
基于叶片和果实性状的糠椴天然居群表型变异及多样性分析
2023-11-13 09:27:08穆怀志王清玉吕松瞳靳旭红夏富才
植物研究 2023年6期
穆怀志 王清玉 张 玥 吕松瞳 靳旭红 张 骁 夏富才*
(1.北华大学林学院,吉林 132013; 2.吉林省林业勘察设计研究院,长春 130022)
糠椴(Tilia mandshurica)为椴树科(Tiliaceae)椴树属(Tilia)高大乔木,主要分布在中国东北、朝鲜半岛以及俄罗斯西伯利亚南部地区,其树形通直、树姿美丽,是珍贵的阔叶用材和绿化树种;糠椴为虫媒花,主要通过蜜蜂采蜜过程中进行传粉,是东北地区重要的蜜源树种[1]。然而长期不合理的采伐,导致糠椴资源遭受严重破坏[2]。目前,糠椴天然居群处于生长衰退期,自然更新缺乏,个体数量不断减少,加之现有天然居群的背景和多样性水平不清晰,导致糠椴种质资源的保护及改良受到严重制约[3]。
植物的表型性状具有环境可塑性,反映了植物对环境因子的适应程度,是研究植物变异和进化的重要标记[4-5]。天然居群表型性状的变异及多样性能够反映居群变异的大小,是揭示居群适应性的有效途径[6-7]。近年来,众多学者开展了植物天然种群变异和多样性研究。楼明祝等[8]调查了新疆9 个野生黄花苜蓿(Medicago falcata)居群的22个表型性状,发现黄花苜蓿变异丰富,其变异主要来自居群间。米老排(Mytilaria laosensis)各表型性状在种源内的变异均大于种源间的变异,种源间平均表型分化系数高达35.10%[9]。无患子(Sapindus mukorossi)天然居群内的多样性高于居群间,Nei’s 多样性指数分别为0.390 9 和0.298 0[10]。上述研究为植物种质资源保存提供参考。目前,关于糠椴表型多样性缺乏全面且深入的研究。针对这一问题,本研究对东北地区8个糠椴天然居群的11 个叶片和果实表型性状进行分析,探讨各表型性状的变异和多样性,以期为糠椴种源区划和糠椴天然林可持续保护利用提供科学依据。
