
濒危植物春剑的胚胎发育及果实和种子特征研究
2023-11-13 09:23王仁睿
植物研究 2023年6期
王仁睿 刘 鑫 李 杰
(西南科技大学生命科学与工程学院,绵阳 621010)
春剑(Cymbidium goeringiivar.longibracteatum)是春兰变种[Cymbidium goeringiivar.longibracteatum(Y.S.Wu et S.C.Chen)],国兰代表种之一,有正宗“川兰”之称[1]。春剑为国家重点二级保护野生植物[2],被列入《华盛顿公约》(CITES)附录Ⅱ和《世界自然保护联盟濒危物种红色名录》[3-4]。有性生殖是植物生长发育的重要过程,胚珠和种子发育是其中的关键环节。通常兰科植物授粉一段时间之后受精才发生[5],正常的胚胎发育是种子发育和萌发的基本条件。研究表明,兰科植物的种子细小,无胚乳形成,胚柄形态因种而异[5],具有分类学意义。种子萌发是影响春剑自然繁殖的生物学瓶颈问题[6],其萌发机理和条件尚不明确。目前关于春剑胚胎发育的相关研究尚鲜见报道。本研究通过体式显微镜、石蜡切片和扫描电镜3种方法观察春剑的果实和种子形态特征、胚珠发育以及胚胎发育,了解胚胎发育特征,分析可能影响其种子萌发的胚胎学因素和胚胎败育情况,是春剑繁殖生物学研究的重要内容和再引入的基础。
1 材料与方法
1.1 试验材料
春剑种植在绵阳西南科技大学兰科植物资源圃(31.5°N,104.7°E)。通过人工授粉进行自交,采集不同发育时期的子房、幼果和成熟果实,固定于FAA 溶液后,制备石蜡切片,并在显微镜下观察发育特征。
1.2 果实形态结构观察
用解剖刀将春剑自然成熟未开裂的蒴果切开,沿横切面和纵切面,在体视显微镜下观察果实内部结构,包括胎座、种子着生情况,结合差减法和粘取计数法估算蒴果中种子的数量和单粒质量[7]。称量待测蒴果的质量,记作M1,将每个果实内的种子全部取出,称量空蒴果的质量,记作M2,蒴果中种子的总质量为M1-M2。取一定量的种子称量质量,记为m1,从m1中取出部分种子,再将剩余的种子称质量,记作m2,取出的种子用粘取法计数,种子数量记作n,则种子单粒质量m=(m1-m2)/n。试验取5 个蒴果,5 个重复,称量计算,取平均值。蒴果中种子的含量(N)可根据N=(M1-M2)/m计算得出。
1.3 胚胎发育过程及胚柄细胞观察
