
林龄对毛白杨功能性状和林下土壤理化性质的影响及其耦合关系
2023-11-13 09:27杨雨薇何宝辉韩学娇时海香张贵民路兴慧
植物研究 2023年6期
杨雨薇 何宝辉 韩学娇 时海香 张贵民 路兴慧*
(1.聊城大学农学与农业工程学院,聊城 252000; 2.聊城市林业发展中心,聊城 252000)
黄泛平原是由黄河多次冲积形成的平原,受季节性风沙危害严重,原生植被稀少,现存植被多为人工构建的防护林[1]。黄河流域生态保护和高质量发展战略倡导生态恢复与可持续发展,在此背景下,提升森林植被质量能够更好地恢复该区域的生态环境[2-3]。植物功能性状是指对植物体定植、存活、生长和死亡存在潜在显著影响的一系列植物属性[4],植物功能性状会随时间推移而发生变化,因此研究植物年龄对性状的影响具有重要意义,大量的研究显示,林龄的变化会影响植物自身、生境和群落组成[5-6]。当前的研究表明,叶片和枝条的功能性状会随林龄与环境的变化而改变,Henn 等[7]的研究发现,木质密度、木材产量或许是指征木本植物随林龄的变化主要指标。Song 等[8]表明,植物在不利生境中,植物功能性状随林龄的变化并不显著,这是植物随林龄增长逐渐适应环境使自身性状更加稳定的结果。目前,关于林龄与植物氮磷元素的关系成为了研究热点[9],靖磊等[10]通过对不同林龄杨树人工林的研究,发现叶的碳、氮、磷质量分数与林龄显著相关,且存在明显的时间变异性。Zhang 等[11]在研究中也表明,不同的养分元素随时间序列表现出不同的变化,而磷的变化最为显著。前人的研究表明,研究林龄与植物功能性状的关系能为更好地经营人工林提供理论支持。
毛白杨(Populus tomentosa)是一种速生、高产的树种,具有良好的耐干旱与耐盐碱能力,在我国华北平原种植广泛[10]。鲁西黄泛平原土地沙化严重,为减少风沙危害,大力构建防护林体系[1]。杨树作为主要树种,在该区域人工林中种植面积广泛。近些年关于黄泛平原的研究主要集中在土壤理化性质、林下植被方面[1],而对人工林植物功能性状与环境关系的研究较少。
本研究以鲁西黄泛平原区域内10、20、35年生毛白杨人工林为研究对象,在测定叶片功能性状与环境因子的基础上,分析毛白杨人工林功能性状随林龄的变化规律,阐明影响毛白杨人工林功能性状随林龄变化的主要指标,辨析毛白杨人工林功能性状与环境因子之间的关系,旨在揭示鲁西黄泛平原毛白杨对环境的适应策略,为人工林的经营与管理提供理论依据。
1 研究区域和调查方法
1.1 研究区域概况
在山东省聊城市段鲁西黄泛平原冠县毛白杨林场和高唐旧城林场(35°47′~37°2′N,115°16′~116°32′E)开展研究。研究区属于暖温带季风气候区,半干燥大陆性气候。年均气温13.2 ℃,年平均降水578.4 mm,主要集中在夏季,全年光照时间为2 463~2 741 h,无霜期200 d 左右。鲁西黄泛平原由于长期受季节性风沙危害,导致土地沙化严重,大多数森林资源为人工栽植的防护林,常见木本乔木有毛白杨、槐(Sophora japonica)和柳(Salix babylonica)等。
