
昆虫触角感器类型及其功能研究进展
2023-11-09 12:15兰晓娜向姗姗
环境昆虫学报 2023年5期
兰晓娜,向姗姗,朱 慧
(东北师范大学生命科学学院/植被生态科学教育部重点实验室/吉林松嫩草地生态系统国家野外科学观测研究站/吉林省动物资源利用与保护重点实验室,长春 130024)
在长期进化过程中,昆虫为适应食物和环境变化形成了独特的形态特征和生活习性。从形态特征角度看,身体的每一结构都被认为执行特定的功能。昆虫头部着生的一对形态各异的触角,是其接受和传递信息的重要结构,不仅能够感知外界环境的化学气味、湿度和温度等,也在寻找寄主、交配和防御等方面发挥关键性作用。触角的这些功能主要是通过触角上固着的不同类型的感器实现,环境中的信息通过感器中相关蛋白传入神经系统,进一步传递给脑,使昆虫做出相应的行为反应。因此,深入认识昆虫触角感器的种类和功能对于探究昆虫与外界环境的相互交流有着重要意义。Steinbrecht借助于光学和扫描电子显微镜对家蚕Bombyxmori的触角形态学进行了详细研究,并报道了毛形感器和锥形感器具有嗅觉功能(Schneider, 1964)。当前,研究学者将扫描电子显微镜和透射电子显微镜结合使用,对昆虫触角感器的研究已深入到超微结构。由于昆虫触角感器在识别环境、寄主、配偶等方面极为重要,国内外研究学者对各类群昆虫的触角感器分别进行了广泛研究,并已取得显著进展,特别针对直翅目、鞘翅目、鳞翅目、同翅目、半翅目、双翅目、膜翅目等昆虫的触角感器类型开展大量研究。本文针对已有研究中的昆虫触角感器种类进行系统性总结,探究感器类型在种内种间的特异性,并对不同类型的触角感器可能具有的生物功能进行梳理,有助于理解昆虫触角感器在长期演化过程中对环境的适应性变化,旨在为深入研究揭示昆虫与寄主、天敌和环境的相互关系提供重要的理论基础。
1 昆虫触角感器类型
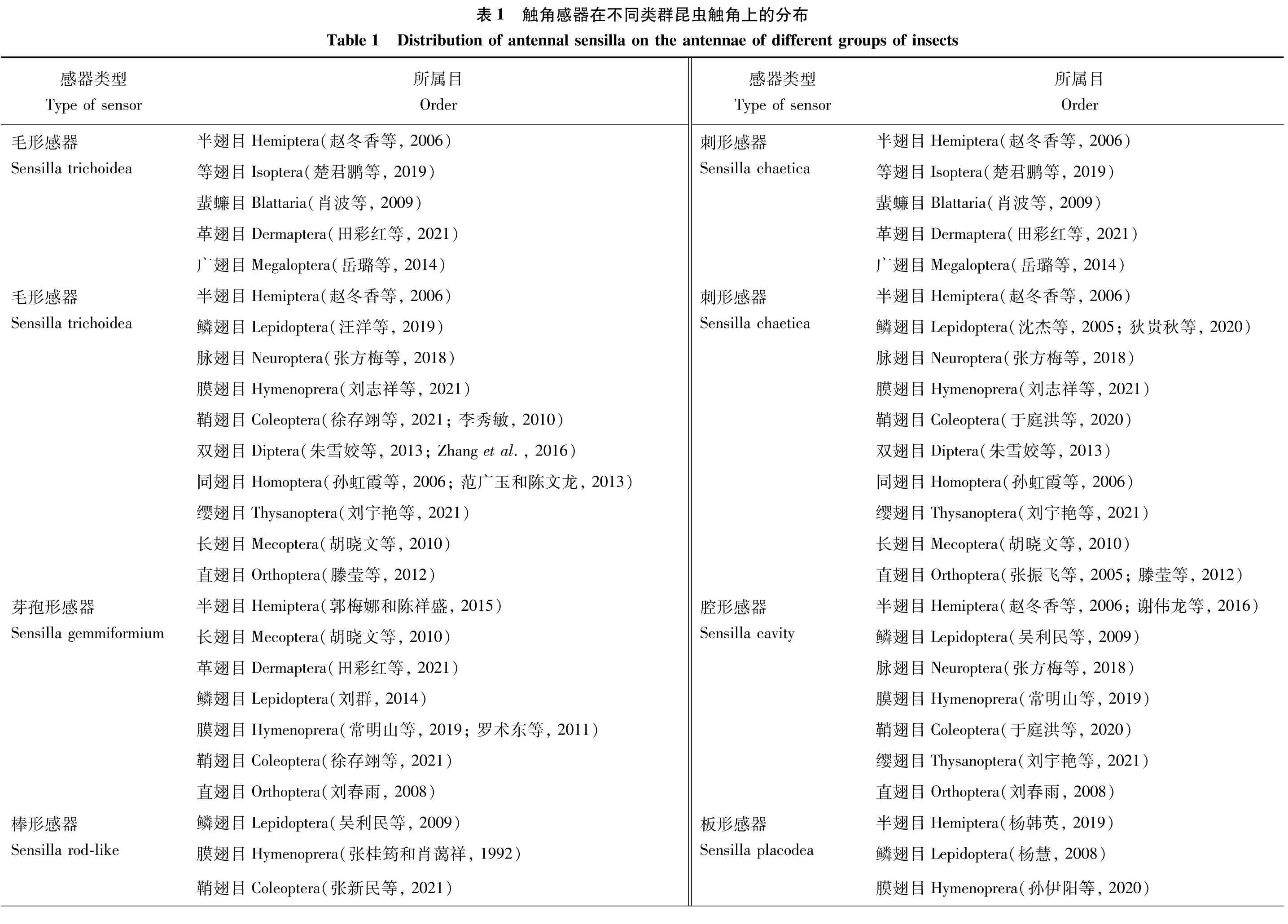
昆虫触角感器是由昆虫的体壁特化而来,由毛原细胞、神经细胞和各种鞘细胞组成。在早期关于昆虫触角感器的研究中,Snodgrass提出了9种基本类型,接着Schneider在基本类型的基础上增加了2~3种类型,并且针对基本的触角感器类型详细区分了各种亚型(Zacharuk, 1980)。直到目前,随着研究的昆虫种类逐渐增多,不断有新的触角感器类型被发现。本文汇集已有研究,确定目前已发现昆虫触角感器类型61种,每种感器都有特定的形态特征,并进一步总结昆虫触角感器类型在不同类群昆虫触角上的分布情况(表1)。
1.1 毛形感器(Sensilla trichoidea)
呈长毛状,着生在表皮凸上成簇分布,直立或弯曲,基部粗大,由底部向顶端逐渐变细,某些昆虫的毛形感器在顶端有一念珠状突起(袁轲等, 2020)。根据外部形态的不同,将毛形感器分为毛形感器Ⅰ型、毛形感器Ⅱ型、毛形感器Ⅲ型、毛形感器Ⅳ型和毛形感器Ⅴ型(尹宁娜等, 2020)(图1)。毛形感器在目前研究的昆虫类群中都有分布,包括鳞翅目、膜翅目、双翅目、鞘翅目、半翅目、等翅目、蜚蠊目、革翅目、广翅目、脉翅目、同翅目、缨翅目、长翅目和直翅目类群的昆虫触角上都发现有毛形感器的存在(表1),可以初步认为是分布最广的触角感器类型之一。
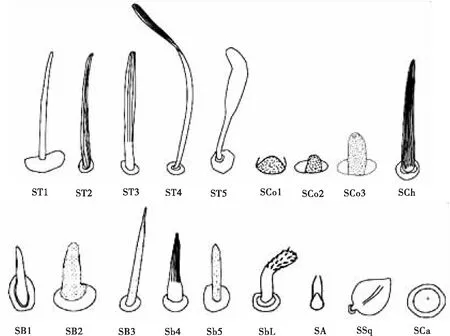
图1 八种昆虫触角感器类型示意图(Nowiń
1.2 刺形感器(Sensilla chaetica)
呈针刺状的刚毛形态,感器表面有明显的纵脊,部分类型表面光滑,基部与触角表面的连接处有一凹槽,由底部向顶端逐渐变细,顶端尖锐(图1)。大部分感器与触角表面的夹角在60~80°之间,少部分直立于触角表皮。截至目前,刺形感器已发现有4~5种亚型:刺形感器Ⅰ型、刺形感器Ⅱ型、刺形感器Ⅲ型、刺形感器Ⅳ型和刺形感器Ⅴ型(曹延昆, 2017)。刺形感器也是分布最广的触角感器之一,目前,在已有研究的半翅目、等翅目、蜚蠊目、革翅目、广翅目、鳞翅目、脉翅目、膜翅目、双翅目、鞘翅目、同翅目、缨翅目、长翅目和直翅目昆虫触角上都有分布(表1)。
1.3 锥形感器(Sensilla basiconica)
呈短锥状,生长在突出于触角表面的圆形穴内,整体呈乳突状或小钉状,表面光滑无纹路,基部膨大,并逐渐变细,顶端有数量较多的微小孔,在触角表面直立,几乎与触角轴成直角,壁薄,梯度酒精清洗过程中容易断裂。根据锥形感器外部形态的不同,将其分为锥形感器Ⅰ型、锥形感器Ⅱ型、锥形感器Ⅲ型、锥形感器Ⅳ型和锥形感器Ⅴ型五种亚型(于庭洪等, 2020)(图1)。该类感器分布较广,包括半翅目、蜚蠊目、广翅目、鳞翅目、脉翅目、膜翅目、鞘翅目、双翅目、同翅目、缨翅目、长翅目和直翅目中的昆虫触角上均有发现(表1)。
