
不同修枝处理对10年生三倍体毛白杨生长和光合特性的影响
2023-11-13 09:23任天梦苏亚勋陈庆斌魏立颖纪若璇于夏新莉尹伟伦
植物研究 2023年6期
任天梦 鲍 雨 苏亚勋 陈庆斌 魏立颖 纪若璇于 笑 夏新莉 尹伟伦 刘 超*
(1.北京林业大学生物科学与技术学院,北京 100083; 2.京蓝北方园林(天津)有限公司,天津 300000)
杨树(Populus)是目前世界中纬度平原地区栽培面积最大、木材产量最高的速生用材树种之一,也是我国速生林主栽树种,具有速生、优质、高产、适应性强和易于更新等特点[1-3]。修枝是人工林经营培育中比较重要的措施之一,通过人为修除杨树下部枯枝和密集枝,合理调节干、枝、叶之间的物质分配,使树干圆满通直,有利于培育大径级用材[4-5]。修枝不仅可以改善树木的通直度和尖削度,还可以改善林分环境状况,对林分的稳定性和林木病虫害的防治也具有一定的积极作用[6]。
国内外对杨树修枝已有大量研究,主要涉及不同强度修枝对林木树高、胸径、材积等生长指标,以及净光合速率、蒸腾速率、气孔导度、瞬时水分利用效率等生理指标的影响,如:方升佐等[7]通过修枝降低了2 年生I-69 杨(Populus deltoides‘Lux’)的平均胸径和平均单株材积,但与对照相比,未达差异显著水平;陈森锟[8]通过修枝一定意义上提高了2 年生欧美107 杨(P.×euramericana‘74/76’)剩余叶片净光合速率和蒸腾速率,但对树木生长无明显促进作用;宋立志[9]通过重度修枝显著促进5 年生中辽1 号杨(P.×canadensis‘Zhongliao 1’)树高、胸径以及材积的生长;李广德[10]通过轻度修枝提高了8 年生三倍体毛白杨B301(P.tomentosa‘301’)叶片光合速率的同时显著促进了林木胸径和材积的生长。修枝处理能够显著影响低幼龄林的生长及生理活动,但以往研究集中在2~8年生杨树,对于10 a及以上的中高龄杨树,却鲜少有相关的修枝报道。
中高林龄杨树有利于大径材的培育,其已成为胶合板、纤维板等工业用材的重要加工原料,而大径材的培育所需年限较长,采伐年龄最低界限为10 a[11]。修枝可以提高林木材质、改善林木干形,但目前修枝技术对10 a 及以上林龄杨树的生长和光合特性的作用规律缺乏足够的研究。本研究以10 年生三倍体毛白杨(P.tomentosa‘Yixian Cizhu’)无性系B301 与S86 为研究对象,通过对不同强度修枝后3 年间林木生长及其光合生理特性的响应研究,探索符合毛白杨大径材生长的合理修枝技术,为集约化经营提供理论与实践依据。
