,可作为新型油料作物利用[8]。此外,三叶木通茎蔓柔软多姿,花和果实具有一定的观赏价值[4-5]。
种子生物学特性及萌芽特性研究是种子繁殖及种质保存的基础[9-11]。三叶木通果实种子数量多,但种皮内含有大量油脂、天然胶,蜡质化程度较高,野生环境中种子发芽率较低,野外调查中很少发现实生小苗[12]。近年来,三叶木通已进军中国高端水果市场,野生资源产量难以满足市场需求,亟需人工大量繁殖良种苗木[13]。另一方面,人类活动导致三叶木通生长环境恶化,野生资源流失日益严重,三叶木通资源的收集、保存和管理亟待加强[1,12]。一般而言,种子保存是植物种质资源长期保存最理想的方式。根据种子的贮藏特性,可将种子分为正常型种子、顽拗性种子和中间型种子,这三种类型种子的特征差异较大,对水分敏感性、贮藏条件均不相同[4,14]。正常型种子种胚发育成熟后脱水耐性强,在一定程度上能承受脱水而不影响种子的活力和发芽率[4]。而顽拗性种子在脱水过程中,由于其脱水耐性差,导致种子发芽率不断下降,直至活力丧失[14]。
目前,对三叶木通的研究主要集中于野生资源调查、营养成分、药用成分等方面[2,6-7],而对三叶木通种子的生物学特性如种子的形态结构、形成和发育、休眠与萌芽特性等方面研究较少[15-16]。因此,本研究从三叶木通种子的形态及解剖特征、吸水率、发芽条件、脱水敏感性等方面进行系统研究,旨在为三叶木通种子繁殖及种质资源保存提供参考。
1 材料和方法
1.1 试验材料
野生三叶木通成熟果实采自贵州省镇远县三叶木通自然分布区,室内常温放置一周后从果瓤中分离种子并反复搓洗干净,于阴凉通风处晾干后立即进行试验。
1.2 试验方法
1.2.1种子基本形态特征观测
随机挑选饱满的三叶木通种子,在解剖镜下观察其形状、颜色、光泽及表皮等特征。随机取50粒新鲜种子,采用游标卡尺(精度0.01 mm)分别测量种子的长度、宽度、厚度,重复3次。
1.2.2种子千粒重的测定
参照农作物种子检验规程(GB/T 3543-1995)[17],采用百粒重法测定三叶木通种子千粒重。随机挑选100粒种子,用万分之一电子天平称其重量,重复8次后取其平均值,换算成千粒重。