1.4 腔锥形感器(Sensilla coeloconica)
1.5 栓锥形感器(Sensilla styloconica)
与其他类型的感器在形态上截然不同,栓锥形感器有一个球形突起,其上分布着纵形沟槽,有研究证实这些沟槽是由大量的微孔组成,其基部位于一个突出的圆台状基窝中,端部钝尖。分为两种亚型:栓锥形感器Ⅰ型和栓锥形感器Ⅱ型(徐伟等, 2020)。栓锥形感器分布在革翅目、鳞翅目、膜翅目、鞘翅目、双翅目和直翅目的部分昆虫触角上(表1)。
1.6 鳞形感器(Sensilla squamiformia)
形状类似剑鞘,表面具有纵脊,云南锦斑蛾Achelurayunnanensis的鳞形感器的两条纵脊间具横纹,两者形成网状结构(李根层等, 2020)。与普通鳞片相似,但比一般鳞片狭长且端部较尖,基部有臼状窝(图1)。分为两种亚型:鳞形感器Ⅰ型和鳞形感器Ⅱ型(汪洋等, 2019; 狄贵秋等, 2020)。据现有的触角感器研究,在6个目的昆虫触角上发现鳞形感器,分别为半翅目、鳞翅目、膜翅目、鞘翅目、双翅目和同翅目(表1)。
1.7 波氏鬃毛(Böhm bristles)
呈短毛状,直立,较细短,表面光滑无孔无纵纹,基部具不明显凹陷,顶部圆钝。像一根根小刺成簇地分布在触角的柄节和梗节的节间周围,侧面和腹面分布较多,呈灰白色。分为两种亚型:波氏鬃毛I型(汪洋等, 2019; 徐伟等, 2020)和波氏鬃毛Ⅱ型(李根层等, 2020)。波氏鬃毛分布非常广泛,在大部分类群的昆虫触角上都发现到了这类感器,包括半翅目、长翅目、等翅目、蜚蠊目、鳞翅目、脉翅目、膜翅目、鞘翅目、双翅目、同翅目、缨翅目、直翅目和革翅目(表1)。
1.8 板形感器(Sensilla placodea)
为椭圆或纵长形的板状结构,着生在形状相同的穴窝内,其长轴平行于触角长轴,没有突起,一般覆于触角表面,表面有许多辐射状的沟壑从盘中央延伸开来,沿沟壑密布小孔。从扫描电镜结果来看,板型感器分2种类型,板形感器I型为细长板形,长度与直径的比例大;板形感器II型的长度与直径的比例较小,看上去较粗胖(田小霞, 2021)。在半翅目、鳞翅目、膜翅目和鞘翅目昆虫触角上都有板形感器分布(表1),但不同目中存在板型感器的物种之间数量差异较大。
1.9 钟形感器(Sensilla campaniformia)
根据其外部形态的不同,分为钟形感器Ⅰ型和钟形感器Ⅱ型。钟形感器I型由表皮凹陷形成,表面光滑,顶部具有一小孔;钟形感器II型呈半球状,是体壁凹陷的圆形小穴内伸出的一个帽状结构,表面光滑,周围有光滑清晰的厚壁边缘(徐伟等, 2020),两种亚型的外形有相似之处(图1)。钟形感器分布较广,主要存在于半翅目、等翅目、蜚蠊目、鳞翅目、脉翅目、膜翅目、鞘翅目、双翅目、同翅目、缨翅目、长翅目和直翅目中(表1)。
1.10 舌形片状感器(Sensilla ligulate flaky)
呈舌形,薄片状,背腹面较宽,整体稍微扭曲,与触角表面大体呈45~60°角,表面有数条较深的凹槽,具基窝,端部稍弯向触角腹面,末端呈现圆弧形的弯曲(狄贵秋等, 2020)。目前仅在鳞翅目、脉翅目和膜翅目昆虫触角上有发现到了舌形片状感器(表1)。
1.11 腔形感器(Sensilla cavity)
表皮内陷形成的空腔,内部无锥体,触角表面呈孔状深陷(常明山等, 2019)。腔形感器在半翅目、鳞翅目、脉翅目、膜翅目、鞘翅目、缨翅目和直翅目昆虫触角上有分布(表1)。
1.12 刷形感器(Sensilla brush)
外观形态呈刷状(图1)。目前共发现2种刷形感器,刷形感器Ⅰ型从中下部开始有分叉,但分叉均统一朝向触角方向,与触角表面完全平行,几乎与触角表皮呈90°,有基窝,表面光滑;刷形感器Ⅱ型与刷形感器Ⅰ型较相似,但基部比刷形感器Ⅰ型多一簇感器,类似附属物,像一个小型刷形感器,与触角表皮呈45°角(段云博等, 2020)。截止目前这种感器类型仅在鞘翅目的一种昆虫触角上被发现(表1)。
1.13 簇状感器(Sensilla cluster)
有两种亚型,分别为簇状感器Ⅰ型和簇状感器Ⅱ型。簇状感器Ⅰ型形似毛笔头,短小较粗,有基窝,几乎垂直于触角表皮,中下部开始分叉,类似刷形感器,但分叉更多;簇状感器Ⅱ型形似火山爆发,只在表面有短分叉,且密集而短小,形似触角上的纹路,无基窝,几乎紧贴触角表皮着生。簇状感器仅在鞘翅目中一种昆虫触角上有发现(段云博等, 2020)(表1)。
1.14 芽孢形感器(Sensilla gemmiformium)
其在触角上的排布形态像植物种子萌发的芽,外壁光滑,数量较多,着生于触角表皮鳞片状凸起的边缘,一片鳞片上的多个芽孢形感器连成锯齿状,其中单个感器外形短小直立,顶端钝圆(徐存翊等, 2021)。在半翅目、长翅目、革翅目、鳞翅目、膜翅目、鞘翅目和直翅目中都有芽孢形感器的存在(表1)。