1 材料与方法
1.1 研究地概况
试验地设在河北省威县林木良种繁育基地(37.04°N,114.30°E),地处华北平原南部,自然环境优越,地势平坦,气候四季分明,为暖温带大陆性半干旱季风气候,年平均气温13 ℃,年平均降水量584 mm,全年日照2 574.8 h。
1.2 试验设计
试验材料为三倍体毛白杨无性系B301和S86,于2007年造林,试验地毛白杨总面积为10 hm2,株行距4 m×3 m,南北走向,无间作。试验分3 个修枝强度(见图1),分别为轻度修枝(Q,修至树高1/3 处,约7 m)、重度修枝(Z,修至树高1/2 处,约10 m),并以不修枝作对照(CK)。试验地采用随机区组设计,每个处理设置3 个小区,试验林地共3×3 个小区,每个小区内设置8 行,毛白杨无性系B301 和S86 每2 行间隔分布,每行3~4 株,试验小区四周以泡桐(Paulowniasp.)或毛白杨1316 为保护行。于2017 年4 月集中修除指定高度下的枝条,并于2018、2019 年4 月修去修枝高度下新萌发的枝条,保证各处理的修枝强度。
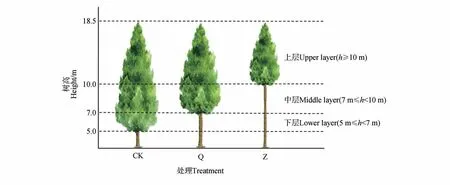
图1 2017年修枝处理示意图修枝前CK活枝下高为5.0 m,林分平均树高为18.5 mFig.1 Diagram of pruning treatment in 2017 Before pruning,the height under the CK live branches was 5.0 m,and the average stand height was 18.5 m
1.3 数据收集
1.3.1 生长指标的测量
在2017、2018、2019、2020 年春季4 月用激光测高测距仪(Vertex VL400,瑞典)和胸径尺测定所有毛白杨无性系植株修枝前、修枝1 a 后、修枝2 a 后和修枝3 a 后的树高、胸径等生长指标,根据公式V=0.513 40H0.828958D1.995375计算各修枝处理的平均单株材积[12]。为了消除林木个体优势可能造成的影响,采用树高、胸径、材积增长率分析生长指标