1 材料与方法
1.1 供试材料
在东北林区的糠椴天然分布区内选择东丰、辉南、敦化、珲春、桦甸、龙潭、蛟河和萝北8个天然居群,分别位于吉林省辽源市、通化市、延边朝鲜族自治州、吉林市和黑龙江省鹤岗市。各居群生境概况见表1。
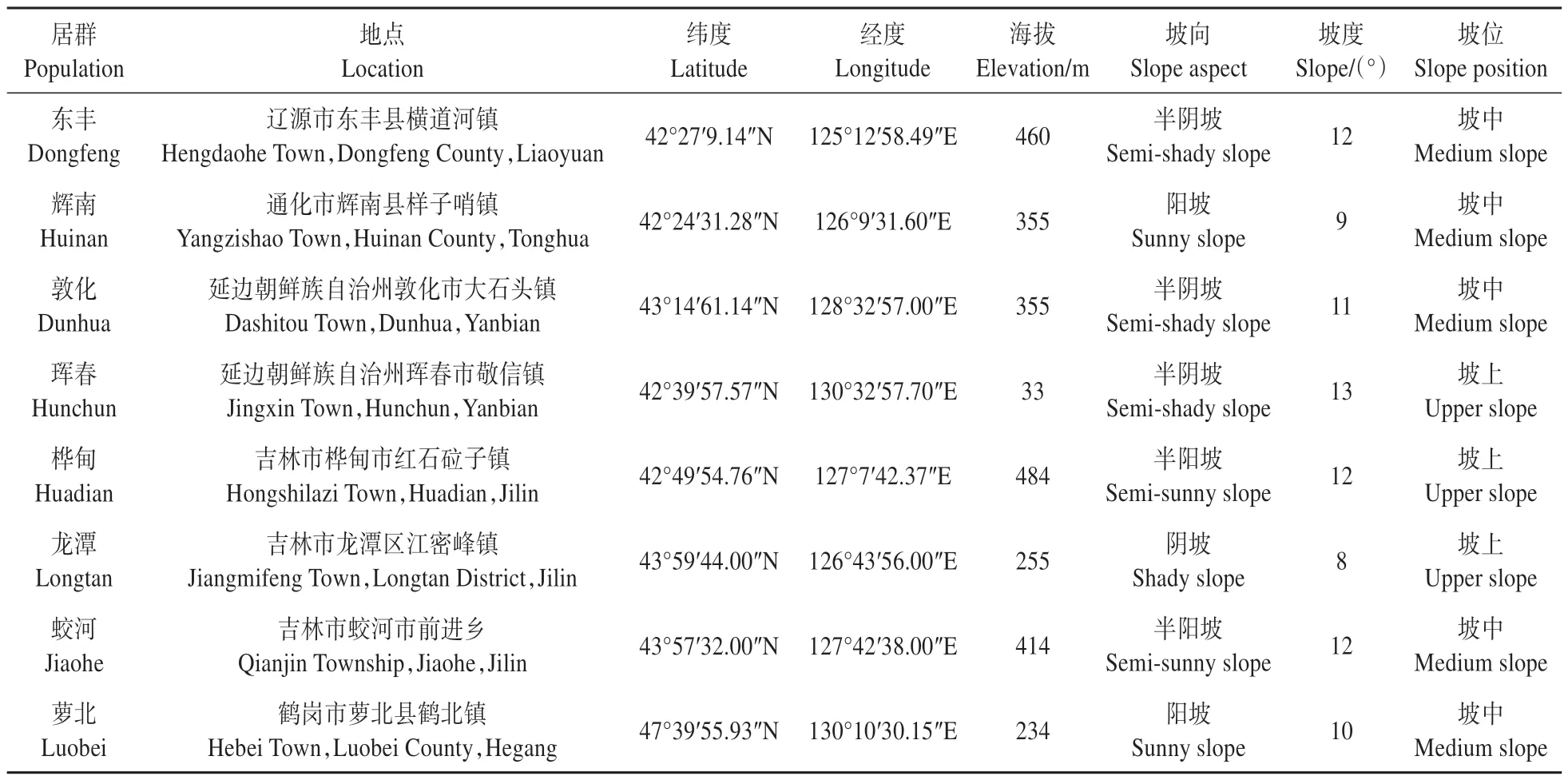
表1 供试糠椴天然居群生境概况Table 1 Habitat status of T.mandshurica natural populations tested
参照严灵君等[11]的研究方法以及蜜蜂授粉的有效范围[12],于2022 年9 月末至10 月初糠椴果实成熟期,在每个糠椴天然居群中随机选取10 株生长正常、无严重缺陷、无病虫害的糠椴单株作为采样株。采样株树龄30 a 以上,胸径不低于12 cm,树高不低于10 m,采样株间距30 m 以上。在每个糠椴采样株的向阳方向采集树冠中部的枝条,选取枝条中部的成熟叶片和果实各10 枚,放入带有冰袋的保温箱中用于表型性状测定。
1.2 叶片和果实性状测定
利用Microtek 便携式扫描仪(ScanMaker i800 Plus,上海)测量叶长、叶宽、叶柄长和叶柄宽,利用游标卡尺测量果实纵径、果实横径、果柄长、苞叶长、苞叶宽、苞果合生长、果序柄长,测量精确到0.1 mm。
1.3 数据分析
利用SAS v9.21 软件对获得的数据进行统计分析。采用嵌套模型对不同糠椴居群的表型性状进行方差分析,并利用Duncan 法进行多重比较[13]。在此基础上,采用表型分化系数分析糠椴表型性状差异的主要来源[14];利用变异系数反映糠椴不同表型性状的变异程度[15];采用Shannon-Wiener 多样性指数分析糠椴居群表型性状的多样性程度[16];利用主成分分析对糠椴居群进行评价[17];采用Ward 法对糠椴居群进行聚类分析[18]。