1.2.3种胚长度的测量
随机取15粒新鲜种子,用手术刀将种子沿纵轴切开,用解剖针将种胚剥出,在体视显微镜下测量种胚长度,计算种胚长度/种子长度比值,重复3次。
1.2.4种胚的解剖观察
选取三叶木通外观饱满的种子,去除外种皮后固定于FAA固定液中,采用常规石蜡切片法制作样品切片[18]。切片厚度10 μm,采用甲苯胺蓝O (Toluidine blue O,TBO )染色,在ECLIPSE NI-U显微镜下观察并拍摄图片。
沥青质量直接影响混合料质量,应选用具有良好温度敏感性、抗老化性,含蜡量低的优质沥青。本文采用70#道路石油沥青,其主要技术指标如表4所示。
1.2.5种子吸水率的测定
取大小一致、种粒饱满的新鲜种子分成2份,一份种子保留完整种皮,另一份自胚根端用刀片切除少量部分,得到破皮种子。两个处理种子分别称重,在25 ℃恒温条件下于蒸馏水中浸泡1,3,5,7,9,11,24,48 h后分别取出,滤纸吸干种子表面水分后用分析天平称其重量,直至种子恒重。计算其吸水率,以此分析种皮的透水性。每个处理重复3次,每个重复30粒种子。
吸水率/%=[(浸泡后质量-浸泡前质量)/浸种前质量]×100%。
1.2.6种皮对种子萌发的影响
采用培养皿发芽方式。选取颗粒饱满、均匀一致的新鲜种子,0.1% KMnO4浸泡30 min后用无菌水冲洗3次,用滤纸吸干表面水分后分成2份,一份种皮完整,另一份进行破皮处理。分别将2种处理种子放置于预先平铺双层滤纸的无菌玻璃培养皿内,每培养皿放置15粒种子,种子均匀平放于滤纸上,加入蒸馏水使其充分湿润。种子处理好后置于人工气候箱中培养,培养条件为温度(25±1)℃,相对湿度70%,每天光照12 h。在培养过程中,每隔3 d喷洒适量无菌水以确保种子处于湿润状态,每天观察并记录种子萌发状况,连续5 d没有种子发芽则结束调查。发芽标准为胚根突破种皮并生长至种子长度的1/2以上。每个处理60粒种子,3次重复。
发芽率/%=(发芽种子数/供试种子总数)×100%。