分别取人工自交授粉后6 个不同发育阶段的果实,包括:60、80、100、120、150、180 d,去除外果皮,将带有胚珠和种子的材料,横切成5 mm×5 mm的小块,参照李和平[8]的常规石蜡制片方法,将材料至于FAA 固定液(40%甲醛5 mL+冰醋酸5 mL+70%乙醇90 mL)固定24 h 后,依次加入不同体积分数的乙醇脱水,各处理2 h,100%乙醇和二甲苯混合溶液(体积比1∶1)处理2 h,纯二甲苯处理2 h。将材料放入液态石蜡中,放置于恒温箱(博准BZ-300BSH,上海)中45 ℃连续浸蜡24 h。将处理好的石蜡和材料一起倒入包埋底膜中,盖上包埋框,将模具迅速放入冷水中,使其冷却凝固。用徕卡轮轴式石蜡切片机(Leica RM2235,德国)把包埋好的材料,切成8 μm 的薄片。将连续的蜡带放置于40 ℃热水中平展,贴在载玻片上,再将载玻片放入恒温烘箱(澳德玛ZKGT-6053,深圳)40 ℃烘片24 h,脱蜡及染色,用中性树脂封片,将切片置于徕卡光学显微镜(Leica DM750,德国)观察大孢子的发生过程。
1.4 种子形态观察
春剑成熟果实自然风干,开裂后,取部分种子直接固定在双面导电胶上,贴于样品台上,放入离子溅射仪(Quorum SC7620,英国)喷金,将样品置于蔡司扫描电子显微镜(Zeiss EVO 18,德国),观察种子表面超微结构特征。
选取5个自然开裂的成熟蒴果,每个蒴果随机选取100 粒种子放置于载玻片上,加一滴清水,置于体视显微镜下(Nikon SM645S,日本)下观察种子形态大小,用IMAGE3000 显微镜图像分析软件系统测量种子及种胚的长宽,种子和种胚体积的计算参照Arditti等[9]的方法。
式中:r为1/2 种子宽;h为1/2 种子长;a为1/2种胚宽;b为1/2种胚长。
本文的数据是平均值±标准误。
2 结果与分析
2.1 果实结构及种子含量
春剑成熟蒴果的横切面轮廓为1 个近似三角的六边形,子房由6 枚心皮组成,侧膜胎座,构成1 室子房(图1A)。其中,3 枚心皮具胎座,着生胚珠并发育为种子,为形态较大的可育心皮;另有3枚胎座缺乏胎座,为形态较小的不育心皮。成熟果实外壁呈六棱状突起,将果实分为6 瓣,每个突起为1瓣,其中3瓣体积较大,向内形成“V”字形的胎座,胎座向果实内部分化出多个分支,种子着生在胎座及其分支上。另外3瓣体积较小,无向内突起的胎座,无种子着生(图1B~E)。春剑果实有3 层果皮结构,分为外果皮、中果皮和内果皮。外果皮较薄,表面光滑无毛,且呈木栓化,较坚硬;中果皮较厚,细胞呈海绵状排列较疏松,颜色由外到内逐渐变浅,靠近内果皮部分颜色最浅,为淡绿色;内果皮为透明薄层,将种子包裹在果腔内。蒴果纵向切开后,种子自然洒落,颜色呈白色,细小如尘,肉眼难以观察单粒种子形态,种子数量众多充满整个果腔(图1F)。