1.15 叉形感器(Sensilla furcatea)
外壁具纵沟,着生于臼状窝内,基部较宽,顶端有叉形分支。根据长度及分叉形状将其分为2种亚型:叉形感器Ⅰ型(段云博等, 2020)和叉形感器Ⅱ型(徐存翊等, 2021)。叉形感器仅在半翅目、鳞翅目和鞘翅目3个目的昆虫触角上有分布(表1)。
1.16 耳形感器(Sensilla auricillica)
由两侧向内卷,外形呈现禾本科植物类似的卷心叶片状,具有耳状凹槽,常着生于触角附枝基窝中,与触角表面夹角小,近乎平行,端部钝圆(李根层等, 2020)。在半翅目、鳞翅目、脉翅目、膜翅目、鞘翅目、双翅目、同翅目和缨翅目昆虫触角上有分布(表1)。
1.17 指形感器(Sensilla finger)
形似手指,粗细较均匀,一般不弯曲,表面光滑,无基窝。斑鞘豆叶甲Colposcelissignata中的指形感器具明显较深的漏斗形小孔,基部有延伸到圆台上的明显树状突起(徐伟等, 2020)。在鳞翅目、膜翅目和鞘翅目昆虫触角上有该类感器的分布(表1)。
1.18 乳状感器(Sensilla mammilliformia)
常细分为3种亚型:乳状感器Ⅰ型着生于基窝内,顶端膨大呈球状,形似乳头状突起;乳状感器Ⅱ型无基窝,末端较钝,呈锥状;乳状感器Ⅲ型也处于基窝内,但基窝大而深,有圆环状纹路,整体呈乳头状(孙伊阳等, 2020)。乳状感器在半翅目、革翅目、鳞翅目、膜翅目、鞘翅目、双翅目、同翅目和缨翅目昆虫触角上有分布(表1)。
1.19 柱形感器(Sensilla cylindrical)
呈柱形,表面光滑,着生于臼状窝内,基部到端部的粗细基本一致,顶端钝圆,直立于触角表面(岳璐等, 2014)。这类感器在广翅目、鳞翅目、膜翅目、鞘翅目、双翅目和同翅目中都有发现(表1)。
1.20 坛形感器(Sensilla ampullacea)
又称瓶形感器,生于表皮凹槽内的锥状体,与腔锥形感器形状非常相似,但表皮上的凹陷更深,并且通常表皮上的开口更窄(马瑞燕和杜家纬, 2000; 余海忠, 2007)(图1)。研究发现,坛形感器在革翅目、鳞翅目、膜翅目、鞘翅目和双翅目这4个目的昆虫触角上均有分布(表1)。
1.21 缘感器(Sensilla margin)
呈细小的毛状结构,常出现在鞭节相连处,一般是在鞭节的第一节最前端与梗节相连处密集分布的一种小型毛状结构,末端尖而锐(朱雪姣等, 2013)。目前仅在膜翅目和双翅目昆虫触角上发现具有缘感器(表1)。
1.22 槽钉形感器(Sensilla grooved peg)
呈钉状突起,表面光滑,顶端圆钝。着生于圆形隆起的凹窝内,长度明显短于锥形感器(刘晓梅等, 2015)。槽钉形感器仅在脉翅目和鞘翅目昆虫触角上具有(表1)。
1.23 毛板形感器(Sensilla hair plate)
短小直立,表面光滑,顶端较钝,基部深深插入触角表面的凹窝中(刘晓梅等, 2015)。在鳞翅目和鞘翅目昆虫中都发现了毛板形感器的存在(表1)。
1.24 多孔板形感器(Sensilla multiporous placoid)
形似板状,明显隆起于触角表皮。从基部到端部宽度基本一致,端部钝圆,顶端与触角主轴分离(赵健等, 2015)。目前发现多孔板形感器只在半翅目和膜翅目中存在(表1)。
1.25 纽扣形感器(Sensilla button-shaped)
形似纽扣,圆形,主要着生在触角鞭节各亚节端部以及亚节与分支的交界处,数量较少(岳璐等, 2014)。纽扣形感器在广翅目和膜翅目昆虫触角上均有分布(表1)。
1.26 剑梢感器(Sensilla scolopadia)
附于体壁下面的感受器,由感觉细胞、冠细胞和围细胞组成,是典型的弦音感器。在虫体上一定的部位常有由若干剑梢感器集在一起的感觉器官,包括江氏器和鼓膜器等(那杰等, 2007)。江氏器最早由Johnston于1855年在雄性埃及伊蚊Aedesaegypti成虫的触角梗节内发现,是一种机械感受器。鼓膜器普遍存在于发音昆虫中,包括由表皮形成的鼓膜和内气囊等(马瑞燕和杜家纬, 2000)。鳞翅目、膜翅目和鞘翅目昆虫触角上着生有剑梢感器(表1)。
1.27 竹丫形感器(Bamboo branch-like setae)
形似竹子枝丫的分支,大部分在主干距顶端1/2处开始分支,形成竹丫状。一般着生于鳞状表皮的凹陷处,自基部向端部逐渐变细,且表面有纵脊(姚丹等, 2021)。竹丫形感器的研究较少,目前仅在膜翅目昆虫触角上发现有这类感器,且只在一个物种中发现(表1)。
1.28 刚毛(Setae)