式中:ΔH表示树高增长率(%),H1为每年初所测树高(m),H2为次年初所测树高(m);ΔD表示胸径增长率(%),D1为每年初胸径(cm),D2为次年初胸径(cm);ΔV表示材积增长率(%),V1为每年初材积(m3),V2为次年初材积(m3)。
1.3.2 生理指标的测定
于2017、2018、2019 年5~9 月用LI-COR 便携式光合仪(Li-6400,美国)测定试验地毛白杨人工林1 个生长季的光合特性变化。按照修枝高度将植株分为上部(h≥10 m)、中部(7 m≤h<10 m)、下部(5 m≤h<7 m,10 a 龄苗木活枝下高4 m 左右) 3层,CK 选取上部、中部、下部枝条,Q 选取上部、中部枝条,Z 选取上部枝条,采用搭建脚手架对向阳枝条进行活体测定。测定时选择晴朗天气,采用自然光源,CO2摩尔分数设定为400 μmol·mol-1,各处理选取3 株样木,从08:00—18:00每2 h每株各部位随机选取3个枝条,测定中部成熟叶片(3~5个)的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等指标,并计算瞬时水分利用效率(WUE=Pn/Tr)。
1.4 数据分析
采用Excel 2019 软件对三倍体毛白杨不同修枝后的生长和光合特性变化等数据进行处理,利用SPSS 26.0 统计分析软件进行差异显著性检验(LSD 法,P<0.05)和多因素方差分析,比较三倍体毛白杨生长及光合特性在不同修枝强度、年份、品系以及冠层之间的差异。
2 结果与分析
2.1 修枝对毛白杨生长的影响
对试验地修枝前后树高、胸径、材积进行测量计算,得到不同强度修枝毛白杨生长变化情况(见图2)。多因素方差分析显示修枝处理对毛白杨树高、胸径增长率有一定的影响(P<0.05,见表1),但进一步分析显示(见图2),仅2018 年B301 杨胸径增长率未修枝显著大于重度修枝、S86杨未修枝显著大于修枝处理,以及2019年B301杨树高增长率轻度修枝显著大于其他处理;其余处理在修枝后的3 年间,两品系三倍体毛白杨树高、胸径以及材积的增长率在各修枝强度下均无显著性差异,甚至修枝处理对各生长指标的增长率在修枝前2 年间表现出不同程度的抑制。综合可知,修枝处理对于10年生杨树生长无明显促进作用。
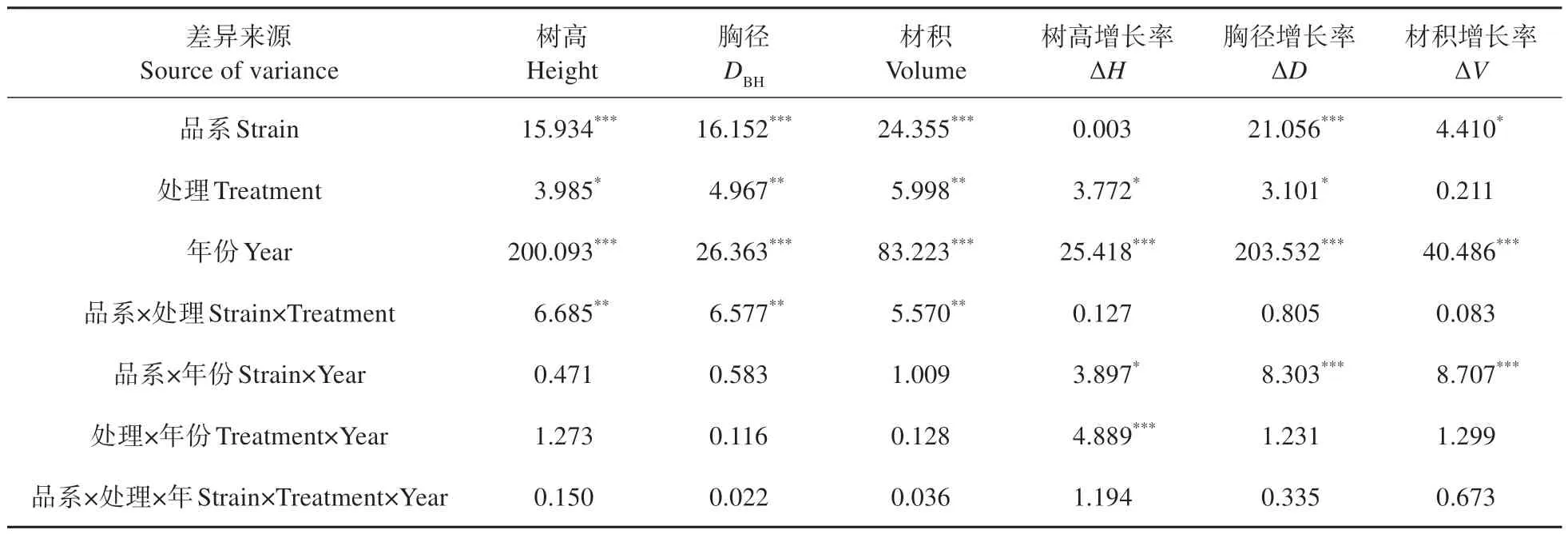