1.2.7不同温度对种子萌发的影响
采用湿沙层积催芽方式。试验前三叶木通新鲜种子用0.1% KMnO4消毒30 min,用无菌水冲洗3次。将种子均匀平铺于高温高压灭菌后的湿沙(含水量约60%)中,之后再覆盖一层湿沙,定期喷洒适量无菌水以确保沙子处于湿润状态。层积温度分别设置4 ℃、8 ℃、16 ℃、25 ℃等4个处理,每个处理30粒种子,3次重复。中层积至第30天时分别统计各处理温度种子的发芽率。
1.2.8脱水时间对种子活力的影响
采用硅胶干燥法对三叶木通种子进行脱水处理,具体操作为:在玻璃干燥器底部放一层硅胶(厚约3 cm),将三叶木通新鲜种子平铺于硅胶上,然后再平铺一层厚约4 cm的硅胶。分别在脱水处理的0,4,8,12,24,48,96 h时取样,采用TTC法测定种子活力。具体方法如下:用手术刀沿种子纵轴切开,取其中种胚保留完整的一瓣浸入0.2% TTC溶液中,置于30 ℃生化培养箱中避光染色12 h,染色结束后用蒸馏水冲洗3次,观察胚的染色情况,统计新鲜种子活力。每处理30粒种子,重复3次。
1.3 数据处理与分析
试验数据利用Excel2010软件进行整理。吸水率、发芽率、种子活力数据进行反正弦转换后,采用SPSS23.0软件对不同处理种子的吸水率和发芽率进行双因素方差分析,脱水处理后种子活力进行邓肯多重比较检验。
2 结果与分析
2.1 三叶木通种子的形态及解剖特征
本研究中采收的成熟三叶木通果实呈长椭圆形,果皮为紫色,具有不规则深皱纹,腹缝线为白色(图1A)。果肉为白色或淡黄色,种子数较多(图1B)。种子为扁长椭圆形或扁椭圆形,外表皮为黑褐色,有光泽(图1C)。种子由种皮、种阜、胚乳、胚等4部分组成,种胚较小,有发达的白色胚乳,基部钝圆,种脐端有一个突起的白色种阜(图1D)。从三叶木通种子纵切面解剖结构可以看出,种胚由子叶、胚轴、胚芽、胚根组成,种胚直生于基部,呈棒状,已分化出2片子叶(图1E)。
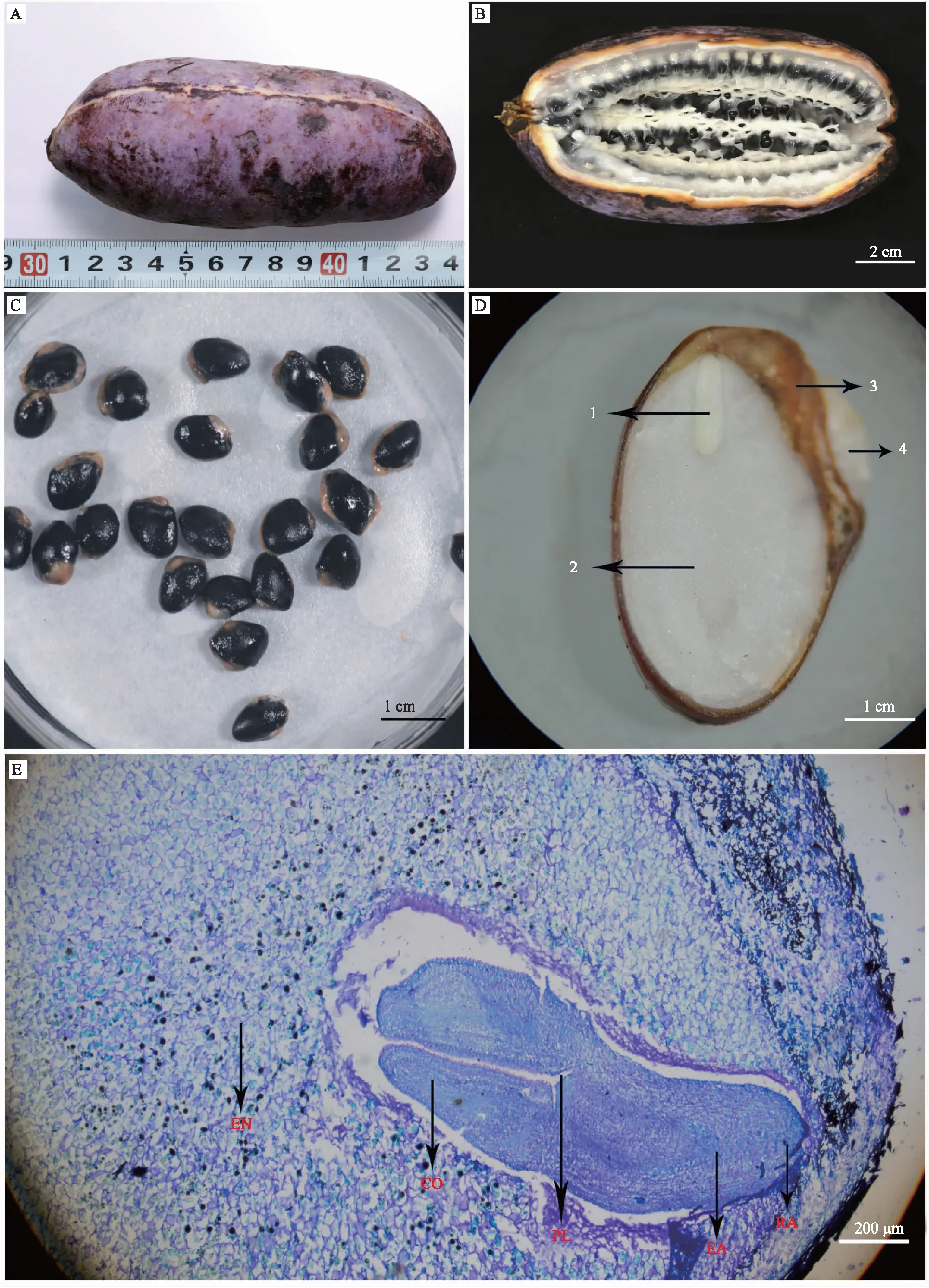