方差分析的线性模型为:
式中:Yijk为第i个天然居群,第j个采样株,第k个性状的测量值;μ为模型截距对模型的影响;Si为第i个天然居群对模型的影响;T()i j为嵌套在第i个天然居群内第j个采样株对模型的影响;zijk为系统误差对模型的影响。
表型分化系数计算公式为:
式中:Vst为表型分化系数;δ2t s为居群间的方差分量;δ2s为居群内的方差分量;S表示天然居群;t表示天然居群的单株。
变异系数计算公式为:
式中:CV为变异系数;S为表型标准差;xˉ为某一性状群体平均值。
Shannon-Wiener多样性指数计算公式为:
式中:H′为Shannon-Wiener 多样性指数;Pi为某一性状第i级材料内分布频率的有效百分比。
主成分分析综合指数计算公式为:
式中:Y为综合指数;m为每个主成分组下指标数;n为每组下各指标总样本数;Wij为第j指标第i样本的权重数;Hij为第j指标第i样本原始数据正规化后的值。
2 结果与分析
2.1 糠椴居群表型差异
对糠椴居群间和居群内的叶片和果实性状进行方差分析(见表2),结果表明:叶长、叶宽、叶柄长和叶柄宽在居群间和居群内的差异均达到极显著水平,果实纵径、果实横径、果柄长、苞叶宽、苞叶长、苞果合生长和果序柄长在居群间和居群内的差异也均达到极显著水平。为了进一步明确糠椴居群间和居群内表型性状的差异,对各性状的方差分量和表型分化系数进行计算(见表3),发现各性状居群间的方差分量百分比均大于居群内,说明居群间是糠椴表型性状差异的主要来源。居群间,果实横径的方差分量百分比最大(79.49%),叶宽的方差分量百分比最小(56.87%);居群内,叶长的方差分量百分比最大(11.75%),叶柄长的方差分量百分比最小(4.09%)。在表型分化方面,叶柄长的表型分化系数最大,高达0.95,叶长的表型分化系数最小,仅为0.83。
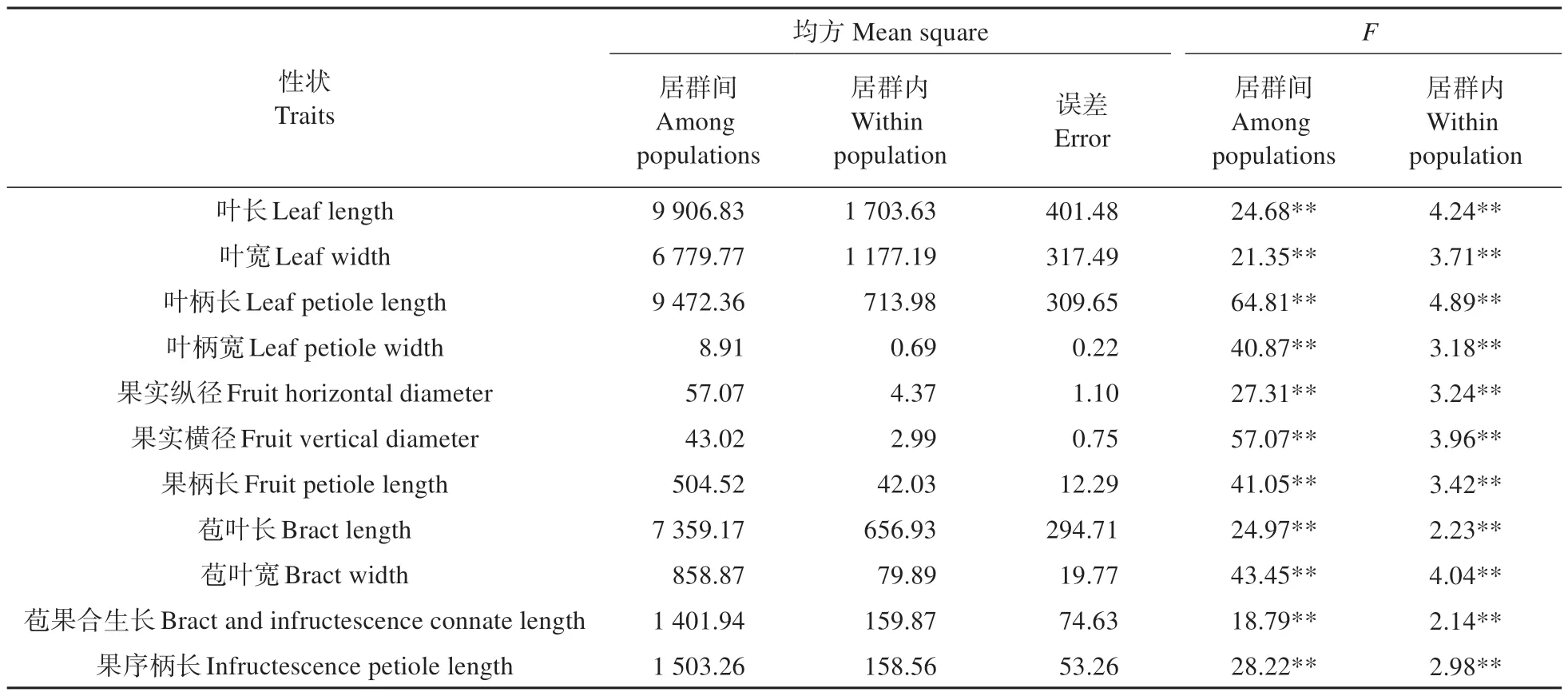