1.2 实地调查和样地描述
于2022 年7—8 月,开展毛白杨人工林样地调查工作。在实地踏查的基础上,于冠县毛白杨林场和高唐旧城林场选择立地条件相近、林龄分别为10、20、35 年生的毛白杨人工林作为研究对象。建立20 m×20 m 样方进行调查,每个样方之间设置10 m 的缓冲区。共建立样方38 个,其中,10 年林龄的样地12 个,高唐与冠县各6 个;20 年林龄的样地12 个,高唐与冠县各6 个;35 年林龄的样地14 个,高唐8 个,冠县6 个。对样地内胸径10 cm 以上的所有植株进行刷漆、挂牌及定位,并对其物种名、数量、胸径(DBH)及树高进行记录(见表1)。
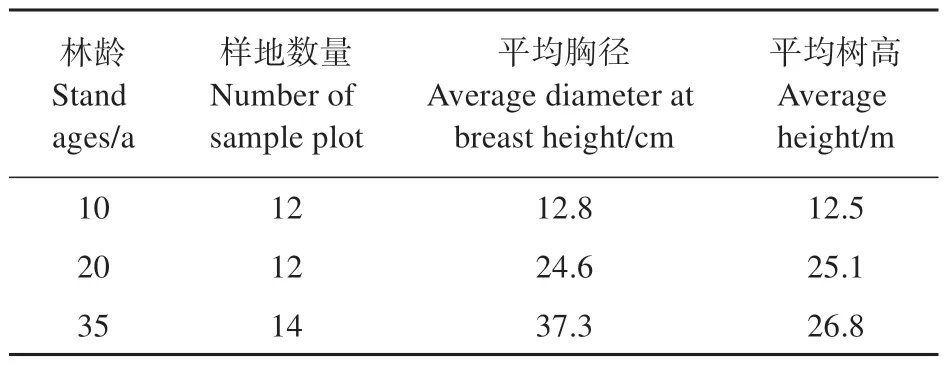
表1 样地基本情况Table 1 Basic situation of the sample plot
1.3 植物功能性状的取样和测定
对样地中胸径1 cm 以上的植株进行采样,使用高枝剪在每个个体的外围向阳面采集3~5 片完好健康的、成熟叶片进行功能性状的测定,同时采集长度为3 cm 的当年生枝条进行枝条密度(DW)的测定。采集的枝条与叶片均使用自封袋保存带回实验室,择优选取3 片树叶对其叶面积(AL)、叶鲜质量(mLF)及叶干质量(mLD)进行测定,将叶片按照样方进行混合,测定叶片全氮质量分数(ω(NL))、全磷质量分数(ω(PL))和全钾质量分数(ω(KL))。
测定的详细步骤如下:首先将叶片样品表面附着的灰尘擦净,叶面积通过扫描仪(HP G4050,中国)获取图像,使用ImageJ 软件计算叶面积,然后利用电子天平(精度0.000 1 g)测定叶鲜质量,鲜质量测定完成后将样品在75 ℃下烘干至恒质量后称量,获得叶干质量。枝条密度先用排水法测定枝条体积,然后将样品在70 ℃下烘干96 h 至恒质量,然后使用电子天平(GT50,精度0.001 g)测量其干质量,通过枝条干质量与枝条体积之比计算枝条密度[12]。对于叶片全氮、全磷、全钾质量分数的测定,需要将采集的叶片烘干后研磨,使用180 μm 的筛子进行过筛,用硫酸-双氧水消煮,叶片全氮质量分数采用凯氏蒸馏法测定;叶片全磷质量分数采用钒钼黄比色法测定;叶片全钾质量分数采用火焰光度计法测定[13]。
叶片干物质含量(CLDM)、比叶面积(ASL)、叶片氮磷比(ω(NL)∶ω(PL))及枝条密度的计算公式如下:
叶干物质含量=叶干质量/叶鲜质量×100%(1)
比叶面积=叶面积/叶干质量 (2)
叶片氮磷比=叶片全氮质量分数/叶片全磷质量分数 (3)
枝条密度=枝条干质量/枝条体积 (4)
1.4 土壤因子测定