注:A、B为果实外观及纵切特征;C与D分别为种子外观及纵切结构;E为种子纵切面显微结构。1为胚;2为胚乳;3为种皮;4为种阜;EN为胚乳;CO为子叶;PL为胚芽;EA为胚轴;RA为胚根。
由表1可知,三叶木通种子平均千粒重为62.29 g,平均长度、宽度、厚度分别为8.62 mm、5.80 mm、3.21 mm,种胚平均长度为3.01 mm,平均种胚长度/种子长度为0.35,说明三叶木通种子的胚所占比例较小。这些指标的变异系数在6.79%~11.61%之间,其中种胚长度/种子长度、千粒重、种胚长度、种子厚度的变异系数均超过10%,表明三叶木通种子的形态差异较大。
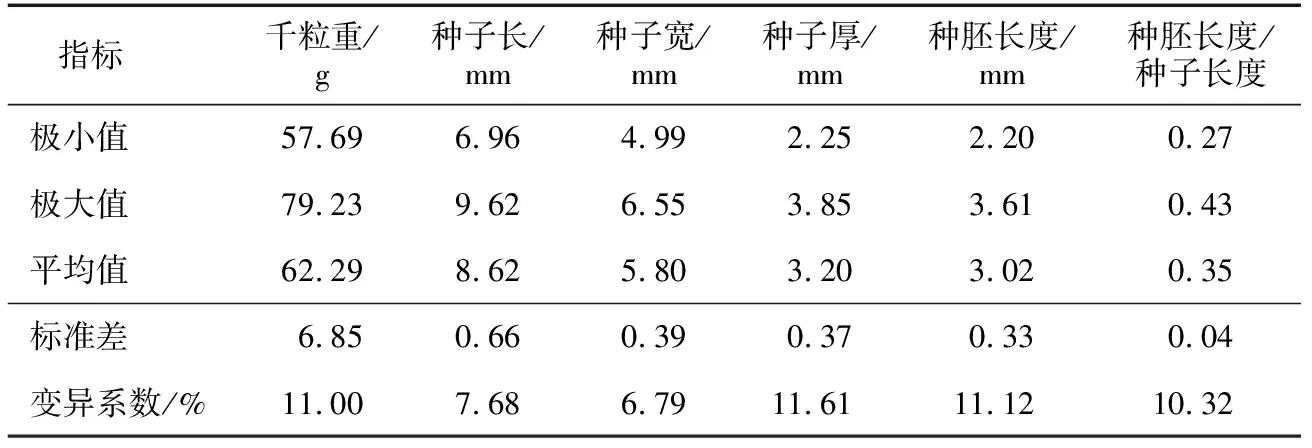
表1 三叶木通种子的基本特征
2.2 种皮对种子吸水率的影响
如图2所示,种皮完整种子与种皮破损种子的吸水趋势基本一致,均呈现先快速上升后趋于平缓的趋势。破皮处理种子浸种前3 h吸水率急剧上升,3 h后吸水率上升稍缓,11 h后种子吸水速率逐渐趋于平缓,直至达到饱和。而种皮完整种子在0~5 h内吸水率增加较快,5~11 h期间吸水率上升趋势变缓,11 h后种子吸水量逐渐变小。总体而言,在整个浸种过程中,种皮破损种子吸水率明显高于种皮完整种子,浸种后前11 h种皮破损种子及种皮完整种子的吸水量分别占总吸水量的73.21%和61.70%。方差分析结果(表2)表明,种皮完整与否及浸泡时间均对种子吸水率有极显著影响,而种皮完整与否与浸泡时间的互作对种子的吸水率没有显著影响。这些结果表明,三叶木通种子的种皮对种子吸水有极显著阻碍作用,破皮处理、延长浸泡时间有利于种子对水分的吸收。
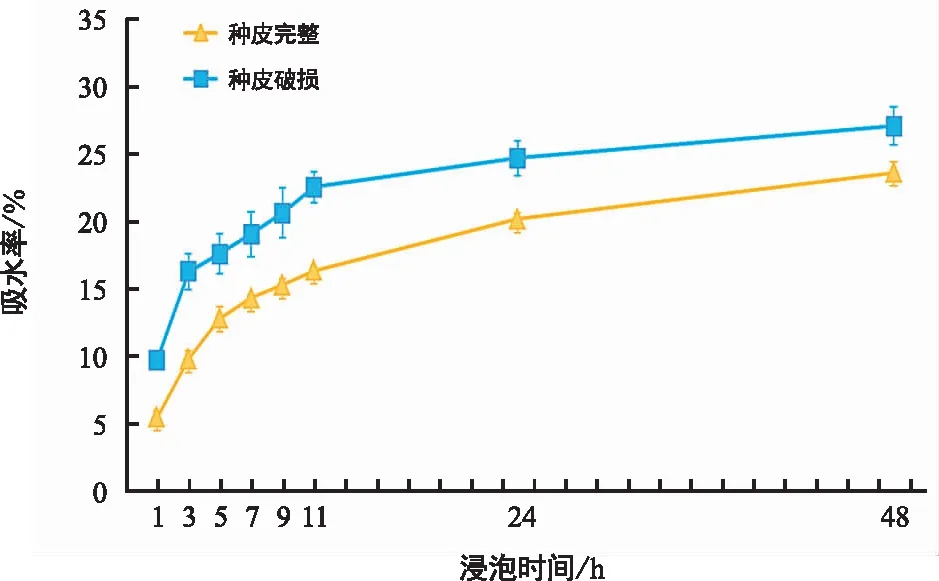
图2 三叶木通种子的吸水特性
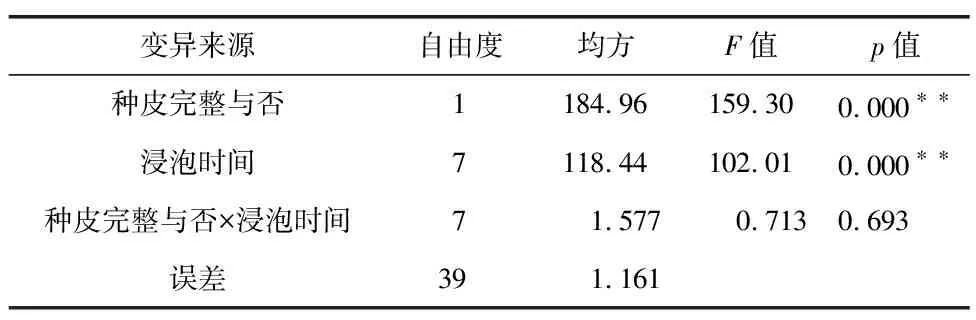
表2 种皮对三叶木通种子吸水率的影响
2.3 种皮完整与否对种子萌发的影响