图1 春剑蒴果的解剖结构A.成熟蒴果横切;B.成熟蒴果横切结构简图(P.胎座;FV.可育瓣;SV.不育瓣);C.可育瓣及“V”字形胎座;D.胎座分支;E.蒴果纵切;F.蒴果中种子生长情况Fig.1 Anatomy of the capsule of C. goeringii var. longibracteatum A.Cross section of the capsule;B.A diagram of the cross section of the capsule(P.Placentation;FV.Fertile valve;SV.Sterile valve);C.fertile valves and a‘ V’ shaped placentation;D.Placentation branchs;E.Longitudinal section of the capsule;F.Seeds growth in the capsule
单个蒴果质量平均为6.576 g,其内种子质量平均为1.476 g。根据表2 利用差减法得到的数据可知,春剑种子单粒质量为(3.92±0.40) μg,单个春剑蒴果中种子含量约(37.98±3.71)万粒。
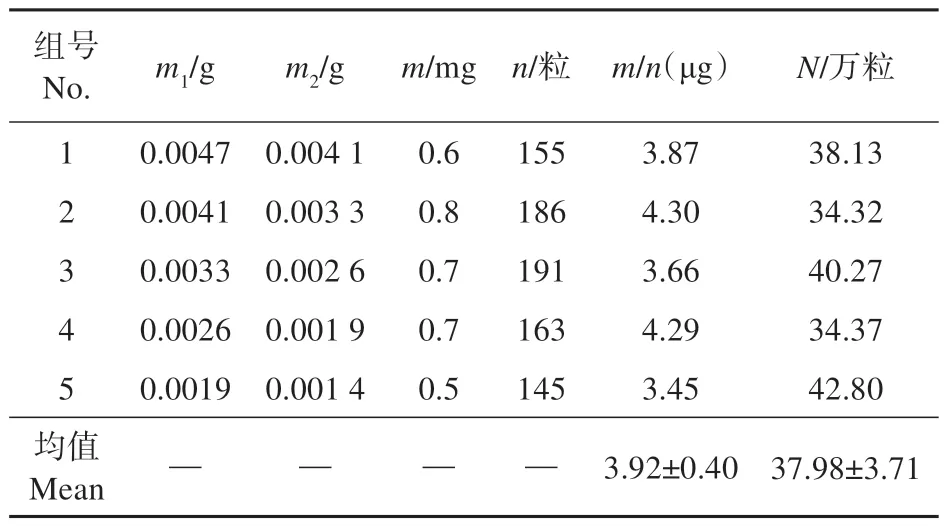
表1 春剑蒴果中种子数量Table 1 The number of seeds in capsules of C. goeringii var. longibracteatum
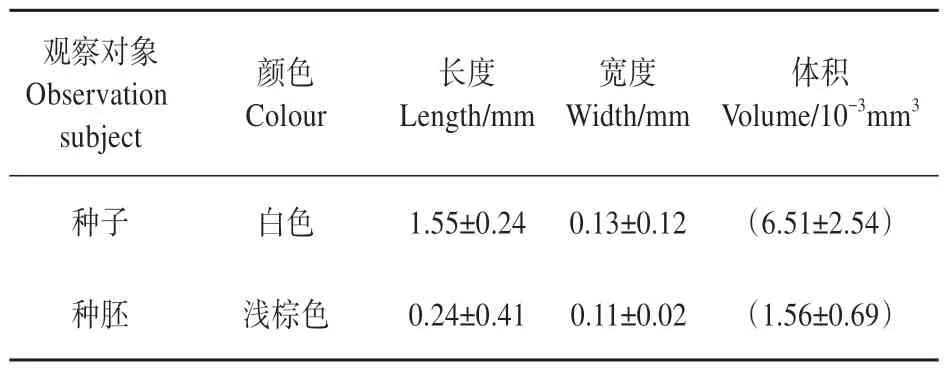
表2 春剑种子和种胚特征Table 2 The characteristics of seed and seed embryo of C. goeringii var. longibracteatum
2.2 种子形态
春剑种子为白色,长约1.55 mm,宽约0.13 mm,长宽之比约为13.4,种子呈梭形,种子体积约6.51×10-3mm3。种胚为浅棕色,长约0.24 mm,宽约0.11 mm,种胚长宽之比约为2.3,种胚呈椭球形,种胚体积约为1.56×10-3mm3(表2)。春剑种子微小,种胚在种子中体积占比较小,约占24%,种胚能储藏的营养物质较少。
正常发育的春剑种子中间大、两头稍尖,胚柄端种皮有开口,种子表面形成网状纹路(图2A~B)。种子结构简单,由内种皮、外种皮和未分化的球形种胚组成。内种皮紧密包裹着胚体生长,内外种皮之间存在明显的空气腔,空气腔占种子体积的比例约为75.61%。外种皮细胞为长方形的薄壁细胞,细胞表面不光滑,有斜条状纹路,细胞壁加厚突起,种皮呈白色透明状,种皮细胞里无内容物,透过种皮可观察种胚形态(图2C~D)。
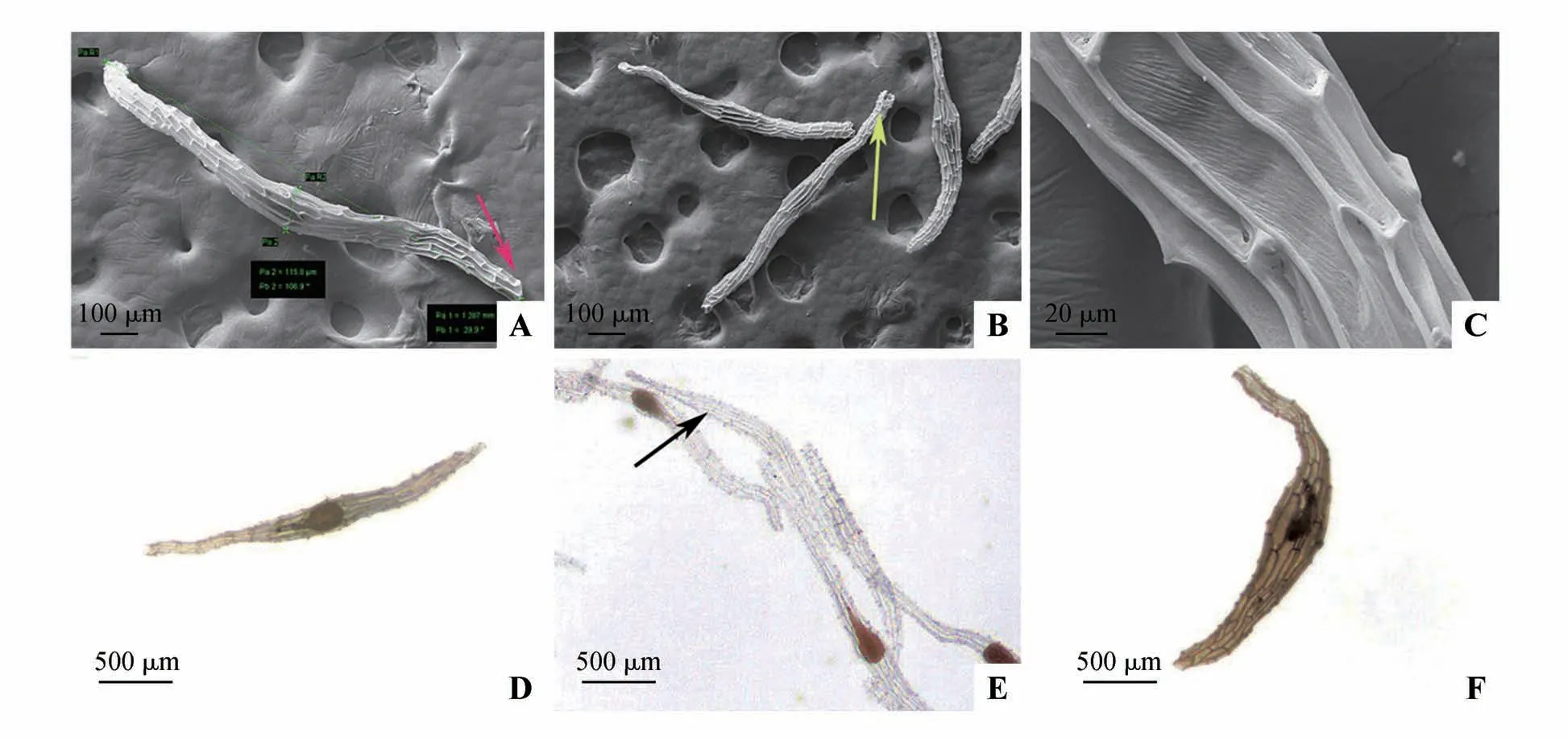