表1 2017—2019年修枝处理对毛白杨生长的多因素方差分析(F值)Table 1 Multivariate variance analysis of pruning treatment on growth of P.tomentosa from 2017—2019(F value)
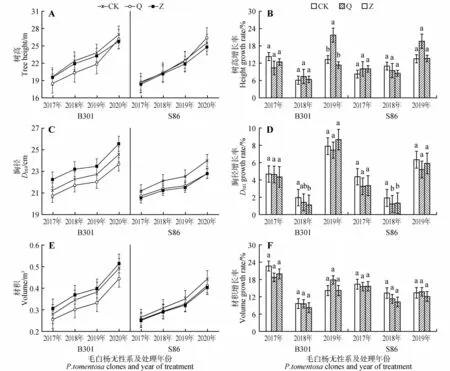
图2 不同修枝对10年生三倍体毛白杨生长的影响A、C、E分别为三倍体毛白杨树高、胸径、材积的年际变化;B、D、F分布为各年份毛白杨树高、胸径、材积增长率;不同小写字母表示修枝处理间差异显著(P<0.05);下同Fig.2 Effects of different pruning on the growth of 10-year-old triploid P.tomentosa A,C,E were the interannual changes of height,DBH and volume of triploid P.tomentosa respectively;B,D,F were the ΔH,ΔD and ΔV of P.tomentosa in each year respectively;Different lowercase letters indicated significant difference between pruning treatment(sP<0.05);the same as below
多因素方差分析显示(见表1),三倍体毛白杨不同品系间胸径、材积增长率存在显著性差异(P<0.05),树高增长率无差异(P>0.05)。修枝后不同年份毛白杨树高、胸径和材积增长率差异极显著(P<0.001),修枝处理和年份的交互作用对树高增长率有显著影响(P<0.01),品系和年份的交互作用对树高、胸径、材积增长率均有显著影响(P<0.05)。进一步多重比较分析显示(见表2),不同修枝强度下,树高、材积增长率在三倍体毛白杨B301 和S86 两品系间均无显著差异;但在轻度和重度修枝处理下,毛白杨B301 胸径增长率显著高于S86。修枝后不同年份毛白杨增长率比较显示,2018 年两品系毛白杨胸径、材积增长率低于其他年份。2017 和2019 年毛白杨B301 树高、胸径、材积增长率要显著高于S86,但在2018年S86的树高增长率、材积增长率要显著高于B301。2018 年2个品系的毛白杨生长率都有一定程度的降低,但B301杨在2018年的降幅较大。
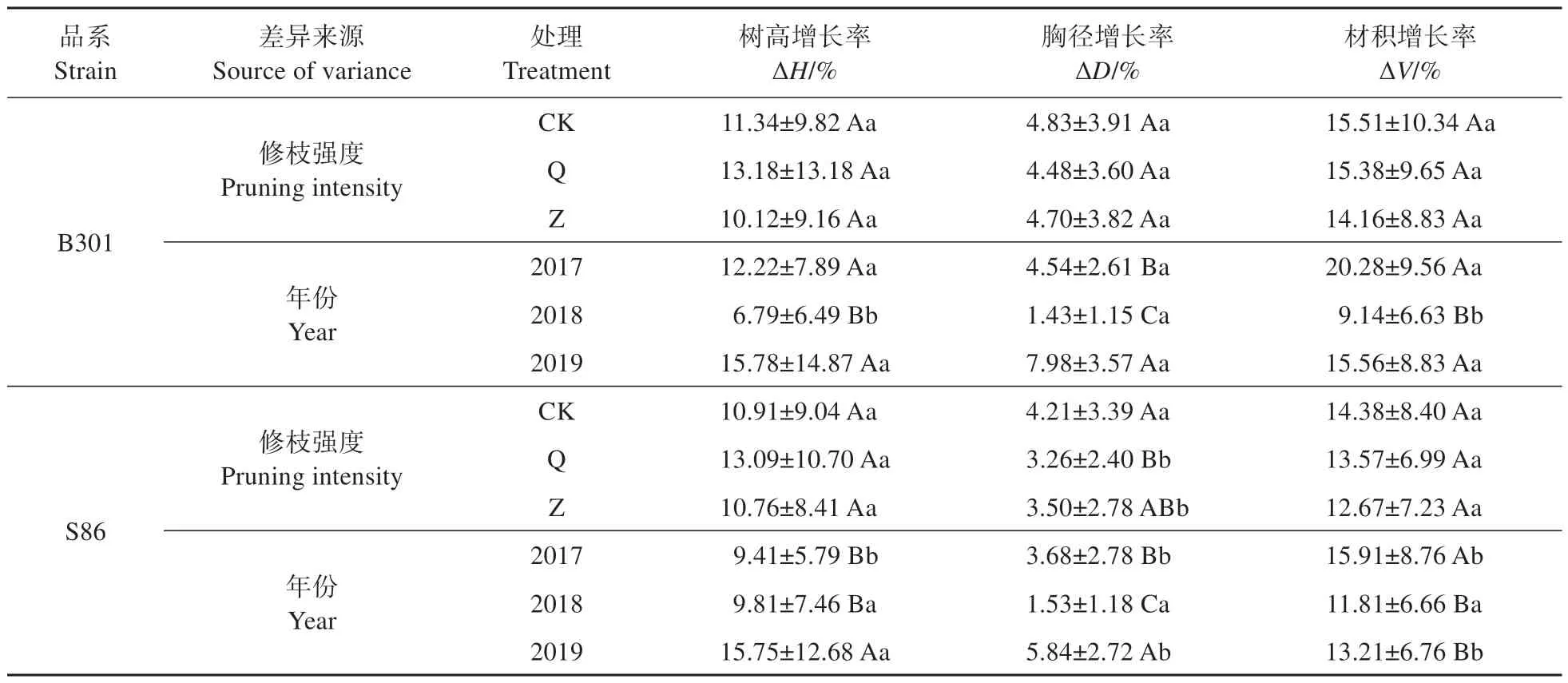