种皮对三叶木通种子发芽率的影响如图3所示。当培养至第5天时,种皮破损种子中已观察到少量种子开始萌芽,发芽率达到1.97%,而种皮完整种子直至第10天时才有个别种子发芽。随着培养时间的延长,种皮破损种子的发芽率呈现明显增加的趋势,至第30天时78.97%的种子已发芽,在整个试验过程中其发芽率均远高于种皮完整种子。而种皮完整种子的发芽率在0~25 d内一直较低,直至培养至第30天时才达到17.81%。根据方差分析结果(表3)可知,种皮完整与否、培养时间及二者的互作效应均对三叶木通种子发芽率有极显著影响。这些结果表明,种皮对三叶木通种子发芽有明显的抑制作用,破皮处理并适当延长培养时间能极显著提高种子的发芽率。
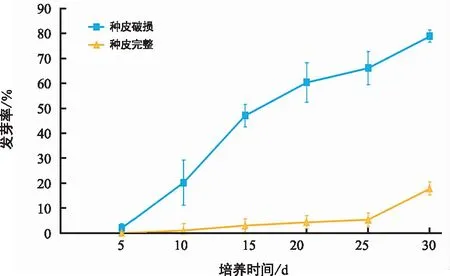
图3 种皮对三叶木通种子发芽的抑制作用
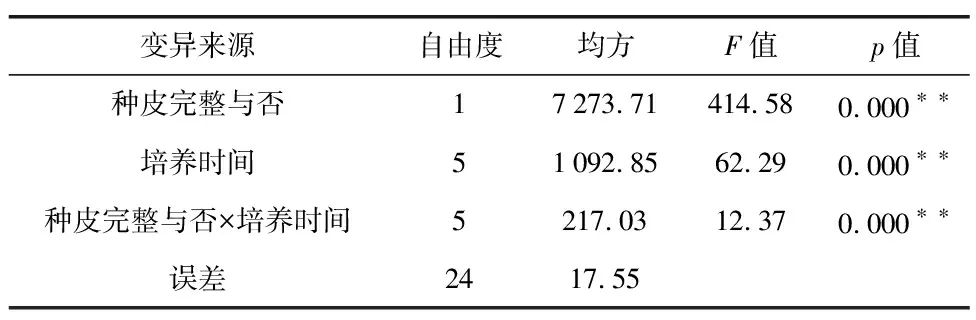
表3 种皮对三叶木通种子发芽率的影响
2.4 不同温度对种子萌芽的影响
三叶木通种子湿沙层积30 d后,温度对发芽率的影响如图4所示。4种温度处理下种子发芽率依次为:16 ℃(80.87%)>25 ℃(73.03%)>8 ℃(69.57%)>4 ℃(60.03%)。显著性分析结果显示,16 ℃层积处理种子的发芽率显著高于低温处理(4 ℃和8 ℃),但与高温层积处理(25 ℃)之间没有显著差异;25 ℃处理种子的发芽率显著高于4 ℃,但与8 ℃处理之间没有显著差异。因此,可认为16 ℃湿沙层积处理是三叶木通种子催芽的理想方式。此外,4 ℃低温处理30 d时发芽率也高达60.03%,表明三叶木通种子萌芽受低温影响较小。
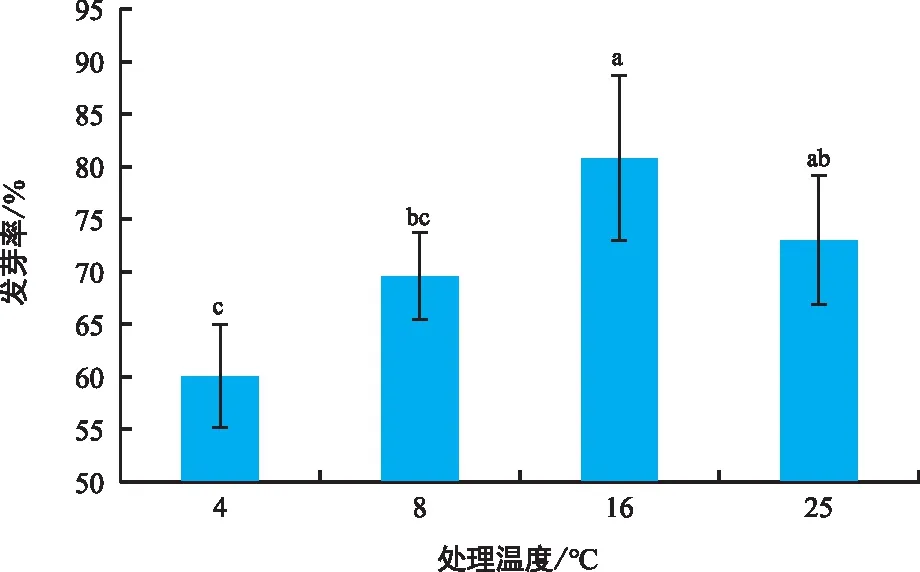
图4 不同温度下三叶木通种子的发芽率
2.5 脱水时间对种子活力的影响
不同硅胶干燥时间对种子活力的影响如图5所示。三叶木通新鲜种子经TTC染色后,其种胚和胚乳呈深红色,活力高达94.43%。随着硅胶处理时间不断延长,种子活力逐渐下降。干燥前12 h种子活力下降速度较慢,处理至第12小时时种子活力下降至76.42%,与新鲜种子相比活力下降了18.01%。