在采集植物样品的同时,在每个20 m×20 m的样方对角线上均匀设置5个取样点取土,去除土壤表层的枯枝落叶,使用环刀取样。利用烘箱和电子天平(精度0.001 g)测定土壤含水量(SWC)及土壤容重(SBD);采集表层土样后均匀混合,带回实验室自然风干后测定土壤化学性质。其中,土壤全氮质量分数(ω(NS))采用半微量凯氏定氮法测定,土壤速效磷(ω(PSA))采用钼锑抗比色法测定,土壤速效钾(ω(KSA))采用醋酸铵火焰光度计法测定[13],土壤氮磷比由土壤全氮质量分数与土壤速效磷的比值获得[14]。
1.5 数据处理
对不同林龄下毛白杨功能性状及环境因子进行变异系数分析和单因素方差分析(one-way ANOVA)并用Tukey法进行多重比较,采用Pearson法分析功能性状之间的相关关系,并用主成分分析法确定不同林龄下毛白杨功能性状变化的主要指标。以功能性状值为因变量,环境因子为自变量进行多元逐步回归,确定影响毛白杨功能性状的主要环境因子。数据的初步整理在Excel 中进行,数据处理和作图在R4.1.2中完成。
2 结果与分析
2.1 不同林龄毛白杨功能性状差异
在不同林龄下,各功能性状都存在不同程度的变异(见表2),叶片氮磷比变异系数最大,枝条密度最小,表明叶氮磷比受林龄的影响较大,而枝条密度则较稳定。毛白杨功能性状的变异系数由大到小依次为叶片氮磷比、叶片全氮质量分数、叶片全磷质量分数、叶干质量、叶片全钾质量分数、叶鲜质量、叶面积、比叶面积、叶干物质含量、枝条密度。其中,叶片氮磷比、叶片全氮质量分数、全磷质量分数、叶干质量及叶全钾质量分数变异系数大于50%,变动幅度较大,属于强变异;叶鲜质量、叶面积及比叶面积为中等变异;叶干物质含量与枝条密度变动幅度较小,属弱变异。
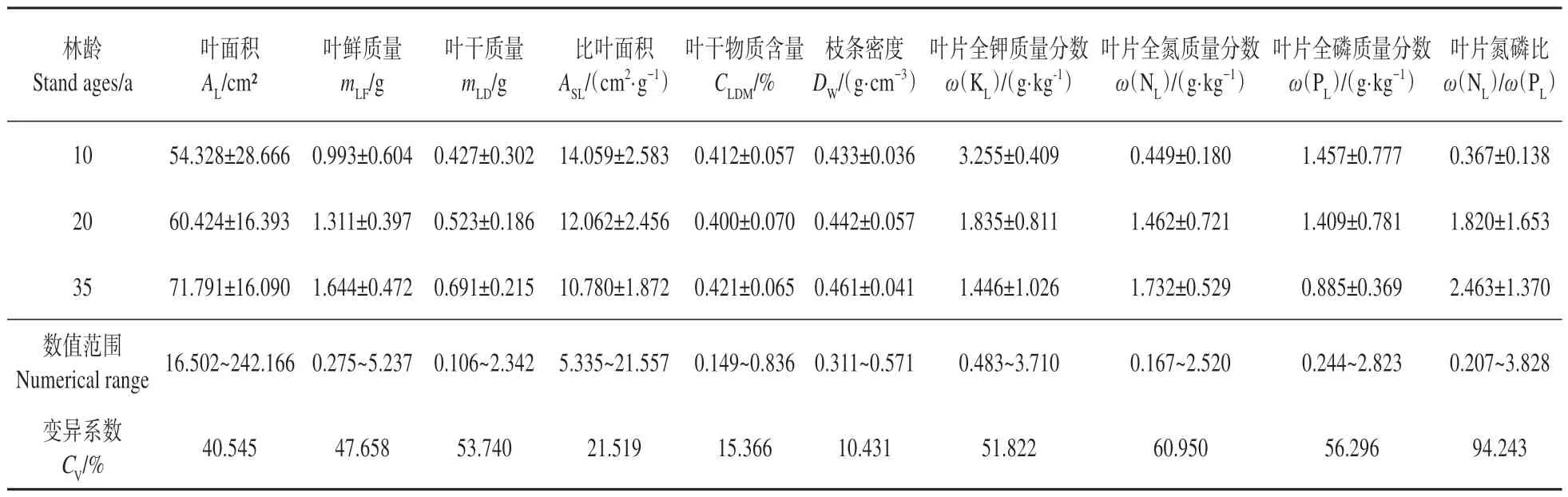
表2 不同林龄毛白杨功能性状的变异系数Table 2 Coefficient of variation of functional traits in plantation P. tomentosa of different stand ages
毛白杨功能性状均随林龄的增长而变化(图1)。叶面积、叶鲜质量、叶干质量、叶干物质含量、枝条密度、叶片全氮质量分数和叶氮磷比均呈现出随林龄增加而增长的趋势。其中,叶面积、叶鲜质量和叶干质量在不同林龄下差异显著(P<0.001,图1:A~C),在35 a 林龄时达到较高水平。叶干物质含量虽变化微弱,但仍呈增长趋势(P<0.05,图1E)。枝条密度在20 a 林龄时达到最高(P<0.001,图1F),后趋于稳定。叶片全氮质量分数与叶氮磷比在20 a 林龄时达到最大,与10 a 林龄差异显著(P<0.01,图1H,J),并在35 a林龄时趋于稳定。比叶面积、叶片全钾质量分数及叶片全磷质量分数呈现出随林龄的增长而逐渐减小的趋势。比叶面积随林龄的增加而逐年减小,在不同林龄下差异显著(P<0.001,图1D);叶片全钾质量分数在10 a林龄时较高,且与20、35 a 林龄差异显著(P<0.001,图1G),在20 a 林龄往后差异变小,趋于稳定;叶片全磷质量分数虽呈下降趋势,但在不同林龄下差异不显著(图1I)。