表2 糠椴天然居群叶片和果实性状方差分析Table 2 Variance analysis on leaf and fruit traits of T.mandshurica natural populations
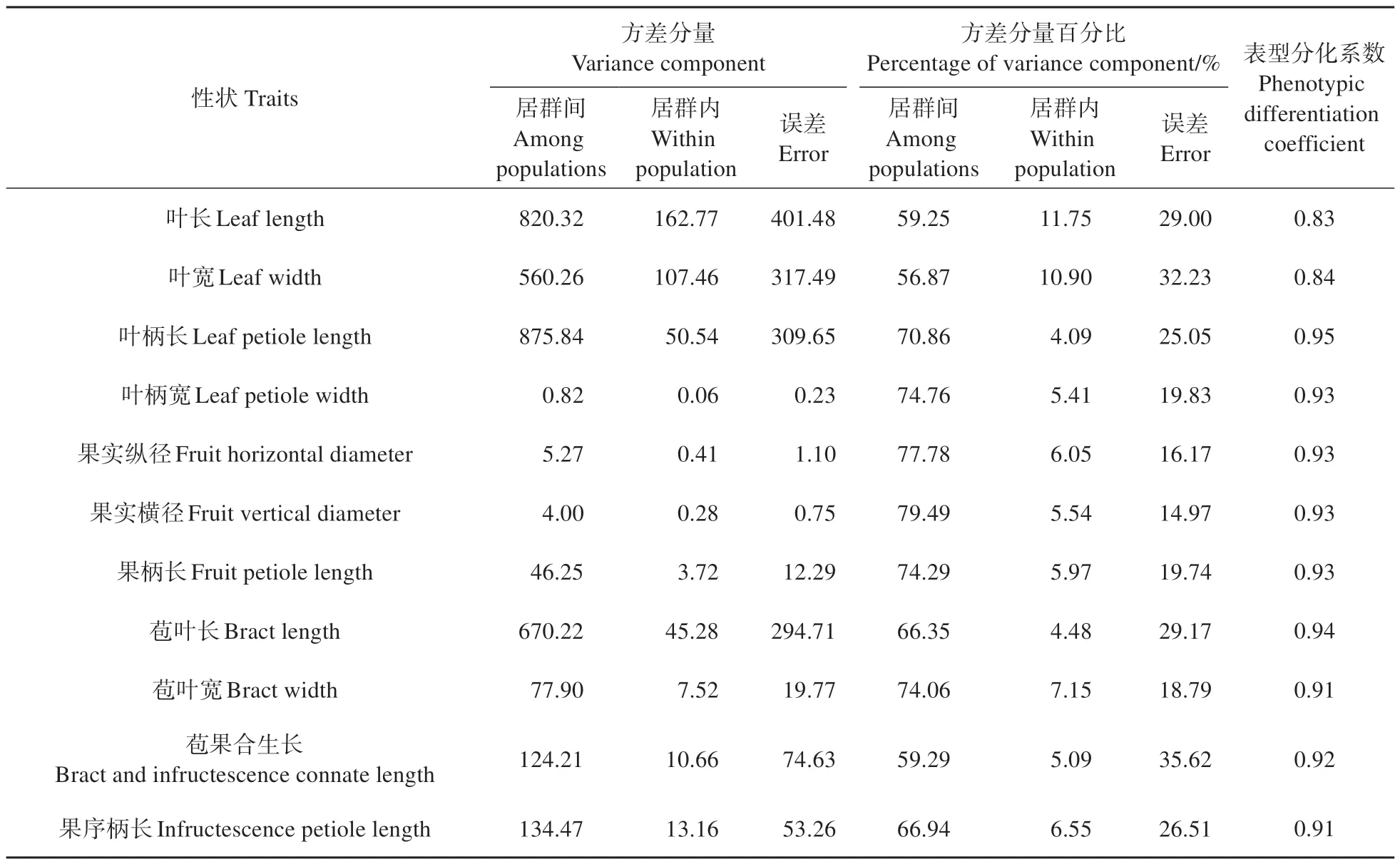
表3 糠椴天然居群叶片和果实性状的方差分量和表型分化系数Table 3 Variance component and phenotypic differentiation coefficient of leaf and fruit traits in T.mandshurica natural populations
2.2 糠椴居群表型变异与多样性
对糠椴居群间的叶片和果实性状进行多重比较,并计算变异系数和Shannon-Wiener 指数(见表4),结果表明:糠椴叶长、叶宽、叶柄长和叶柄宽的平均值分别为113.69、98.81、56.48、2.06 mm,果实纵径、果实横径、果柄长、苞叶长、苞叶宽、苞果合生长和果序柄长的平均值分别为9.18、8.01、15.95、85.39、16.06、37.03、27.8 mm。其中,东丰居群的叶长和叶宽均最大,分别为132.77、113.59 mm;萝北居群的叶柄长最大,高达76.31 mm;珲春居群的叶柄宽、果柄长和果序柄长均最大,分别为2.36、18.21、34.19 mm;龙潭居群的果实纵径和果实横径均最大,分别为10.63、9.17 mm;蛟河居群的苞叶长和苞叶宽均最大,分别为97.03、21.08 mm;桦甸居群的苞果合生长最大,达到44.87 mm。糠椴叶片和果实性状的变异系数介于20.0%~39.9%,Shannon-Wiener 指数介于1.262~1.443,各性状变异系数和Shannon-Wiener 指数均较大,说明糠椴天然居群具有丰富的变异和表型多样性,可根据表型性状对糠椴居群进行选择。