有叶状刚毛和松针状刚毛两大类。叶状刚毛形似韭菜叶片,扁平,表面光滑无纵脊,从基部到端部逐渐变细,其中在顶部略有弯曲,与基部呈60°左右的夹角,且弯曲方向一致,着生的表皮处有凹陷。松针状刚毛分为有明显螺纹和无明显螺纹2种,有明显螺纹的刚毛较无明显螺纹刚毛长而细,螺纹自根部似扭转而上,越往上越不明显;无明显螺纹刚毛也似栓锥形(赵慧婷等, 2019)。此类感器仅在膜翅目昆虫触角上有发现(表1)。
1.29 端指形感器(Sensella finger-like)
端指形感器形态稍侧扁,基部与端部大小基本一致,壁上具明显的纵脊,形似植物的“维管束”,并于末端分叉3~5支,分支长短不尽相同,形似手指,分别指向触角的端部(罗术东等, 2011)。目前发现端指形感器主要分布在膜翅目和鞘翅目昆虫的触角中(表1)。
1.30 棒形感器(Sensilla rod-like)
棒形感器与端指形感器相似,但棒形感器为圆形,端部和基部几乎等粗,表面具有明显的纵条纹,稍弯曲,末端近截型,但边缘不整齐,与触角表面的角度为15~20°(张新民等, 2021)。在目前的研究中,鳞翅目、膜翅目、鞘翅目和双翅目昆虫中都存在棒形感器(表1)。
1.31 表皮孔(Cuticular pore)
在触角表面分布的许多不规则小孔(张健等, 2007)。仅在膜翅目和鞘翅目中发现有表皮孔的存在(表1)。
1.32 棘刺(Spine)
相邻棘刺的基部相连,基部呈鱼鳍状,顶端尖细,尖端分叉或不分叉,并沿触角轴线向触角尖端一侧弯曲(马涛等, 2019)。目前仅在鳞翅目昆虫中有发现(表1)。
1.33 锯齿形感器(Sensilla zigzag)
形似锯齿,表面有纵向条纹,在侧面有多个锯齿,着生于基窝内(王全坡等, 2013)。在鞘翅目的昆虫触角上有分布(表1)。
1.34 栓锥形乳突状感器(Sensilla basiconic capitate peg)
感器位于凹槽内,高度略高于凹槽,状如乳突,基部较为膨大,表面具有多条纵向沟槽,分为栓锥形乳突状感器I型和栓锥形乳突状感器II型,前者顶端较钝,后者顶部尖细(刘志祥等, 2021)。该类感器在膜翅目、鳞翅目、鞘翅目昆虫触角上均有分布(表1)。
1.35 盘形感器(Sensilla disk)
中央凹陷,呈椭圆形,外周有沟壑状突起,沟壑上着生有毛形感器,分为盘形感器I型和盘形感器II型。盘形感器I型周边一般有毛形感器呈旋形分布,盘形感器II型外周有3层沟壑状突起,其上着生有3种毛形感器(王颖娟和李子忠, 2012)。只在脉翅目中发现盘形感器(表1)。
1.36 角锥形感器(Sensilla obscurum)
位于凹槽内,基本平伏于凹槽表面,形似锥体,表面有纵向沟槽,顶部柔软易弯曲(刘志祥等, 2021)。仅在膜翅目中有角锥形感器(表1)。
1.37 腔乳头感器(Sensilla cavitata-peg)
呈乳头状,端部钝圆,深陷在宽阔的凹窝内,突起物高于腔深或接近于腔的上表面(刘群, 2014)。腔乳头感器在鳞翅目和脉翅目昆虫触角中分布(表1)。
1.38 槽纹感器(Sensilla fluted)
形似火炬,下半部如锥形,表面光滑,基部往上逐渐变细,上半部有纵向扭曲向顶端延伸的条纹,形成凹槽状,沟纹明显(王全坡等, 2013)。在鞘翅目昆虫触角上有分布(表1),但只在极少数的类群中被发现到,如鞘翅目昆虫。
1.39 火柴形感器(Sensilla match-shaped)
形似火柴,由棒状体和顶部的小圆球构成,棒状体表面光滑,小圆球表面凹凸不平,基部无凹状基窝(李文禄等, 2011)。截至目前,只在同翅目昆虫中发现火柴形感器(表1),可能是因为这类感器较为罕见,只在特定的物种中出现。
1.40 鳃形感器(Sensilla branchia)
感器表面有条纹凹陷,其上有4条横纹平行排列,形似软骨鱼的鱼鳃(王颖娟和李子忠, 2012)。鳃形感器分布在脉翅目的昆虫触角上,并且目前仅在一个物种中有发现(表1)。
1.41 爪状感器(Sensilla claw-like)
生长于表皮内陷的窝内,其端部有3个小指,到中部愈合(梁雪梅, 2014)。该类感器在少数半翅目昆虫触角上有分布(表1)。
1.42 U形感器(Sensilla U-shaped)
也被称为叉状感觉锥,形似U字母,着生于浅基窝内,基部膨大粗壮,两臂在顶部皱缩呈针状(黄丽莉等, 2016)。仅在缨翅目昆虫触角上发现有U形感器的分布(表1)。
1.43 带状感器(Sensilla zonated)
狭长呈带状弓形,部分略扭曲,与触角表面呈锐角(于玮台和陈文龙, 2013),在半翅目昆虫中发现带状感器(表1)。