表2 不同修枝年份和品系间三倍体毛白杨树高、胸径、材积增长率多重比较Table 2 Multiple comparisons of ΔH,ΔD and ΔV of triploid P. tomentosa among different pruning years and strains
2.2 修枝对毛白杨光合生理的影响
2.2.1 对光合日变化的影响
分析B301和S86杨单叶在不同修枝处理下各月份净光合速率日变化折线图发现(见图3),不同修枝强度净光合速率、蒸腾速率和气孔导度日变化规律基本呈现双峰曲线,在10:00左右达到一天中的第1个峰值,12:00左右出现“午休现象”,14:00左右达到第2个峰值。可以看出,重度修枝在各月份日变化峰值点高于其他处理,且在中午出现低谷时仍能保持较高光合速率,而轻度修枝与未修枝之间差异不大。此外,从表3 可知,毛白杨两品系B301和S86的冠层上部叶片均表现出较高净光合速率和蒸腾速率,且显著高于冠层中部叶片(B301)和冠层下部叶片(S86)。
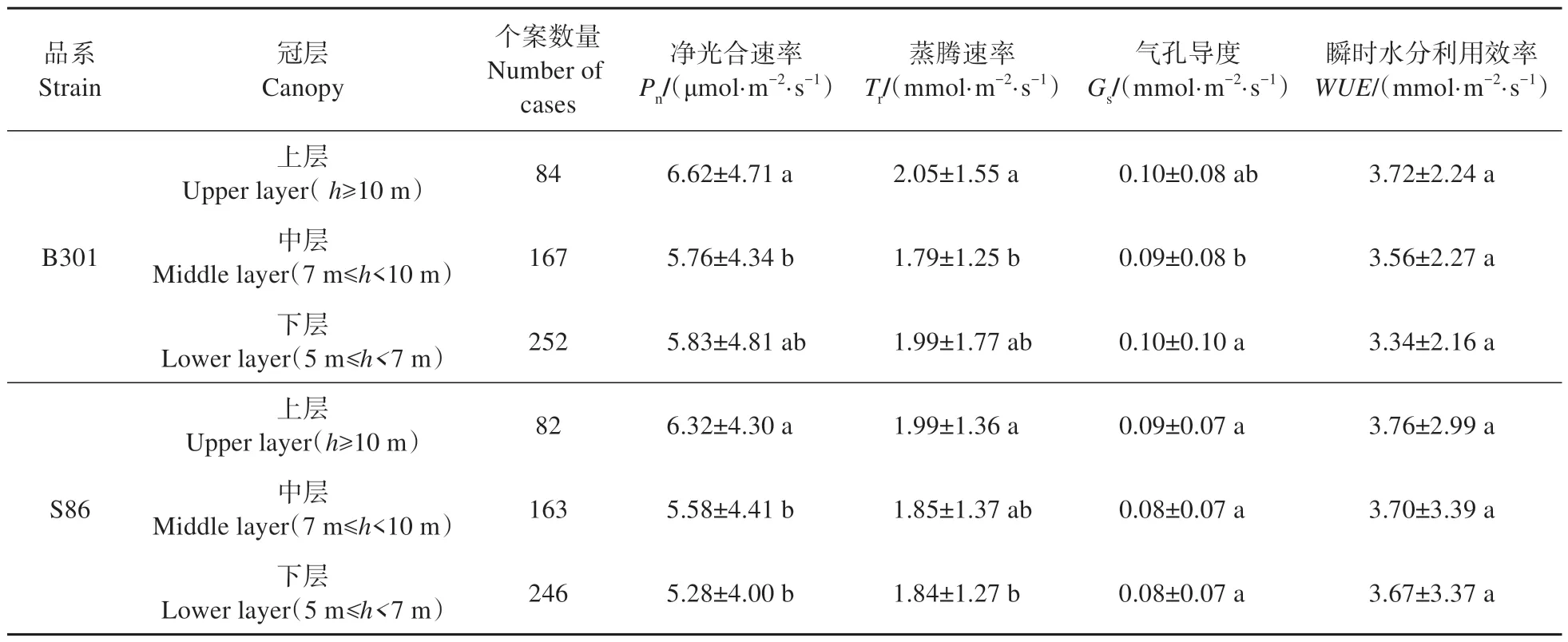
表3 毛白杨不同冠层叶片光合特性比较Table 3 Comparison of photosynthetic characteristics of different canopies of P. tomentosa
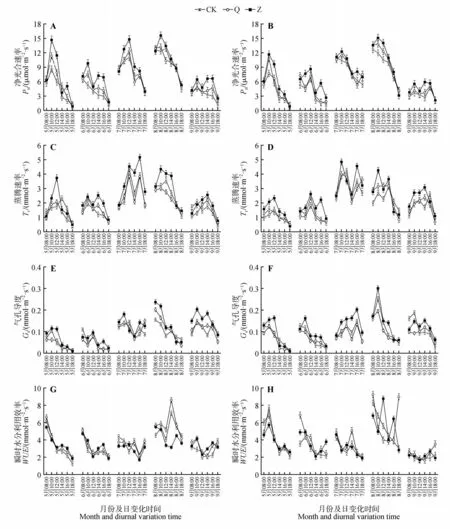
图3 不同修枝强度对10年生毛白杨光合日变化的影响A、C、E、G为三倍体毛白杨B301无性系;B、D、F、H为三倍体毛白杨S86无性系;下同Fig.3 Effects of different pruning on diurnal variation of photosynthesis of 10-year-old P.tomentosa A,C,E and G were triploid Populus tomentosa 301;B,D,F and H were triploid P.tomentosa S86;the same as below
2.2.2 修枝对毛白杨光合特性的影响