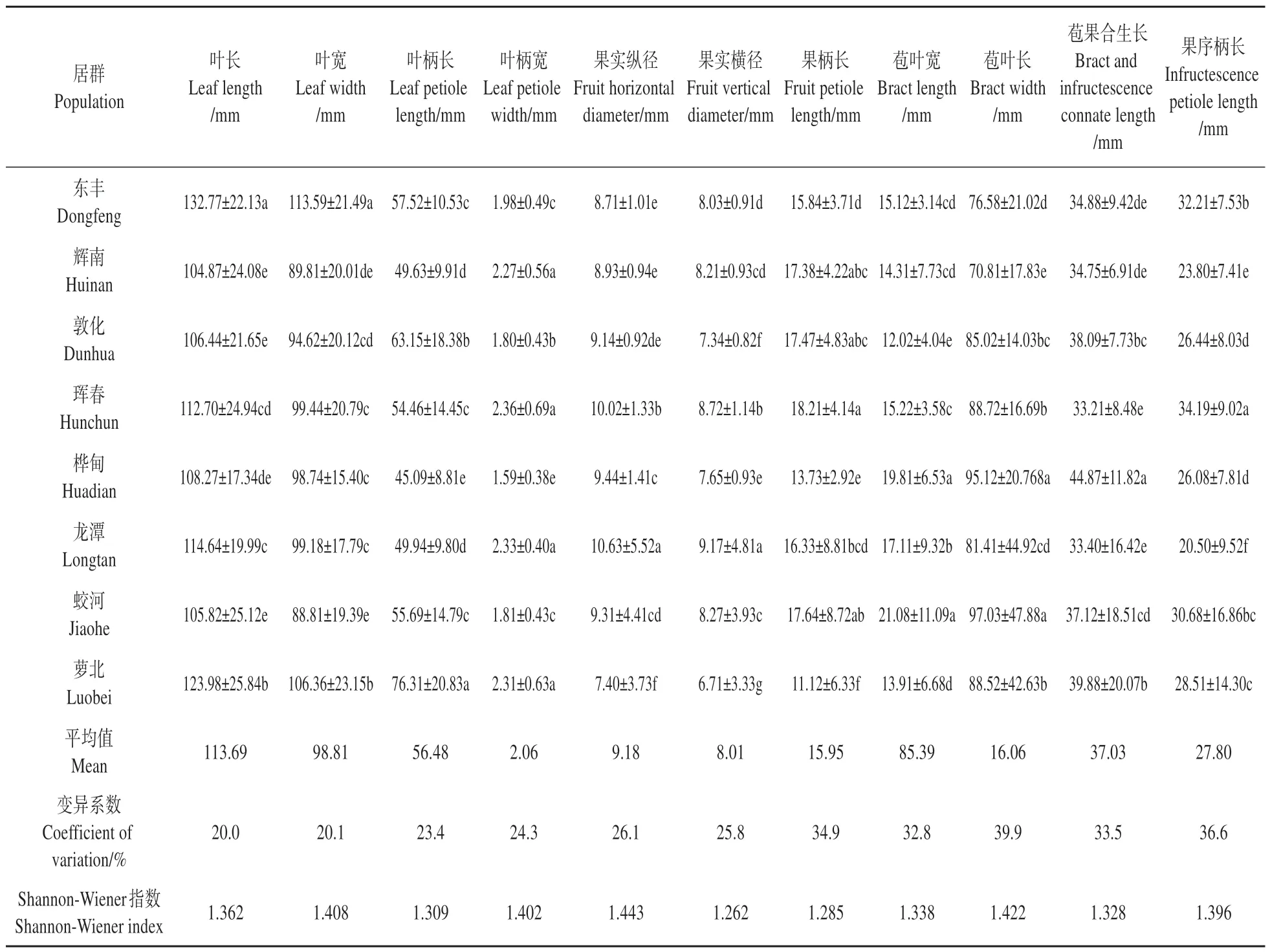
表4 糠椴天然居群叶片和果实性状的多重比较、变异系数和Shannon-Wiener指数Table 4 Multiple comparisons,coefficient of variation and Shannon-Wiener index of leaf and fruit traits in T.mandshurica natural populations
2.3 糠椴居群主成分分析
对糠椴的叶片和果实性状进行主成分分析(见表5),结果表明:11 个表型性状可分为4 个主成分,累计方差贡献率达到88.46%。其中,主成分1 方差贡献率为36.97%,果实纵径和果实横径的特征向量绝对值较大,说明该主成分主要反映果实大小;主成分2 方差贡献率为26.94%,苞叶长和苞果合生长的特征向量绝对值较大,说明该主成分主要反应苞叶特征;主成分3 方差贡献率为13.87%,叶长和叶宽的特征向量绝对值较大,说明该主成分主要反映叶片大小;主成分4方差贡献率为10.68%,果柄长和果序柄长的特征向量绝对值较大,说明该主成分主要反映了果柄和果序柄的大小。以每个主成分对应的特征值占所提取主成分特征值之和的比例为权重,得出各表型性状主成分综合模型为:
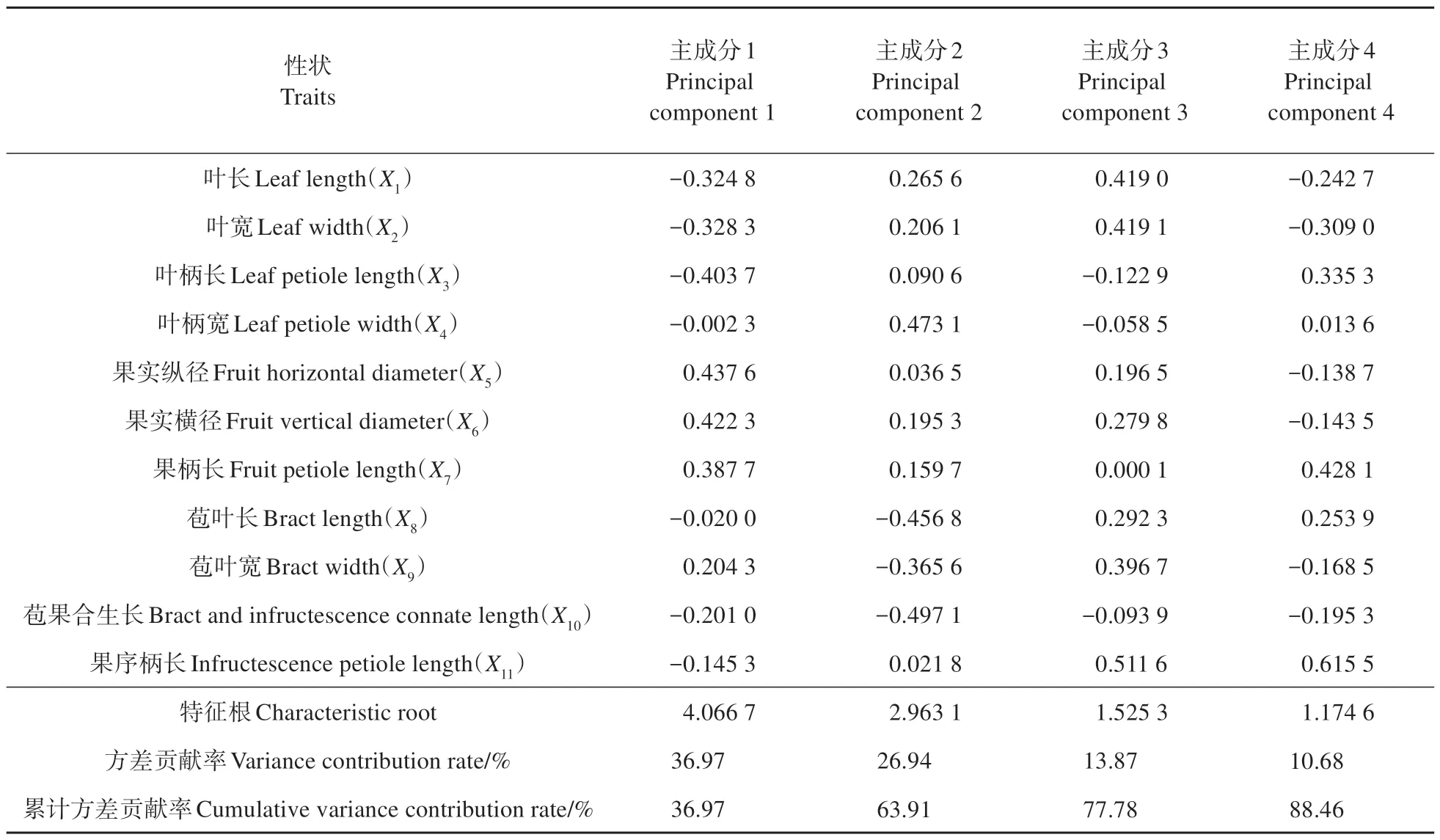
表5 糠椴天然居群叶片和果实性状主成分特征值及贡献率Table 5 Principal component eigenvalues and contribution rates of leaf and fruit traits in T.mandshurica natural populations
根据表型性状主成分综合模型,8个居群综合排序由大到小依次为:珲春、龙潭、辉南、蛟河、东丰、敦化、桦甸、萝北。