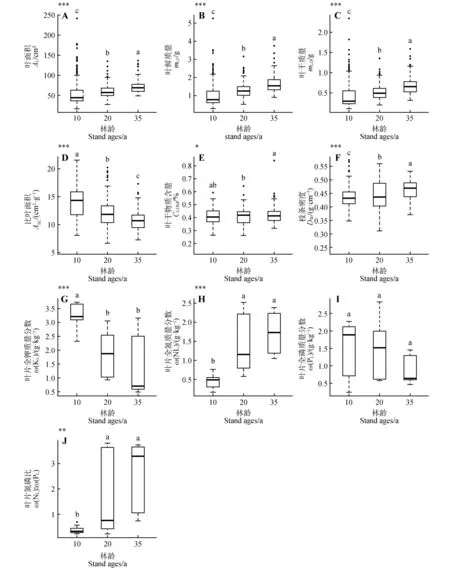
图1 不同林龄毛白杨功能性状的变化*P<0.05,**P<0.01,***P<0.001,为单因素方差分析结果,表示在3个林龄下,毛白杨功能性状的差异显著,不同小写字母表示每个林龄之间差异显著(Tukey检验,P<0.05);下同Fig.1 Variation of functional traits at different stand ages of Populus tomentosa*P<0.05,**P<0.01,***P<0.001,indicated significant differences according to a one-way ANOVA,different letters indicated significant differences in different stand age(sTukey’s test;P<0.05);the same as below
2.2 不同林龄毛白杨林下环境因子变化
土壤因子在不同林龄毛白杨人工林林下的差异显著(见表3)。其中,土壤全氮、土壤速效钾质量分数呈现随林龄的增长而增大的趋势;土壤含水量、土壤氮磷比均呈现随林龄增长先增加后减少的趋势,在20 a 林龄下最高;土壤容重、土壤速效磷、林分密度(DS)均呈现随林龄增长而减小的趋势。
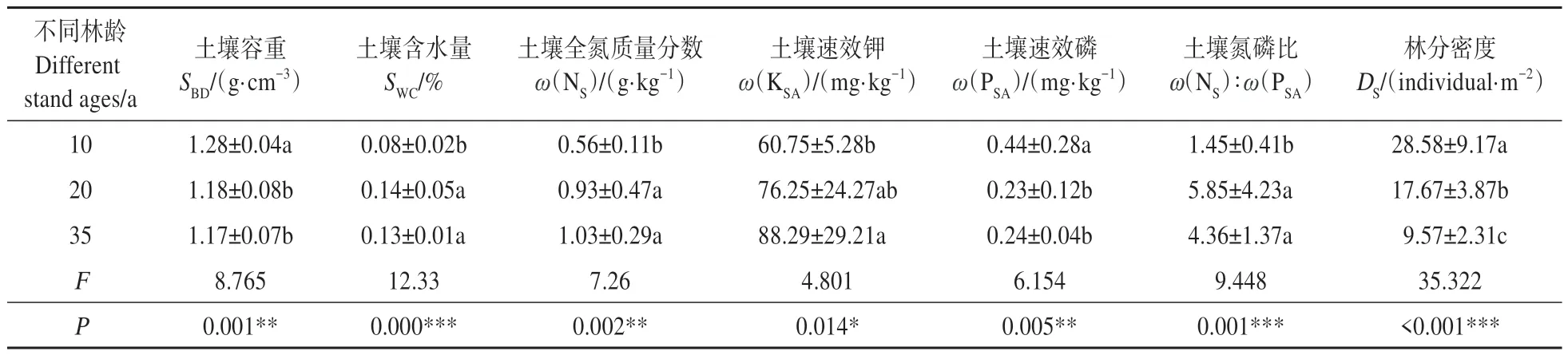
表3 不同林龄毛白杨人工林林下环境因子的差异Table 3 Differences in understory environmental factors of plantation Populus tomentosa with different stand ages
2.3 植物功能性状相关性分析
毛白杨功能性状间的相互作用强烈(见图2)。叶片全磷质量分数与叶干物质含量、叶片全钾质量分数呈极显著正相关,与叶干质量呈显著正相关,与叶片全氮质量分数、叶氮磷比、比叶面积呈极显著负相关。叶干物质含量与枝条密度、叶干质量、叶面积存在极显著正相关,与叶全钾质量分数呈显著正相关,与叶氮磷比、比叶面积呈极显著负相关,与叶全氮质量分数呈显著负相关。枝条密度与叶干质量呈极显著正相关,与叶鲜质量呈显著正相关,与比叶面积呈极显著负相关。叶干质量与叶面积、叶鲜质量呈极显著正相关,与比叶面积呈极显著负相关。叶面积与叶鲜质量呈极显著正相关,与比叶面积呈极显著负相关。叶鲜质量与叶全氮质量分数呈极显著正相关,与叶氮磷比呈显著正相关,与比叶面积呈极显著负相关,与叶全钾质量分数呈显著负相关。叶全氮质量分数与叶氮磷比呈极显著正相关,与叶全钾质量分数呈极显著负相关。
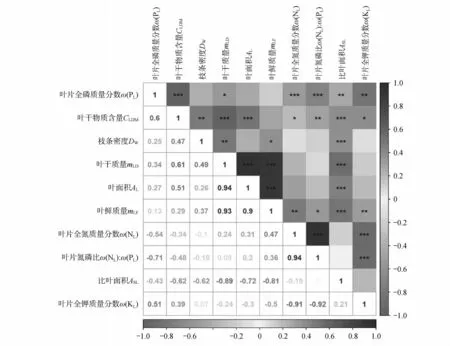