图2 春剑的种子形态和表皮超微特征A~B.电镜扫描下的种子形态,箭头所示种孔;C.种皮细胞加厚的细胞壁以及表面纹饰;D~F.体视显微镜下正常发育的种子形态(D.含胚胎的正常种子;E.有胚胎和无胚胎(箭头所示)的种子;F.畸形的种子)Fig.2 Seed morphology and epidermal ultrastructure of C. goeringii var. longibracteatum A-B.Ultrastructure of seed under SEM,arrow showed the micropyle;C.Epidermal cells with ridiged on seed coat;D-F.Seed morphology under microscope(D.Normal seeds with embryos;E.Seeds with and without embryo(arrows);F.The seed with abnormal embryo)
春剑种子发育过程中有少量的败育情况,其败育情况可分2 种:一是无种胚结构,种子仅有1 层透明的种皮结构(图2E);二是种胚发育畸形,正常发育的种胚成熟后形态呈球形或椭球形,畸形发育的种胚仅有1个深色的不规则细胞团,其体积明显小于正常种胚体积(图2F)。
2.3 大孢子发生
春剑种子在具有胎座结构的可育心皮瓣膜的胎座上发育形成,授粉后胎座细胞进行分裂和分化,向子房内部形成数十个分支结构,在胎座和分支表面分化出大量指状突起的胚珠(图3A),受精后,发育成种子。授粉后60 d 观察到胚珠内的胚囊腔出现大孢子母细胞(图3B),80 d 后可见成熟蓼型胚囊结构(图3C)。在大孢子发生过程中,胚珠内部的胚囊腔里,珠心表皮下的1个细胞分化为孢原细胞,孢原细胞再分裂形成珠心细胞和大孢子母细胞,内外珠被(图3B~C)将大孢子母细胞包裹在中间(图3B)。大孢子母细胞经多次减数分裂后发育形成成熟胚囊结构(图3C)。
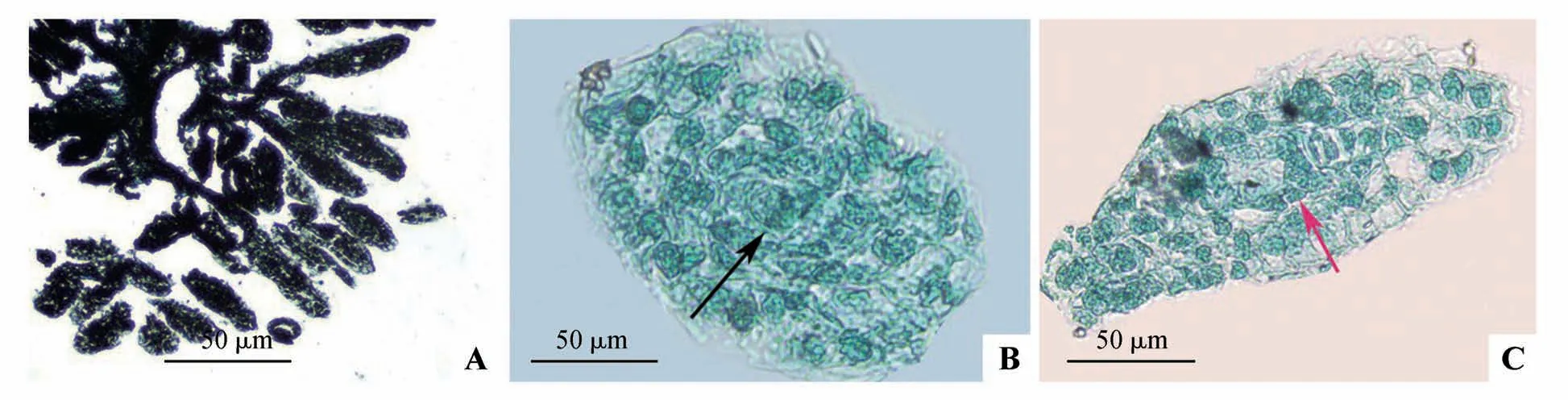
图3 大孢子发生A.胎座表面形成的胚珠原基;B.大孢子母细胞(箭头所示);C.成熟蓼型的胚囊(箭头所示为卵细胞)Fig.3 Stage of megasporogenesis A.Ovule primordium on placentation surface;B.Megasporocyte(arrow);C.Mature embryo sac of polygonum type(egg cell showed by arrow)
2.4 胚胎发育
授粉80 d 后,受精产生合子,合子细胞体积较大,细胞质较浓(图4A)。授粉100 d 后,观察到大量合子,胚胎发育已经开始,合子体积增大,呈现极性,细胞核向合点端移动,合子中部向内凹陷(图4B),随后开始第1 次有丝分裂,形成2 个子细胞,此时为双细胞原胚阶段(图4C)。春剑胚胎发育合子第1次有丝分裂形式为横裂,分别产生顶细胞和基细胞,两种细胞形态各异,且后期发育途径不同。顶细胞靠近合点端,体积较小,基细胞靠近珠孔端,体积较大,呈液泡化,基细胞和顶细胞分别发育形成胚柄和胚体。合子第2次有丝分裂,顶细胞先分裂,横裂形成1 个直线型的三细胞原胚,随后经多次分裂,胚体细胞数量不断增加,种胚体积变大。