2.4 糠椴居群聚类分析
根据糠椴的叶片和果实性状计算聚类系数(见图1),发现当分类数为3 时,聚类系数的上升趋势明显变缓,此时聚类系数为0.631。因此,可以将糠椴8 个居群分为三类(见图2)。第Ⅰ类居群包括东丰和萝北,该类居群叶片性状较为突出,叶片较大,其他表型性状适中。第Ⅱ类居群包括敦化、珲春、桦甸和蛟河,该类居群果柄和苞果性状较为突出,果柄长、果序柄长、苞叶宽、苞叶长和苞果合生长较大。第Ⅲ类居群包括龙潭和辉南,该类居群果实大小性状较为突出,果实纵径和果实横径性状较大。
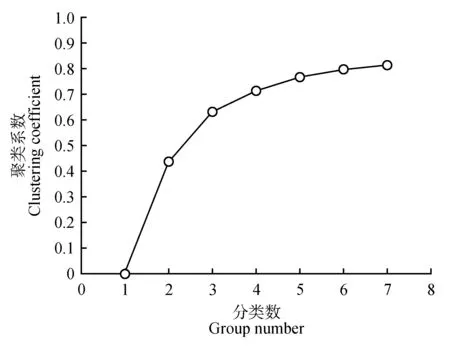
图1 糠椴天然居群聚类系数Fig.1 Clustering coefficient variation in T.mandshurica natural populations
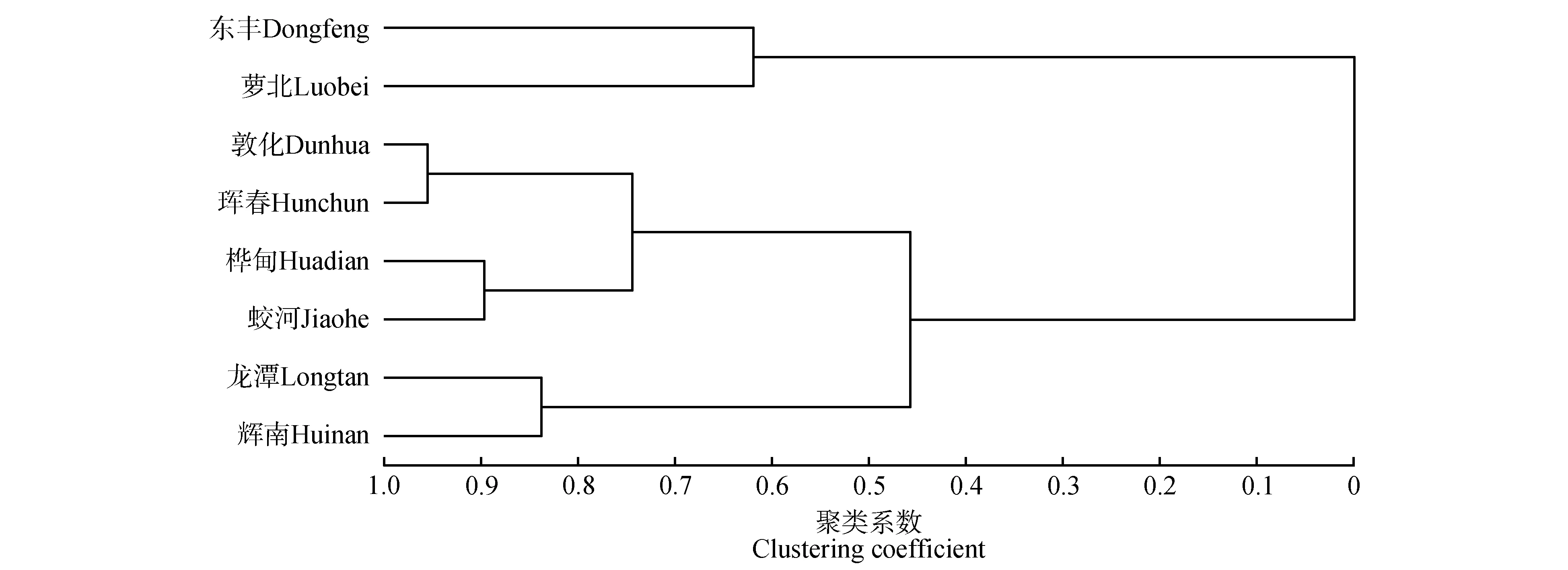
图2 糠椴天然居群聚类Fig.2 Dendrogram of T.mandshurica natural populations
3 讨论
3.1 糠椴居群的表型差异来源
天然居群的表型性状变异系数和表型分化系数可以有效反映表型性状的离散程度和变异来源[19]。糠椴叶片和果实性状在居群间和居群内的差异均达到极显著水平,各性状在居群间的方差分量百分比均大于居群内,表型分化系数介于0.83~0.95,说明居群间是糠椴表型性状差异的主要来源。这与严灵君等[11]对南京椴(Tilia miqueliana)天然居群的表型性状多样性及变异分析的结果一致,南京椴11 个表型性状居群间的方差分量百分比(43.50%)明显大于居群内(14.16%),各表型性状的居群间表型分化系数介于0.66~0.86;但汤诗杰等[20-21]基于ISSR 和RAPD 标记对南京椴天然居群的遗传变异进行分析,认为居群内变异是南京椴分化的主要来源。糠椴的表型性状不仅由因素决定,还受到不同居群的土壤、光照、水分和伴生树种的影响,具有一定的环境可塑性,是植物适应环境因子的一种表现。