图2 毛白杨功能性状间的Pearson分析*表示在0.05水平上显著相关(P<0.05);**表示在0.01水平上显著相关(P<0.01);***表示在0.001水平上显著相关(P<0.001)Fig.2 Pearson analysis among functional traits of Populus tomentosa* indicated significant correlation at the 0.05 leve(lP<0.05);** indicated significant correlation at the 0.01 leve(lP<0.01);*** indicated significant correlation at the 0.001 leve(lP<0.001)
对所有功能性状进行PCA分析(见表4),枝条密度的公因子方差小于0.4,表明枝条密度与其他性状之间关系薄弱,需删除后重新分析。PCA 分析发现,各指标的公因子方差均较大,意味着指标与主成分之间存在较强的相关性。共提取2 个主成分,特征值分别为4.308 和3.637,贡献率分别为47.868%和40.108%,累计贡献率达88.276%。说明这2 个主成分是解释叶性状变化的主要因素。主成分1与叶面积、叶鲜质量及叶干质量呈显著正相关,比叶面积与其呈显著负相关,表明主成分1主要反应叶片的形态特征;主成分2与叶干物质含量、叶全钾及叶全磷质量分数呈显著正相关,表明其主要反应叶片的养分特征。由此可见叶面积、叶鲜质量、叶干质量及叶全磷质量分数可以作为不同林龄毛白杨功能性状变化的主要指标。由PCA 排序图(图3)可见,主成分1 能够明显区分不同林龄下叶性状指标的差异。
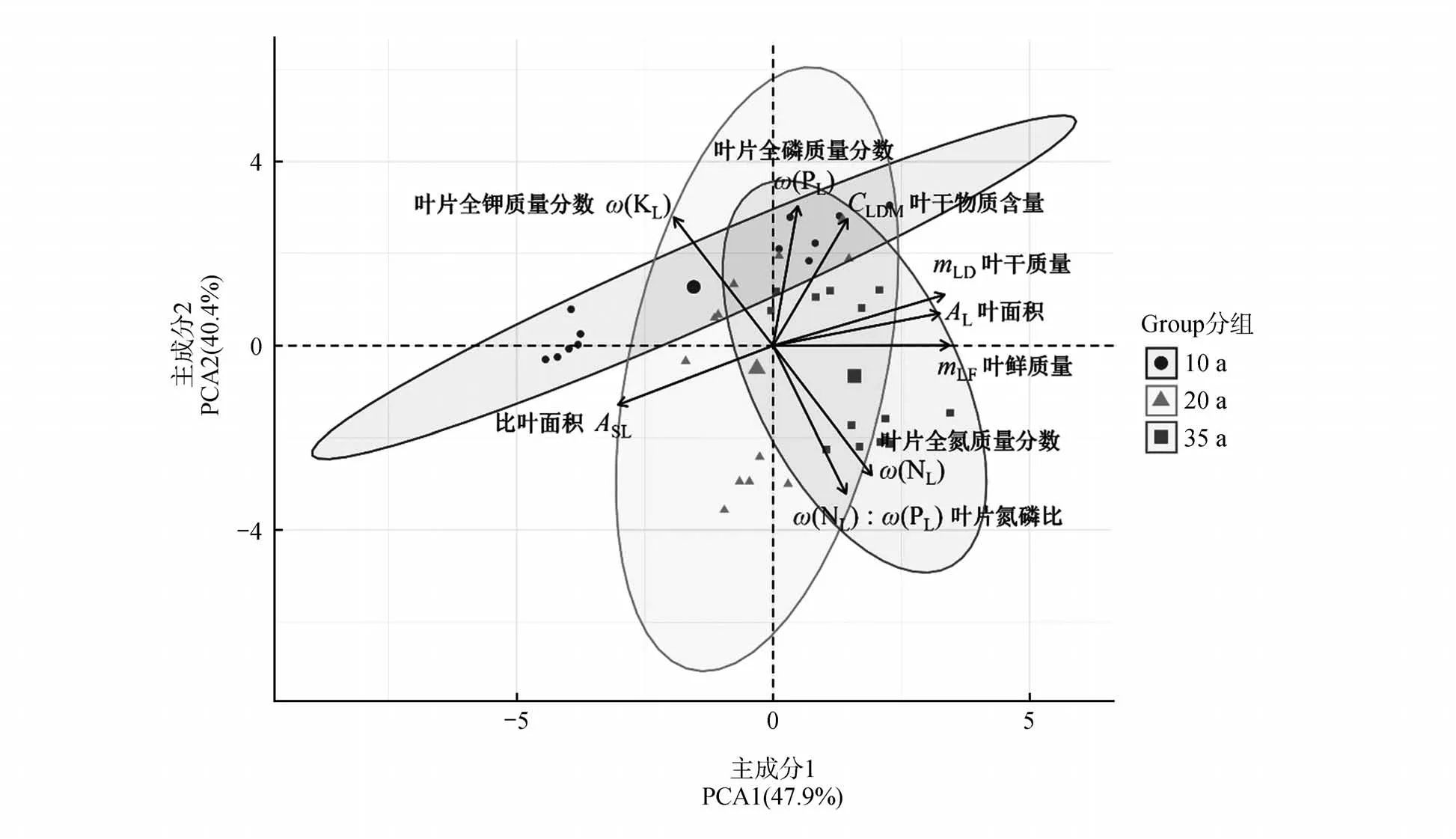
图3 主成分分析排序Fig.3 PCA ranking chart
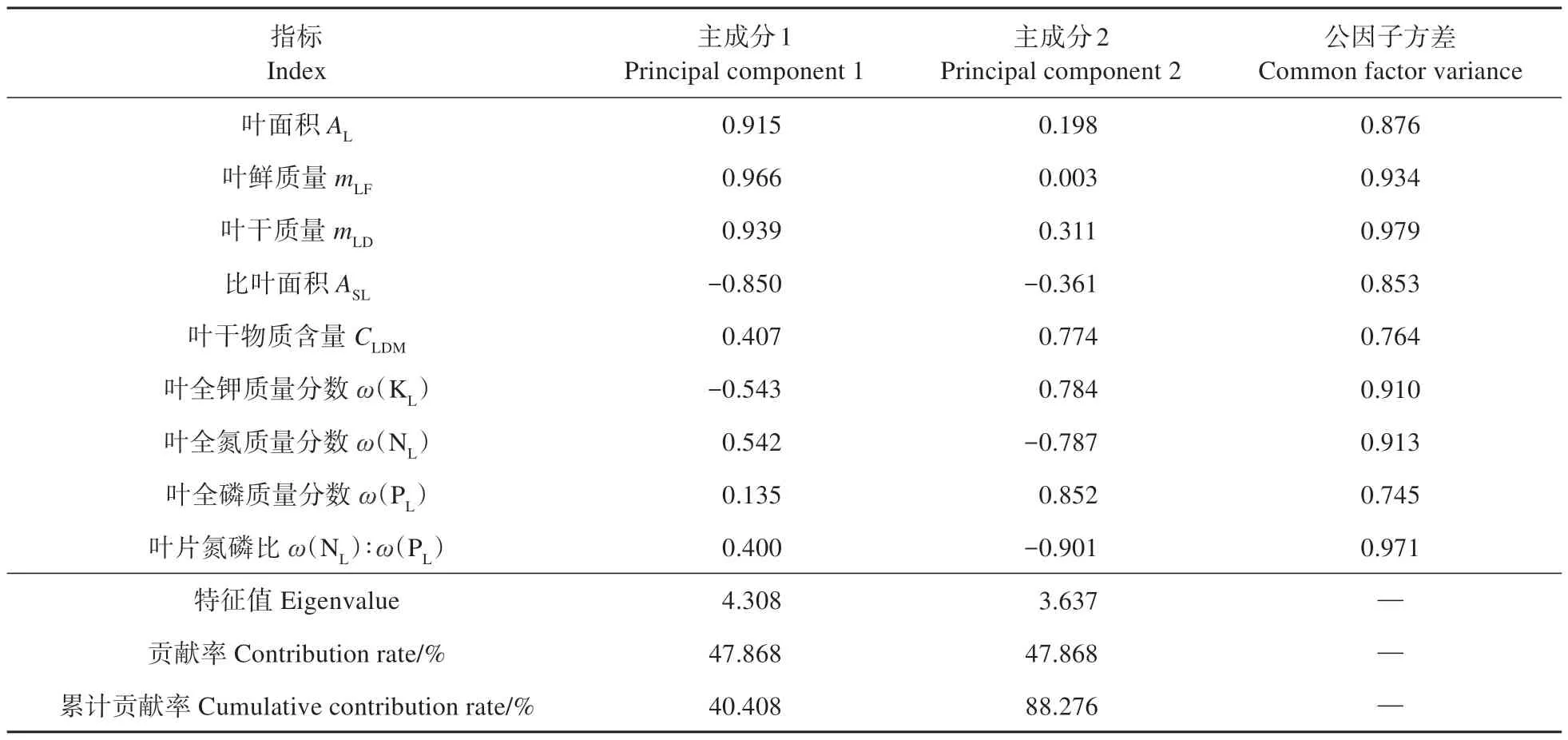
表4 各性状PCA载荷矩阵及主成分贡献率Table 4 PCA loading matrix and principal component contribution of each trait
2.4 植物功能性状与环境因子的关系
逐步回归分析的结果表明(表5),各性状都与环境因子呈显著关系,其中林分密度、土壤速效钾、土壤容重及土壤含水量是影响毛白杨功能性状的主要环境因子。
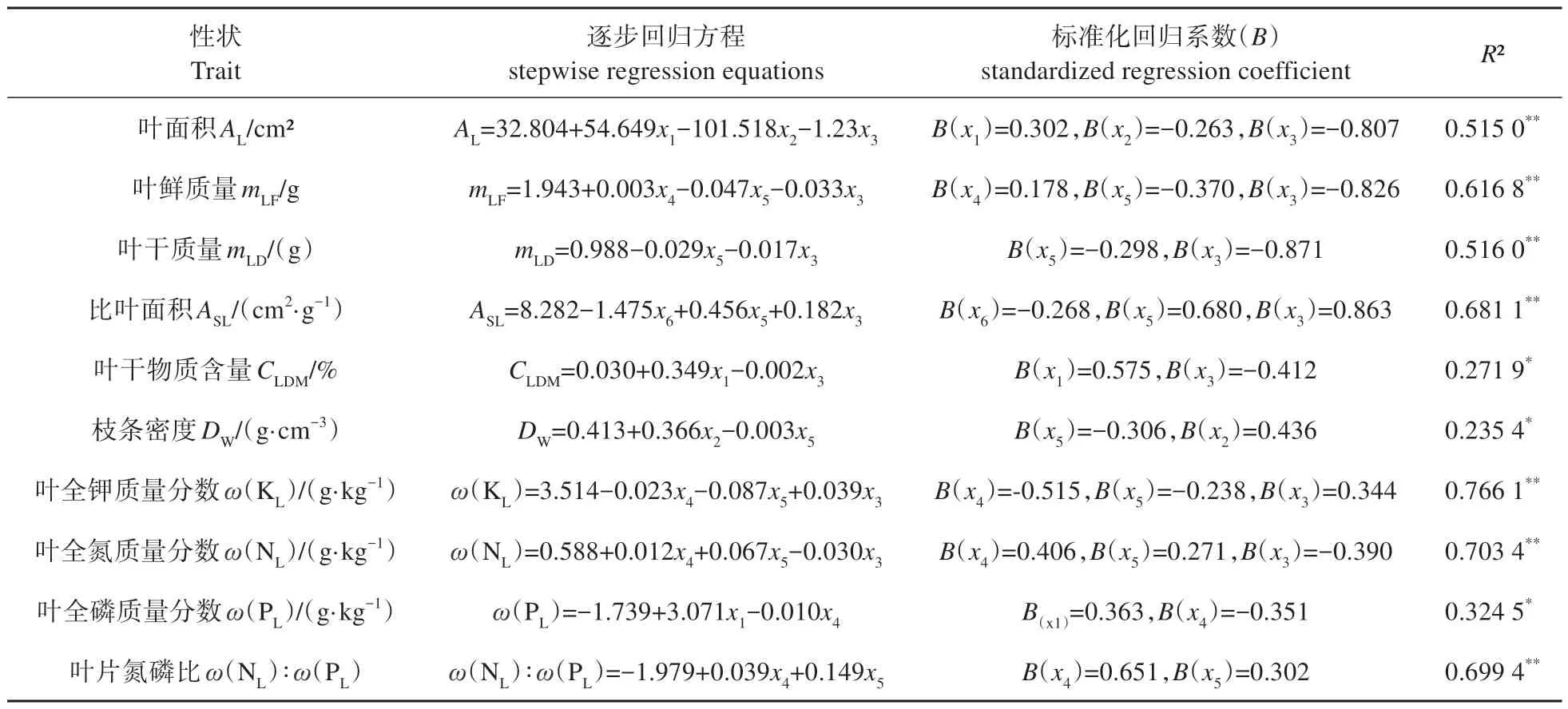
表5 功能性状和环境因子之间的逐步回归分析Table 5 Stepwise regression analysis between functional traits and environmental factors