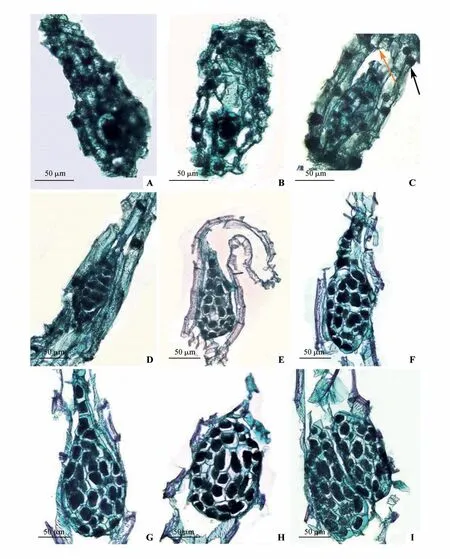
图4 种子胚胎发育过程A.受精后形成合子;B.合子第1次有丝分裂;C.双细胞原胚时期,橙色箭头所示为内珠被,黑色箭头所示为外珠被;D.早期原胚阶段;E.胚柄开始发育;F.胚柄含有5个细胞,呈线性排列;G.胚柄细胞开始退化;H.成熟种子,胚柄退化消失;I.双胚结构Fig.4 The process of seed embryo development A.Formation of zygote;B.The first mitosis of zygote;C.Two-celled proembryonic stage,orange arrow showed the inner integument,and the black arrow showed the outer integument;D.Early proembryonic stage;E.Development of suspensor;F.The suspensor contained five cells in a line;G.Degeneration of the suspensor;H.Mature seed without suspensor;I.Double embryos
授粉120 d 后,早期原胚发育形成,靠近珠孔端位置,具有1 个体积较大的胚柄细胞(图4D)。胚体体积逐渐增大,充满胚囊腔,逐渐发育形成球形胚,胚柄细胞继续生长发育,横裂形成具有2 个细胞的胚柄结构(图4E)。授粉150 d 后胚体仍停留在球形胚结构,胚柄高度发育,胚柄结构包含5 个细胞,呈直线排列向珠孔端延伸(图4F)。授粉180 d 后,胚柄开始退化,胚柄的细胞数量逐渐减少(图4G),胚体细胞不再进行分裂和分化,开始营养物质的储存,直到种子成熟,胚体依旧处于球形胚阶段。种子成熟后其种胚结构为5~7 个细胞宽,7~9 个细胞长,胚柄退化消失(图4H)。春剑种子胚胎发育过程中存在极少数的双胚结构,即1个种皮包裹着2个种胚(图4I),种胚之间独立发育生长,互不影响,且2个种胚的发育程度基本一致。
2.5 内外种皮发育