1.44 长刺具泡状感器(Sensilla long trichodea with a bubble)
基部与长刺形感器相似,其上部位具有泡状的隆起(张新慰等, 2015)。在等翅目中首次发现,在其他昆虫触角上还未发现这类感器的分布(表1)。
1.45 水滴形感器(Sensilla teardrop)
呈水滴状,基部膨大如球,近基部到顶端急剧缩小为刺状(楚君鹏等, 2019)。仅在等翅目昆虫触角中发现水滴形感器(表1)。
1.46 镰刀形感器(Sensilla sickle-shapped)
基部粗,顶部骤尖,形状弯曲似镰刀(高素红等, 2010)。在鳞翅目、膜翅目和鞘翅目3个目的昆虫中都发现了镰刀形感器的分布(表1)。
1.47 具弯钩形感器(Sensilla bent-tipped)
着生于半球状突起上,顶端呈弯钩状,并向触角顶部倾斜生长(孙猛等, 2017)。在半翅目中具有这类感器(表1)。
1.48 具沟端弯感器(Sensilla fluted bent-tipped)
着生于凹窝内,细短有纵沟,端钝弯,垂直着生(高其康和胡萃, 1993)。只在膜翅目昆虫触角中发现具沟端弯感器的存在(表1)。
1.49 脚形感器(Sensilla feet-like)
表面光滑,顶端扁平,形似人脚(谭琼, 2012)。在鞘翅目和鳞翅目的少数物种中分布(表1)。
1.50 念珠形感器(Sensilla moniliform)
呈念珠状(赵冬香等, 2006)。只在半翅目昆虫触角上发现到了念珠形感器(表1)。
1.51 盾形感器(Sensilla scutellatum)
形似钢笔笔头,中间宽,顶端尖,几乎与触角表面垂直,呈盾片状(王长青等, 2009)。鞘翅目和直翅目的部分昆虫中有盾形感器的分布(表1)。
1.52 星形感器(Sensilla stellate)
顶端呈星形,整体侧观呈宝塔糖状,四周具深沟槽,整体向触角顶部倾斜(宋珊珊等, 2017)。在鳞翅目中大部分昆虫都具有星形感器,是鳞翅目昆虫独有的一类感器(表1)。
1.53 V形感器(Sensilla V-like)
着生于凹窝内,顶部呈V字形分叉,表面不光滑(罗丹等, 2018)。仅分布在膜翅目昆虫触角中(表1)。
1.54 三角形感器(Sensilla trinagle)
基部宽扁,顶端尖锐,从基部到顶部由宽变窄,呈三角形(郭梅娜和陈祥盛, 2015)。在同翅目和半翅目昆虫触角上发现有三角形感器的存在(表1)。
1.55 桑椹形感器(Sensilla mulberricum)
由触角表皮突起形成的一个个紧密相接的瘤状结构,整体形态类似桑椹(任国栋等, 2013)。仅在鞘翅目个别种类中发现该感器的分布(表1)。
1.56 齿形感器(Sensilla dentiform)
从中部分为两叉,顶端尖细(李秀敏等, 2010)。目前,这类感器只在部分的鞘翅目昆虫触角中分布(表1)。
1.57 半球形感器(Sensilla hemisphere)
呈半球状,表面光滑,基部凹陷,顶端与表皮持平或略高(楚君鹏等, 2019)。在等翅目中发现有半球形感器的存在(表1),但可能是由于等翅目的研究较少,截至目前也只是在少数类群中分布。
1.58 球形感器(Sensilla knob)
顶端有一细长且不直的柄(田哲豪, 2021)。在半翅目昆虫中发现到了这类感器(表1)。
1.59 端毛(Terminal hair)
触角微小圆锥状突起上的细毛(沈杰等, 2005)。当前仅在鳞翅目昆虫的触角上有分布(表1)。
1.60 火山形感器(Sensilla volcanica)
端部钝圆,状如火山,位于一个卵圆形凹窝内(薛爽等, 2021)。只在膜翅目中发现有火山形感器的存在,非常少见(表1)。
1.61 环丝(Circumfila)
一类形似花冠的环状感器,通过凹槽内的一系列短茎连接到触角表面(张凡, 2020)。目前只在双翅目枸杞红瘿蚊Jaapiellasp.的触角中发现到了环丝(表1)。
2 昆虫触角感器种内与种间特征差异
昆虫触角感器类型多样化可能是环境压力导致昆虫适应生境的结果。不同种类昆虫在各种环境压力的不断作用和选择下,致使各类昆虫触角感器在种类和数量上发生了较大的变化。因此,触角感器类型、数量和分布等存在着明显的种内和种间差异。
2.1 昆虫触角感器的种内特征差异