3.2 糠椴居群的表型多样性
植物表型性状多样性是生物多样性研究的重要内容,有利于了解植物的分化和进化潜力,为优质种质的筛选、保护和利用提供理论依据[22-23]。Mishra 等[24]对番石榴(Psidium guajava)果实品质性状和生物活性的表型多样性进行了分析,发现番石榴的果实品质性状和生物活性均具有高度的表型多样性。本研究中,糠椴各表型性状的变异系数介于20.0%~39.9%,变异程度较大,说明糠椴表型性状存在丰富的变异;Shannon-Wiener指数介于1.262~1.443,表明糠椴的表型性状具有丰富的多样性。植物表型性状越丰富,表明植物的可选择性越高,越有可能产生较高的增益。
3.3 糠椴居群表型性状的综合排序
主成分分析中,特征向量绝对值较大的表型性状为反映植物表型性状差异的主要因子,也是植物多样性保护和目的性状选育研究中的主要观测性状[25]。Vidaković 等[26]分析了亚得里亚海东部海岸地区刺梨(Pyrus spinosa)的表型多样性,将刺梨的10 个叶片性状分为3 个主成分,分别反映了叶片大小、叶形和叶柄特征。本研究将糠椴的11 个表型性状分为4 个主成分,分别反映果实大小、苞果特征、叶片大小、果柄和果序柄大小,累计方差贡献率高达88.46%。说明主成分分析可将存在相关性的性状转换为不相关的主成分,进而反映植物的表型差异。
3.4 糠椴居群分类
植物天然居群保持较高的多样性水平有利于其居群的发展。在糠椴种质资源保护过程中,通过对表型性状的聚类分析,可以阐明不同居群表型性状的差异,进而依据育种目标筛选出适宜的育种材料[27-28]。聚类分析结果表明:当聚类系数为0.631时,可以将8个糠椴居群分为三类。其中,第Ⅰ类居群包括东丰和萝北,叶片较大;第Ⅱ类居群包括敦化、珲春、桦甸和蛟河,果柄和苞叶较大;第Ⅲ类居群包括龙潭和辉南,果实较大。