硅胶干燥处理24 h时种子活力仅为46.25%,而脱水至48 h时活力降至16.90%。48 h后种子活力下降速度减缓,当种子脱水至第96小时时,种子活力仅有11.2%,此时TTC染色后的种胚已呈白色。由此可知,三叶木通种子的活力随脱水时间的延长而减少,特别是在12~48 h硅胶干燥处理间。
图5 脱水时间对三叶木通种子活力的影响
3 讨 论
研究种子生物学特性可为野生资源的繁殖、保存、引种驯化和品种改良奠定基础[9-11]。种子的表型特征主要包括大小、形状、种皮颜色、质地等,同一地区、同一品种的种子其形态结构常存在差异,很多性状可作为鉴别植物种类和品种的依据,如种子的大小、形状、颜色、种子表面的光滑度、胚和胚乳的比值等[10-11]。本研究所用三叶木通采摘于贵州省镇远县同一自然分布区,但结果发现种子的形态指标变异系数较大,尤其是种胚长度/种子长度、千粒重、种胚长度、种子厚度。熊大胜等[16]研究表明,三叶木通种子有较明显的形态后熟过程。本试验三叶木通种子来自于完全成熟且室温放置一周的果实,解剖结构显示,三叶木通种子种胚已分化出2片子叶,但无论是采用培养皿发芽方式还是湿沙层积催芽方式,种子均不能立即萌发,表明其种胚未完全发育成熟。
种子萌发受水分、温度、氧气等诸多外界生态环境影响,也与自身因素诸如种皮限制、抑制物等休眠因素有关[19-20]。三叶木通种子存在轻度种皮障碍引起的休眠现象[15-16],用茶叶或草木灰液浸种、湿沙层积等措施可促进种子发芽。通过比较破皮种子和完整种子的吸水率和萌芽率,发现三叶木通种子存在种皮障碍,从而极显著降低了种子吸水率,抑制了种子的萌发。展晓日等[21]研究发现,三叶木通种子表皮细胞下方有7~8 层厚壁细胞,种皮外层细胞的细胞壁3 面增厚且木质化,这可能是三叶木通完整种子吸水率低的解剖学原因。通过破皮处理,可显著提高三叶木通种子的吸水率和发芽率。在对叶百部[19]、酸枣[10]等其他种皮坚硬的物种研究中,破除种皮障碍后种子的发芽率也得到了显著提高。此外,种子休眠与萌芽特性因物种而异,层积处理是解除种子休眠、促进种子萌发的常见方法之一[9,20,22]。