首先种内差异体现在雌雄两性之间。已有观察和研究发现一些昆虫种类的雌雄性成虫之间在体色上存在较大差异,触角感器往往也会呈现出性二型特征(程红, 2006),即在类型、数量、分布区域上都有所不同。已有研究表明,双翅目的红腹毛蚊Bibiorufiventris雌雄成虫触角感器在毛形感器和锥形感器的亚型上有所差异,雌性是毛型感器Ⅱ型和锥形感器3种亚型,而雄性是毛型感器Ⅰ型和锥形感器2种亚型(袁轲等, 2020)。食蚜瘿蚊Aphidoletesaphidimyza的雄虫触角上存在柱形感器,但是雌虫的触角上未发现该类感器(蔡普默等, 2021)。针对鞘翅目昆虫触角感器的研究表明,斑鞘豆叶甲雌雄成虫的触角感器则在数量、大小上存在不同的表现(徐伟等, 2020)。蓟马科的牛角花齿蓟马Odontothripsloti、端大蓟马Megalurothripsdistalis以及卡式绢蓟马Sericothripskaszabi的触角感器数量在雌雄上有所差异,牛角花齿蓟马中雌性锥形感器Ⅲ型的数量多于雄性,3种昆虫中位于基部柄节的雌性刺形感器Ⅰ型多于雄性(Liuetal., 2021)。鳞翅目的蛾类雄性昆虫的毛形感器Ⅱ型在数量上多于雌性(那杰等, 2007),还有研究证明美国白蛾Hyphantriacunea的毛形感器在雌雄性之间也分为长短两种(狄贵秋等, 2020),长毛形感器多分布于雄虫触角上(余海忠, 2007)。尽管针对昆虫触角及其感器的性二型特征还没有明确其原因(杨慧, 2008),但有研究推测,雌雄之间的触角感器类型及其亚型差异可能是由于识别性信息素、寄主释放的化合物以及生存环境的不同外界机械刺激引起的(Huetal., 2009)。雌性昆虫除了寻找食物等,最大化后代繁殖也极为重要,将会有一类触角感器在寻找合适的寄主中发挥重要作用;而雄性昆虫在发现食物基础上,还需要依赖性信息素定位配偶,因此将有不同触角感器执行这种功能。然而,产生触角感器性二型的内在机制是长期进化过程还是生态因子的作用仍需要更多的深入探究。
除了雌雄差异,昆虫触角感器的种类和数量等特征在昆虫的不同发育阶段之间也存在差异(Brézotetal., 1996; 李竹和陈力, 2010)。有研究表明,昆虫特定部位的触角感器类型在发育过程中消失,如灰飞虱Laodelphaxstriatellus的1龄若虫鞭节基部的毛形感器在第一次蜕皮后退化消失(Fuetal., 2013)。还有的昆虫在不同发育期时触角感器的种类数量也存在变化,例如,黑翅土白蚁Odontotermesfomosanus的1龄幼蚁具有7种感器,2龄幼蚁具有8种感器,而3龄幼蚁具有9种类型触角感器(张新慰等, 2015)。此外,还有研究发现东亚飞蝗Locustamigratoriamanilensis成虫、5龄若虫与4龄若虫的触角感器类型存在显著性差异(高倩等, 2007)。因此,可以看出,昆虫触角感器随着昆虫发育过程显著改变,这可能是由于不同发育阶段与外界环境相适应的结果。同时,这些研究结果也可以表明触角感器的特点也可能成为判断昆虫发育阶段的重要依据之一。
2.2 昆虫触角感器的种间特征差异
2.3 触角感器在属级及以上分类阶元的特征差异
高级分类单元中,昆虫触角感器的差异性是极为明显的,其中膜翅目的昆虫触角具有较为特殊的坛形感器,鞘翅目作为昆虫纲中最大的目,其较为特殊的触角感器主要是芽孢形感器,鳞翅目相比于其他感器较特殊的则是剑梢感器(马瑞燕和杜家纬, 2000)。据相关报道,几乎所有的寄生蜂都分布着大量的板形感器(Castrejnetal., 2003)。这意味着不同类群的昆虫触角特异性感器类型可作为类群鉴别的重要形态学依据。少数研究学者针对单一目级分类单位的昆虫触角感器进行总结,而对多个目级却鲜有比较。
目前,大多数研究主要集中在某一种昆虫的具体触角感器,很少研究触角感器在不同种、不同属和不同科等之间的差异性。在将来的研究中应更多关注种内与种间的昆虫触角感器类型与数量的比较,不仅能够解析昆虫与其所处环境的密切联系,也为不同昆虫在觅食、防御等行为存在差异提供形态学基础。
3 昆虫触角感器功能
触角是昆虫的主要感觉器官,其表面着生各种类型的感器感受机械刺激和化学物质信息,实现个体间的信息交流,完成觅食、求偶、繁殖和躲避天敌等基本生物学行为(蔡普默等, 2021)。不同类型的感器执行不同功能。因此,研究和认识昆虫触角感器功能对于理解昆虫与外部环境的协同适应关系及行为可塑性具有重要意义(Sukontasonetal., 2004)。目前,已有一些研究针对昆虫触角感器的功能进行了验证,发现一些感器类型对寄主挥发性气味以及生境中的气味极为敏感,起到了关键的嗅觉功能。
3.1 毛形感器