分析不同修枝处理B301和S86在生长季的生理指标折线图发现(见图4),净光合速率和蒸腾速率月变化趋势基本呈现单峰曲线,均在7 月或8 月达到一年中的峰值,生长季初5 月、生长季末9 月较低,且在6 月普遍出现低谷值;气孔导度在不同年份表现出不同的月变化规律,在2017 和2019 年逐渐上升,B301在2018年逐渐下降,而S86呈现单峰曲线。
多因素方差分析显示(见表4~5),修枝处理、不同年份、月份对毛白杨2个品系的光合特性均有显著影响(P<0.05),但两品系间差异不显著(P>0.05)。不同修枝处理毛白杨叶片净光合速率、蒸腾速率、气孔导度在3年间表现为重度修枝显著高于其他处理,而轻度修枝与未修枝差异不大,即重度修枝>轻度修枝≈未修枝。修枝处理对WUE无显著影响(P>0.05),瞬时水分利用效率在不同修枝间无明显变化规律。三倍体毛白杨净光合速率、蒸腾速率、气孔导度在修枝后第1 年(2017 年)显著低于修枝后第2 年(2018 年)和第3 年(2019年),后两者差异不大。在品系、修枝处理、年份和月份等因素的交互作用中,仅年份×月份对毛白杨光合生理特性有显著影响(P<0.05),其余均无显著影响(P>0.05)。
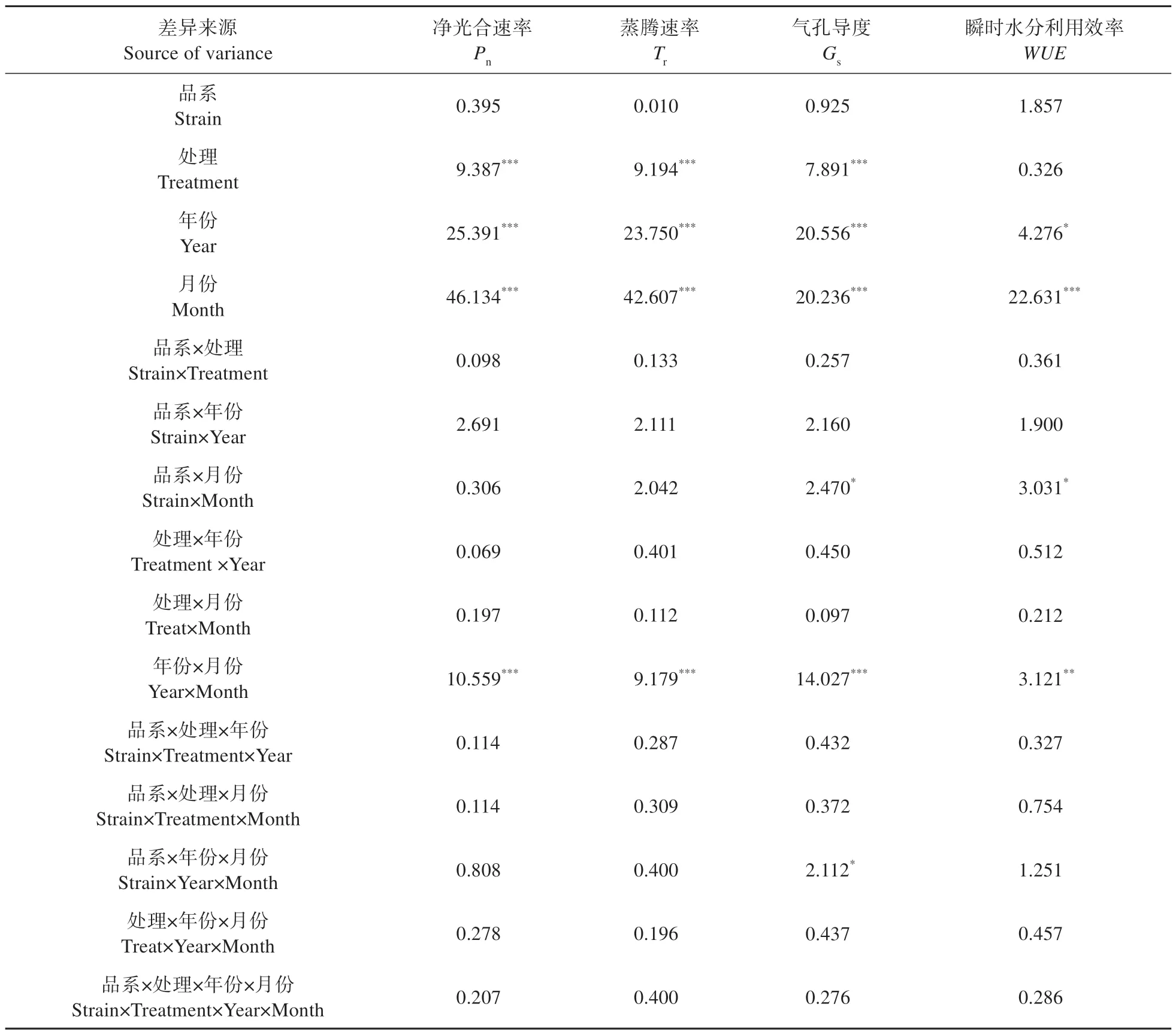
表4 2017—2019年修枝处理对毛白杨光合特性的多因素方差分析(F值)Table 4 Multi-factor variance analysis of photosynthetic characteristics of P.tomentosa under pruning treatment from 2017-2019(F value)
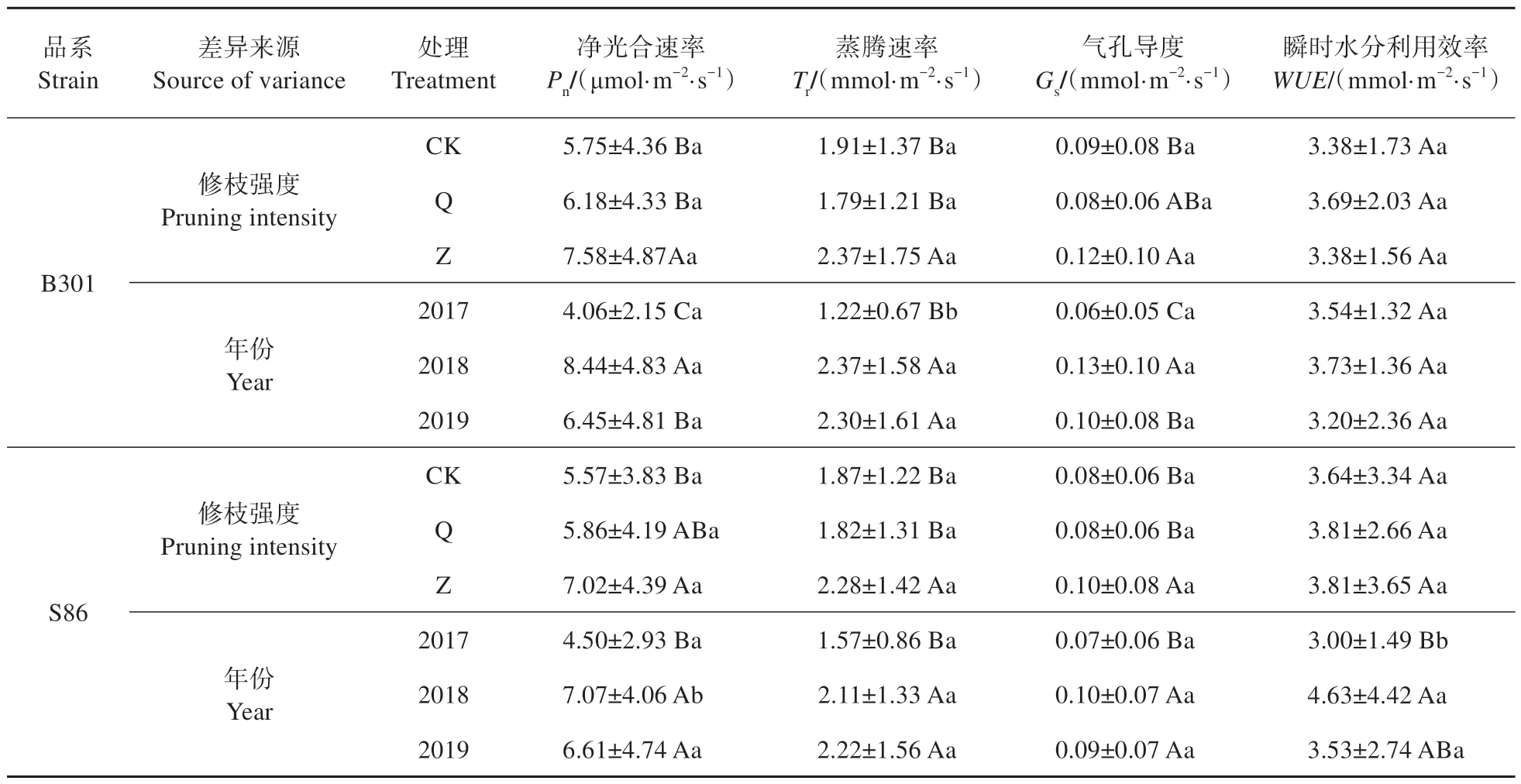
表5 不同修枝处理、年份和品系间三倍体毛白杨光合特性多重比较Table 5 Multiple comparisons of photosynthetic characteristics of triploid P. tomentosa among different pruning treatments,years and strains
3 讨论
本研究以10 年生三倍体毛白杨无性系B301和S86 为对象,发现不同修枝处理后,重度修枝处理(修去树冠下部1/2 枝条)增强了10 年生毛白杨叶片光合能力,而轻度修枝(修去树冠下部1/3 枝条)与未修枝差异不大。尚富华等[13]认为毛白杨无性系中上部叶片净光合速率较高,而下部叶片净光合速率处于比较低的水平,本试验数据与此相似。