糠椴三类居群并没有表现出明显的地理距离特征,在第Ⅰ类居群中,东丰与萝北的地理距离较远却聚为一类,这与亚得里亚海东部海岸地区刺梨[26]的研究结果一致,17 个天然居群没有遵循明确的地理或环境特征。造成这种现象的原因可能是由于糠椴天然居群之间没有明显的地理障碍,不同居群间的遗传物质可以通过以糠椴种子为食的动物进行传播。
4 结论
综上所述,糠椴天然居群的叶片和果实性状具有丰富的表型多样性,居群间是糠椴表型性状差异的主要来源。通过对表型性状的聚类分析对糠椴不同居群进行分类,可依据不同的育种目标筛选出适宜的育种材料,为糠椴核心种质资源挖掘、资源的保护与利用提供参考。然而,本研究选取的糠椴天然居群主要分布于吉林省范围内,具有一定的空间局限性,后续可进一步根据糠椴的分布区域扩大取样范围。与此同时,气候和土壤等环境因子对糠椴表型性状稳定性和环境可塑性的解析有待于进一步研究。
猜你喜欢
新疆农业科学(2023年6期)2023-07-13 03:53:58
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
湖南农业科学(2022年5期)2022-06-22 08:26:06
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
农业与技术(2021年5期)2021-03-31 12:26:16
山东农业工程学院学报(2019年3期)2019-05-11 07:41:50
草地学报(2018年5期)2018-11-07 02:25:00
现代园艺(2017年21期)2018-01-03 06:41:32
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
医学研究杂志(2015年5期)2015-06-10 06:43:26