毛形感器是昆虫触角感器中较为常见的类型,其功能也较为复杂。有研究表明,毛形感器是最基础的嗅觉感器,兼具有感受性信息素和寄主挥发物的作用(马瑞燕和杜家纬, 2000)。蛾类触角上的毛形感器含有非常丰富的信息素结合蛋白,因此,对环境中存在的以及异性释放的化学物质具有较强的结合能力。在家蚕和棉铃虫Helicoverpaarmigera研究中发现,毛形感器在探测性信息素中发挥着关键性作用(Shiotaetal., 2018; Liuetal., 2020)。同时,有学者还推测毛形感器可以帮助昆虫感受寄主挥发物从而寻找合适的产卵场所(徐伟等, 2020),Silva等人(2018)在鳞翅目蛱蝶科昆虫Heliconiuserato的触角感器功能研究中证实了毛形感器的这种重要功能(Silvaetal., 2018)。根据其毛状的外形,推测其除了具有接收化学信号的功能外还能够感受气流,其在雌雄蜂鞭节处有着相反的分布规律,两性个体运动状态差别大,这间接印证了毛形感器可能影响物体运动状态的识别(Dahakeetal., 2018)。
3.2 刺形感器
刺形感器属于感知机械刺激类的感器,但也有研究表明刺形感器可以感受化学信息。在红棕象甲Rhynchophorusferrugineus的研究中发现,雄虫触角柄节有刺形感器,但雌虫的相同部位却未见该类型感器,认为可能与雄虫在相互打斗竞争交配权时感知对方攻击的机械刺激有关(段云博等, 2020)。刺形感器除了可以感知昆虫的机械刺激外,还可以对外界环境的机械刺激做出选择,这种功能在沙棘木蠹蛾Holcocerushippophaecolus的触角感器研究中得到证实(Wangetal., 2015)。此外,还有学者对象甲科的松梢象Pissodesnitidus进行研究推测,刺形感器是执行嗅觉功能的化学感受器,主要是与雌虫寻找产卵场所感受寄主植物挥发的化学物质有关(Yanetal., 2011)。
3.3 锥形感器
锥形感器通常被认为是一种嗅觉感器(Altner and Prillinger, 1980),主要用于气味识别(于庭洪等, 2020),所以在搜寻寄主、趋避危险和寻找食物源等行为中起到关键的作用(王涛, 2017),锥形感器的气味识别功能在红脂大小蠹Dendroctonusvalens的触角感器功能研究中得到证实。而且,有学者在鞘翅目天牛科昆虫Lepturaarcuata的研究中发现,雄性成虫锥形感器的数量显著高于雌性成虫,据此推断雄性触角上的锥形感器具有识别雌性性信息素的功能(Zhangetal., 2011)。
3.4 腔锥形感器
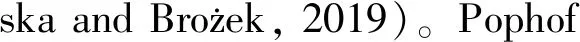
3.5 腔形感器
腔形感器同样是典型的化学信息接受感器。在稻纵卷叶螟Cnaphalocrocismedinalis的触角感器的研究中证明腔形感器能够识别植物释放的气味分子(Sunetal., 2011)。还有研究者发现茶棍蓟马Dendrothripsminowai只有雌性成虫触角和若虫触角上有腔形感器,推测该感器可能参与对异性个体定位交配等活动(吕召云等, 2015)。此外,腔形感器是卡氏步甲Carabuscasaleianus中数量最多的一种感器,推测可感受外界机械刺激,主要起到感受外界湿度的功能(齐钦等, 2015)。
3.6 栓锥形感器
栓锥形感器在夜蛾科中普遍存在(Guetal., 2013)。对鳞翅目昆虫的幼虫口器下颚上的栓锥形感器开展研究,发现这种触角感器对糖和阻碍剂反应敏感(Tangetal., 2015)。同时在双委夜蛾Athetisdissimilis的研究中推测触角上的栓锥形感器具有接受化学信息,感知气味的功能(宋月芹等, 2018)。还有学者推测栓锥形感器在螟蛾科的昆虫中可接受外界刺激,感知温度或湿度(于庭洪等, 2020)。
3.7 板形感器
板形感器能够接受化学刺激。在寄生蜂的产卵行为研究中,发现寄生蜂通过板形感器识别寄主生境中的气味(那杰等, 2007)。但是,Ochieng等人用触角单感器记录技术研究红足侧沟茧蜂Microplitiscroceipes触角上的板形感器时,发现板形感器对植物挥发物具有反应,且反应的强弱随着挥发物浓度增加而增强(Ochiengetal., 2000)。此外,根据蚜虫板形感器的超微结构,和大豆蚜Aphisglycines触角对报警信息素的响应实验,证实了板形感器可感受报警信息素(赵立静和班丽萍, 2011)。
3.8 钟形感器
钟形感器是最典型的机械感器(徐伟等, 2020)。鞘翅目步甲科的黑广肩步甲Calosomamaximoviczi可依靠触角上的钟形感器对栖息地中的温湿度进行探测,避免选择过热或过冷的生境(于庭洪等, 2020)。而且,有学者认为钟形感器还能够感受角质层的机械形变,对表皮的变化迅速做出反应(余海忠, 2007),还可能具有感受触角鞭节的运动产生压力的功能(Dunn, 1978)。
3.9 波氏鬃毛