重度修枝修掉毛白杨10 m以下光合能力较差的消耗枝,显著增强了剩余叶片的光合能力、气孔导度及蒸腾作用,促进了林木有机物的积累并维持相对稳定的保水能力。而轻度修枝仅修去7 m 以下的下部叶片,林分仍处于郁闭状态,林内通风和光照条件没有得到明显改善,相较于重度修枝,对叶片光合的整体促进作用不明显。此外,源汇关系学说认为光合作用会受到碳水化合物源汇比率改变的影响,由于重度修枝修除大量叶片,源相对于汇变小,树木会通过提高剩余叶片光合速率进而促进源的供给能力[14]。试验还表明7—8月份是毛白杨生长的黄金期,净光合速率在7、8月远高于5、6 和9 月,尤其在6 月普遍出现不同程度的低谷值,此时光合能力易受外界影响,进行林木抚育工作时应当注意。
大量林木研究已证明合适强度的修枝处理有利于增强修枝后剩余叶片的光合能力和物质合成能力,但对林木生长影响的认识尚有分歧[10,13,15-16]。一种观点认为修枝能够增强剩余叶片光合能力的同时显著促进林木的生长,如李广德等[10]通过轻度修枝提高了8年生三倍体毛白杨B301叶片净光合速率,进而显著促进林木胸径和材积的生长;尚富华等[13]通过轻度修枝提高了4 年生毛白杨无性系1316 上部叶片净光合速率,同时促进了树高的生长。另一种观点认为修枝虽然提高了树冠上部叶片净光合速率,但短时间内剪除大量树木枝叶,极大减少了整株树木光合产物的总量,尽管树木存在补偿机制[17],修枝后增加的光合能力抵消了树木大量枝叶缺失导致的不利影响,但如果超过了补偿作用的阈值,必然影响林木的生长,甚至降低树木的增长。如陈森锟等[16]认为修枝虽然提高了2 年生欧美107 杨剩余叶片净光合速率,但降低了胸径的生长率,对树高生长影响不显著;Li 等[15]研究表明修枝显著提高了8 年生杉木(Cunninghamia lanceolata)剩余叶片的净光合速率,但对树高和胸径的生长无影响。目前尚缺乏足够的证据来证实修枝处理对林木生长的正向促进作用。
本研究结果与第二种观点相符,修枝处理在提高树冠上部叶片净光合速率的同时,对10 年生毛白杨的生长无明显促进作用,2个品系三倍体毛白杨的树高、胸径、材积增长率总体上在不同处理间没有显著性差异,甚至修枝前2年间各修枝处理对毛白杨的增长率表现出一定的抑制,即使这种抑制不明显。修枝虽然有利于光合速率的提升,但同时也促进了蒸腾耗水(Tr增加),增加了植物水分耗散,加剧了物质的损耗,不同修枝处理间水分利用效率(WUE)没有明显差异[18],而WUE越大表明固定单位数量的CO2所需水分越少,越有利于树木的生长。