表5 显示,叶面积与土壤容重、土壤含水量及林分密度显著相关;叶干质量与土壤氮磷比、林分密度显著相关;比叶面积与土壤全氮质量分数、土壤氮磷比及林分密度显著相关;叶鲜质量、叶全钾、叶全氮与土壤速效钾、土壤氮磷、林分密度显著相关。其中,在叶面积、叶鲜质量、叶干质量及比叶面积的回归方程中,林分密度的回归系数最大,因此叶鲜质量受林分密度影响最大,叶面积、叶鲜质量、叶干质量与林分密度呈负相关,并随密度的增加而减小;而比叶面积与其呈正相关。叶全钾、叶全氮和叶氮磷比受土壤速效钾影响最为显著,叶全钾与土壤速效钾呈负相关,表明土壤速效钾的减少则有更多的养分流入植物之中,叶片全氮及叶氮磷比与土壤速效钾呈正相关。叶干物质含量与土壤容重呈正相关,与林分密度负相关;枝条密度则受土壤含水量影响显著。
3 讨论
3.1 不同林龄毛白杨功能性状的变化规律
植物功能性状受遗传因素与环境因子的共同影响[15]。对不同林龄的毛白杨功能性状进行变异系数分析,可以看到各功能性状的变异系数为10.658%~94.243%。枝条密度在不同林龄条件下变化最为稳定,而叶片氮磷比波动较大。本研究中,枝条密度是相对较为稳定的性状,这可能是由于枝条密度具有较高的遗传控制,因而始终保持相对稳定的状态[16]。叶片氮磷含量主要受环境影响[9],不同林龄叶片氮磷比的变化由叶片氮磷含量的变化引起,出现这种情况的原因可能是由于植物在不同的生长时期对养分的需求不同。
植物叶功能性状与植物对资源的获取、利用及利用效率的关系最为密切。叶面积能衡量叶片的光合能力,叶面积的增大代表植物光合能力强,生长速率快[6,15],叶干物质含量能够更好反映植物对资源的获取和利用能力[15]。在本研究中,叶面积、叶鲜质量、叶干质量、叶干物质含量皆随林龄的增长而增大,表明毛白杨在生长后期更注重对资源的保持与对外界干扰的抵抗。比叶面积影响着叶片的光资源获取,与生长速率呈正相关[6]。本研究中,比叶面积在10 a 林龄时达到最高,后随着林龄的增加而逐渐减小,这可能是因为在10 a 林龄下,林分密度较高,植物可利用的生长空间较小,为了在生长前期保持较高的资源获取能力与生长速率,植物会通过缩小叶面积、降低自身消耗来增大比叶面积。于青含等[17]对红松(Pinus koraiensis)枝性状的变异研究表明,枝条密度随胸径的增长而增大,与本研究结果类似。这可能是因为随着植株的成熟,树木胸径随之增大,植物由早期快速获取资源的阶段转变为更注重对环境变化的适应,因此枝条密度会随林龄的增加而增大。
林龄与植物养分限制具有密切关系,叶片全钾质量分数对维持植物生长有着重要意义,对陆地生态系统植物生产力具有限制作用[18],叶片中的氮、磷元素是植物合成蛋白质和遗传物质的主要元素[9]。叶全钾随林龄的增加而降低,在20 a林龄后趋于稳定,这可能与植物自身的发育有关。在生长前期对钾的需求较高,因而大量吸收土壤中的钾元素,以提升生长速率、增强抗逆性;到了生长后期,植物整体趋于稳定,对钾的需求不如早期旺盛,这也解释了土壤中钾元素在生长后期增多的原因[19]。叶片氮磷比可以用来表征环境对植物生长的养分供应状况[10]。已有研究证实植物氮磷比小于14,表示植物生长受氮限制较大,氮磷比大于16,表示受磷的限制更大[20]。在本研究中,三个林龄的叶片氮磷比依次为0.367、1.820和2.463,皆远低于14,表明该区域毛白杨生长主要受氮限制。其中10 a 林龄中叶片氮磷比显著低于20 a 与35 a 林龄的值,整体呈现增长趋势。结合土壤养分来看,随着林龄的增加,土壤全氮质量分数增多,速效磷减少,因而使叶片氮磷比呈现出增长趋势。