有学者认为波氏鬃毛是一种感受重力的机械感器,当遇到机械刺激时,能够缓冲重力的作用力,从而控制触角下降的速度(董文霞和张钟宁, 2006)。同时,在染翅蝎蛉Sinopanorpatincta和中华蚊蝎蛉Bittacussinensis的研究中发现,波氏鬃毛具有介导昆虫飞行中的定位功能,当将它们从基部消融掉后,触角会与翅碰撞,从而阻碍翅的运动和触角的作用(Shields and Hildebrand, 2001)。
3.10 芽孢形感器
芽孢形感器集中分布于节间部位,当触角受到外界的机械刺激时,芽孢形感器能够起到缓冲重力的作用,因此芽孢形感器可能是一种感受重力的机械感器(Bartletetal., 1999)。对褐纹甘蔗象Rhabdosceluslineaticollis的研究表明,触角受到外界机械刺激时会主动收缩,该过程可能与触角折叠收缩时芽孢形感器感知梗节和喙的机械刺激从而调整收缩程度有关(段云博等, 2020)。
3.11 叉形感器
叉形感器利用透射电镜观察其内部结构,推测可能具有感受机械刺激和化学刺激的功能(Renetal., 2012)。根据其形态推测可能还具有控制触角方向的功能,又因其主要着生于背面,推测可能与昆虫的避阳趋阴性有关(Wangetal., 2012)。Frank等人对茶丽纹象甲Myllocerinusaurolineatus的研究发现,叉形感器在茶丽纹象甲雌、雄虫间的数量差异显著,故推测叉形感器还可能感受化学刺激,具有嗅觉功能,与感受种内信息素有关(Franketal., 2009)。
3.12 剑梢感器
剑梢感器可以感知外界的机械刺激,是典型的弦音感器,虫体上的一定部位常有由若干剑梢感受器集在一起的感觉器官,如江氏器,它能感受声波,辨别音调的刺激,是昆虫的听觉器官(那杰等, 2007)。
总之,关于昆虫感器功能大部分仍然停留在推测阶段,仅有少量研究提供实验性证据。随着更多触角感器的发现,触角感器功能研究不仅仅局限于以往的类型,对于很多新发现的感器认知并不清晰,特别是寄主、配偶等方面的识别功能缺乏深入的理解。随着研究的深入,伴随着相关技术手段的快速发展,昆虫触角感器研究逐步进入到超微结构,甚至是单一感受器,检验这些触角感器功能将成为当前的研究热点。
4 未来研究方向
近年来,关于昆虫触角化学感受外界环境(包括温度、湿度和CO2以及寄主植物)的内在机制研究备受关注,是当前国内外相关领域的研究热点之一。昆虫触角着生的不同类型的感受器是昆虫感受环境中化学信息物质的基本结构单元,将化学信号转换为电信号刺激昆虫神经元的树突膜,并传导到神经中心,从而产生一系列行为反应。随着科技的快速发展,精密仪器的出现推动了昆虫触角感器的研究进展,已经从扫描电镜技术逐渐过渡到透射电镜技术开展昆虫触角的形态学和超微结构的研究,并开始探究其功能,但仍然可以从以下几方面拓展昆虫触角感器的研究角度。
第一,绝大部分昆虫的触角感器研究仍然停留在扫描电镜观察触角感器类型以及在触角上的分布状况,这种研究方法限制了理解不同类型的触角感器如何连接神经中枢。因此,需要开展更多透射电镜观察研究,以便更深入的探讨昆虫触角感器在外界理化信号通过神经元进入脑的过程中发挥的作用。同时,大多数研究只是针对某一种昆虫触角感器类型及分布进行研究,缺少种内与种间的对比性研究工作。通过本文的系统性总结发现部分昆虫种类存在少见或是独有的触角感器,因此,加深这方面研究能为昆虫的分类提供一定的思路。
第二,截至目前,更多的研究主要停留在对昆虫触角感器类型以及其分布研究方面,而更深层次的感受器功能的认识还极为缺乏,大部分触角感器的功能仅处于推测阶段,缺少实验性证据,因此,结合扫描电子显微镜、透射电子显微镜等深层技术的运用,可深入挖掘触角感器在寄主识别、昆虫交配等行为中的重要作用,有效推动昆虫触角感器的结构和功能与昆虫适应性行为关系研究。
第三,触角感器研究主要集中在鞘翅目、鳞翅目、膜翅目和半翅目等昆虫类群,且大部分是针对害虫及其天敌昆虫开展的,其目的是为科学有效防控害虫提供研究基础。随着传粉昆虫重要性逐渐被认识到,未来可深入探讨重要传粉昆虫的触角感受器研究,不仅有利于丰富昆虫触角感器类型多样化,还能够在理解访花昆虫识别开花植物机制方面提供重要的形态学依据,同时更能为开发吸引传粉昆虫的引诱剂提供理论基础。
猜你喜欢
热带农业科学(2023年8期)2023-09-25
中国棉花(2022年1期)2022-04-06
河南农业科学(2019年5期)2019-05-28
电子测试(2018年18期)2018-11-14
陕西理工大学学报(自然科学版)(2018年5期)2018-11-06
安徽农业科学(2018年20期)2018-05-14
湖北农业科学(2017年24期)2018-01-27
现代农业科技(2017年22期)2018-01-08
安徽农业科学(2017年18期)2017-07-10
河北林业科技(2016年5期)2016-11-08