国内外大量研究也表明,林木修枝后短时间内仍处于生长缓慢和生长恢复阶段,随着修枝伤口愈合及林木生长,修枝的作用才会逐渐显现出来。此外,本研究的三倍体毛白杨修枝时树龄为10 年生,已处于杨树大径材培育采伐年龄最低界限。Zhang 等[19]对12 年生杨树(Populus alba×P.talassica)修枝提高了地上器官光合产物的含量,但重度修剪和不修剪处理的平均树高和胸径生长量差异不显著。同样,刘可欣等[20]研究发现不同强度修枝处理的15、20 年生水曲柳平均胸径生长与对照无显著差异。即修枝处理对10年以上的中高龄林杨树的生长的促进作用有限。
值得注意的是,2018 年各修枝处理毛白杨叶片净光合速率相对其他年份较高,而林木胸径、材积生长率整体偏低,这与2018 年生长季相对于2017 年和2019 年较干旱有关,轻度干旱的补偿机制能促进植物的光合作用,但因为环境变化引起的植物应激反应会使植物的生长受抑制[21]。此外,B301品系的毛白杨易受环境变化的影响,轻度和重度修枝处理对B301 的胸径增长率提升要显著高于S86。在水分较为充裕的年份(2017 和2019 年),B301 的株高、胸径、材积增长率显著较高;而在相对干旱的2018年,B301相较S86有较大的生长降幅,树高、材积增长率要显著低于S86。
本研究发现修去树冠下部1/2 枝条处理在一定程度上改善了10年生三倍体毛白杨光合生理状况,促进了有机物质的积累和抗旱能力,但修枝并没有引起树高、胸径和材积生长率的显著变化,对杨树木材的生长无明显影响。同时应该看到,由于林木生长周期较长,短时间内难以精确全面地评价修枝对树木生长和光合特性的影响,因此修枝对中高龄林生长的促进作用还有待进一步论证和研究。
猜你喜欢
中国海洋大学学报(自然科学版)(2023年2期)2023-02-21
科学养鱼(2021年11期)2022-01-11
甘肃林业科技(2021年3期)2021-03-30
新农民(2020年21期)2020-12-08
东坡赤壁诗词(2019年3期)2019-07-05
食品与生活(2019年12期)2019-05-15
生命科学研究(2018年1期)2018-05-29
东坡赤壁诗词(2018年1期)2018-03-31
山西林业科技(2018年4期)2018-03-19
河南林业科技(2017年1期)2017-06-10