3.2 植物功能性状间的相关性
植物的生长过程受环境、生理、系统发育等因素的综合影响,使功能性状之间不断权衡,形成与环境相适应的最优组合,各性状之间存在一定的相关性[8]。叶面积、叶鲜质量和叶干质量是叶片形态特征的基本指标,体现植物对水分的获取能力[3]。在本研究中,3 个性状之间相互作用强烈,结合主成分的结果更加证明这3 个性状是表示叶片形态在不同林龄下产生差异的关键性状。叶全磷与多个养分指标相关密切,主成分的结果证明叶全磷是表征叶片养分的主要指标。叶全钾和叶全氮随叶全磷的变化而变化,叶干物质含量和叶干质量随叶全磷的增加而增加,比叶面积随叶全磷的增加而减少。这是因为植物中的氮磷钾是生长发育必需的养分,三者相互协同[9]。叶片中的氮、磷影响着叶片中蛋白质和核酸的合成[9],因而叶片中氮、磷质量分数的高低也影响着叶干质量和叶干物质含量的大小。
叶干物质含量与叶面积、叶干质量、枝条密度和叶片磷、钾质量分数显著正相关,与比叶面积、叶片氮质量分数及叶氮磷比呈显著负相关,这是植物随着林龄的增加与对环境的适应,对光能与养分的获取能力增强,植物体内的养分得到积累,促使合成更多的蛋白质,从而使叶面积、叶干物质含量、叶干质量增大;到了生长后期,生长速率减缓,对资源的需求降低,植物通过减小比叶面积来降低蒸腾作用,为了提升水分的利用效率,使枝条密度增大。综上,植物功能性状会随着林龄的增加,改变自身的器官形态与养分含量来更好地适应环境,提升自身生长速率。
3.3 环境因子与不同林龄毛白杨功能性状的关系
本研究中,林分密度与叶面积、叶鲜质量、干质量呈极显著负相关,与比叶面积呈极显著正相关。这表明在人工林的培育中,适当的林分密度有助于植株自身对资源的获取,能够更好地促进森林的生长发育。叶片大小能表征植物捕获光能的能力,是长期适应环境的结果[15]。林分密度影响叶片对光能的获取,低林分密度让植株间有更大的生长空间,使植株通过增大叶面积来获取更多的光能,提升叶光合能力,增强其资源利用能力[6]。在本研究中,比叶面积随林龄的增加而减小,这与林分密度有关。在生长后期,林分密度减小,植物可利用的生长空间增多,使叶片之间竞争减弱,因而比叶面积随之降低。