春剑种子为双层种皮结构,内珠被和外珠被分别发育形成内种皮和外种皮。石蜡切片观察发现内珠被由单层细胞组成,细胞排列整齐,细胞质较浓,细胞核体积较小(图4C)。外珠被由2 层呈长方形状的薄壁细胞组成,细胞液泡化程度较高,细胞核较大(图4C)。在胚胎发育过程中内外珠被细胞均不再进行分裂,授粉120 d 后内珠被细胞逐渐失水,形成薄而致密的一层膜状包裹在种胚之外,之后发育成紧贴胚体的内种皮,此时外珠被细胞的双层结构变为单层,靠近种胚的一层细胞退化(图4D)。授粉150 d 后外珠被细胞失水收缩,细胞器退化消失,细胞壁向外加厚,最后发育形成透明且有加厚环纹的外种皮(图4E)。此外在授粉100 d 后内外珠被之间开始形成空气腔,直到种子发育成熟,内外种皮之间可观察到明显的空气腔(图4H)。
3 讨论
春剑果实解剖结构与大多数兰科植物类似,果实分为6 瓣。3 瓣体积较大,且带有“V”字形胎座为可育瓣,种子在胎座及其分支上发育形成,另外3瓣体积较小,无胎座形成,不产生种子,为不育瓣。Johansen研究认为,兰科植物的花瓣和萼片与果实的结构中的可育瓣与不育瓣的起源相关[10]。关于兰科植物果实的结构一直以来都有争议,部分学者认为兰科植物果实是由6 心皮组成,其中3个心皮带胎座,3个心皮不带胎座;另一部分学者认为其果实由3心皮组成,分成6个瓣膜,3瓣为有胎座的可育瓣,3 瓣为无胎座的不育瓣[11],通过本研究的观察,春剑果实为3 心皮结构,2 个心皮的边缘部分组形成了“V”字形胎座。
本研究发现种子在胎座上形成的初期与胎座或胎座分支相连接,能从胎座获取营养物质,当发育到一定程度时种子将脱离胎座及其分支,随着果实成熟,种子不断形成,充满果腔。兰科植物种子数量巨大,通常几万到几十万粒不等,根据估算,1个春剑蒴果中约有37.98万粒种子,种子极轻极小,每粒仅约3.92 μg,种子缺乏贮藏的营养物质。本研究认为在种子形成过程中存在先后顺序,同一时期先形成的种子成熟度相对较高,并被后形成的种子挤向果腔中部,因此位于果实中部的种子成熟度略大于边缘部分,具体结论还需通过对不同部位种子进行活力测定来验证。
不同种类的植物从传粉到受精之间的间隔期存在着差异,通常在10~48 h 内完成受精[7]。本研究中春剑传粉到受精间隔约80 d,较多数植物间隔时间长。授粉至受精的间隔期内,子房内部的发育情况尚不明确,促进胚珠发育的物质及其作用机理,需要进一步研究。
根据被子植物胚胎发育类型的划分[12],春剑的胚胎发育类型属于茄型。春剑授粉100 d 之后,果实纵横径的变化趋于稳定,根据胚胎发育过程观察,授粉100 d 后多数种子胚胎发育开始,因此本研究认为胚胎发育过程不影响果实形态变化,营养物质全部用于胚胎发育及养分贮藏。
由于胚柄结构在形态上的差异,导致了兰科植物胚胎发育的多样性。Swamy[13]对兰科植物的胚柄结构进行了形态分类,分别为单细胞的管状或圆锥状、丝状、葡萄状、胚柄细胞包裹胚体和胚柄肥大5种类型。一些兰科植物胚柄不发达,仅有单个细胞的胚柄或是无胚柄结构[14]。而春剑的胚柄属于较发达的丝状胚柄,由5 个细胞组成,形成吸器结构。胚柄对胚体发育早期营养物质的吸收起重要作用,但胚柄是在胚胎发育中短暂存在的结构,发育到一定程度时发生程序性死亡,春剑种子成熟时无胚柄结构。