层积处理可软化种皮,提高种皮透性,促进种胚后熟过程,有效降低种子中抑制物质的含量[20,22]。湿沙层积30 d后,三叶木通完整种子的发芽率明显高于培养皿发芽方式。一些研究[9,20]表明,层积温度对种子的发芽率有明显的影响。本研究发现,湿沙层积温度对三叶木通种子发芽率有显著的影响,但三叶木通种子萌发对温度适应性较广,即使在4 ℃和8 ℃低温层积处理下也有较高的发芽率。
脱水耐性差是顽拗性种子最重要的特征之一[4]。一些植物如苏铁[14]的种子在脱水过程中,由于脱水敏感性问题,导致种子发芽率随种子含水量下降而不断降低,直至活力丧失。随着硅胶干燥脱水时间的延长,本研究同样发现三叶木通种子逐渐失去活力,脱水至第96 h时种子活力仅有11.2%。这表明三叶木通不能忍受水分的缺失,属于典型的脱水敏感性种子。建立种子库是野生植物种质资源长期保存最理想的方式之一[11]。然而,对于脱水敏感性种子而言,种子含水量的降低会导致种子发芽率的下降,甚至活力完全丧失,传统的低温种子库无法长期保存[4]。超低温保存技术目前被认为可能是脱水敏感性种子和顽拗性种子实现长期保存的唯一方式,但仍处于试验阶段[4,23]。脱水敏感性与种子结构、蛋白质含量、糖类含量、膜脂过氧化、各种酶活性、脱落酸含量以及基因表达等密切相关[23],将来应加强三叶木通种子脱水敏感性机理的研究,以为三叶木通种质资源保存奠定基础。
猜你喜欢
植物资源与环境学报(2022年6期)2022-12-16
今日农业(2022年6期)2022-07-05
潍坊学院学报(2020年6期)2020-11-22
科学导报(2020年8期)2020-03-12
中国食品学报(2020年2期)2020-01-17
许昌学院学报(2018年8期)2018-09-05
基层中医药(2018年2期)2018-05-31
潍坊学院学报(2016年2期)2016-12-01
中国民族民间医药·下半月(2014年5期)2014-12-02
云南中医学院学报(2014年3期)2014-07-31