土壤理化性质能够显著影响植物功能性状格局的变化,并且叶性状能够很好地表达植物对土壤养分梯度的响应[21]。本研究结果表明,林分密度、土壤速效钾、土壤容重及土壤含水量是显著影响毛白杨功能性状的因子。土壤速效钾与叶全钾、叶氮磷比呈极显著负相关,与叶全氮呈显著正相关。本研究中土壤速效钾与叶全钾呈负相关,土壤速效钾与叶氮磷比呈现正相关。这可能是因为幼龄林的林分密度较大,栽植紧密,土壤容重大,使土壤中养分含量少[19],同时,幼龄植株对养分需求较高,所以形成了土壤速效钾与叶全钾的负相关局面;随着林龄的增长,林分密度逐渐减小,土壤容重变小,土壤中养分累积增多,但植株随着林龄的增长,对钾元素的需求也逐渐降低,因而再次形成了土壤中钾元素多,而植株中少的情况,使土壤速效钾与叶全钾始终为负相关的关系。土壤容重与土壤紧实度有关,决定了土壤是否会出现板结等情况。在幼龄时,土壤容重大,易产生板结等不利情况,土壤容重与叶干物质含量和叶全磷呈显著正相关,表明当植物处于恶劣环境时,会通过增大叶干物质含量和叶全磷来抵御不利环境。土壤含水量与枝条密度呈显著正相关,前人的研究表明,影响枝条生长的主要因素是水分,随着枝条密度的增加,储水能力会逐渐减弱[22]。枝条密度在植物的生长前期较低,有助于水分的运输,加快生长速率,到了生长后期,枝条密度会随着生长速率的减慢而增大,产生更高的耐受能力,也更具有抗旱性[4]。本研究中枝条密度与土壤含水量呈正相关,与前人的研究[23]相反,这可能是因为毛白杨属于耐干旱的树种,拥有较高的枝条密度;同时,所处环境的土壤有沙化倾向,保水能力较弱,土壤环境较为干燥,毛白杨为保证自身水分含量因而使自身枝条密度增大。
4 结论
毛白杨会根据不同生长时期对养分的需求与对环境的适应,通过一定的性状权衡使自身处于最优生长空间。本研究发现,在不同林龄下,毛白杨的枝条密度较为稳定,叶片养分变异较大。随着林龄的增加,林分密度减小,叶片为了获取更多的养分与资源会改变叶片形态特征,更注重对资源的保持与对外界干扰的抵抗。在该区域毛白杨叶片氮磷比远低于14,表明受氮限制较大,可以适当的施以氮肥以利于其生长。随着林龄的增加,土壤环境得到改善,土壤全氮与土壤速效钾都呈增长趋势,土壤容重呈减小趋势,这些变化都有利于毛白杨更好的生长。在各环境因子中,林分密度、土壤速效钾、土壤容重及土壤含水量是主要影响毛白杨生长的环境因子。在不同林龄下,毛白杨的功能性状差异显著,表明其在兼顾生长与环境的变化中有着较强的适应性,在生长后期逐渐形成抗逆性更强的性状组合,以利于其在鲁西黄泛平原的生长。研究结果有助于深入理解黄泛平原毛白杨对环境变化的响应及生态适应策略,为森林可持续管理和生态系统稳定提供理论依据。
猜你喜欢
今日农业(2022年14期)2022-11-10
防护林科技(2020年10期)2020-12-22
新农民(2020年21期)2020-12-08
东坡赤壁诗词(2018年1期)2018-03-31
河南林业科技(2017年1期)2017-06-10
广东农业科学(2016年7期)2016-10-08
防护林科技(2016年9期)2016-09-27
防护林科技(2016年5期)2016-09-05
亚热带资源与环境学报(2015年1期)2015-01-22
湖南农业科学(2014年14期)2014-02-27