春剑胚胎发育进程存在不同步性,同时在授粉后80 d 和100 d 均能观察到合子形成;授粉120 d后,部分种子已形成2~3个细胞的胚柄结构,另一部分胚柄刚开始分化;授粉180 d 后,部分种子胚柄已经退化消失,仅存一个未分化的种胚,部分种子仍保持3~4 个细胞的胚柄结构。由果实解剖结构可知,春剑种子起源于胎座,春剑蒴果中种子含量巨大,而胎座上种子的分化有先后顺序,先分化形成的种子优先发育,后分化形成的种子发育相对滞后,因此能在同一时期观察到不同发育状态的种子。
不同兰科植物的种皮发育存在差异,部分兰科植物在胚胎发育过程中内珠被退化,种子成熟时,仅有1 层种皮结构,如兜兰(Paphiopedilum godefroyae)[15]。研究发现春剑种子种皮结构与杓兰属(Cypripedium)相似,在胚胎发育过程中内珠被一直存在,种子成熟时种皮有内外2 层结构,内外珠被分别发育形成内种皮和外种皮[16]。春剑授粉120 d 后内外种皮形态发育基本完成,内种皮薄而致密,呈膜状紧贴胚体;外种皮脱水收缩成透明的薄膜状,种子表面由于外种皮细胞壁加厚而形成网状纹路。兰科植物种子微小,自然状态下随水或风四处传播,其双层种皮的特殊结构,使兰科植物种子在严苛的自然环境下能长期保持活力[17],种子致密的内种皮结构影响了胚体对水分和养分的吸收,致使其在自然状态下萌发困难[16]。因此,春剑种子的双层种皮结构可能是导致其难以萌发的原因之一。
春剑种子微小,形态与蒴果相似,呈梭形,种子结构简单,由种皮和未分化的种胚组成,且内外种皮之间有明显空气腔,与多数兰花种子形态[9]相似。电镜扫描观察到种子的外种皮细胞之间排列紧密,没有间隙,该结构可能导致种子的不透水性和影响外界养分进入种子,阻碍萌发。此外,春剑种子的种胚体积微小,所能贮藏的养分含量极少,难以满足种子萌发过程中系列生理变化的养分需求,因此,自然状态下春剑种子的萌发率较低。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
甘肃农业大学学报(2021年4期)2021-09-22
中国生殖健康(2020年5期)2021-01-18
扬子江诗刊(2018年2期)2018-11-13
中国生殖健康(2018年5期)2018-11-06
扬子江(2018年2期)2018-03-24
福建中医药(2017年6期)2018-01-09
西南农业学报(2016年4期)2016-05-17
新疆医科大学学报(2015年10期)2015-12-26
中国粮油学报(2015年5期)2015-02-06
