
昆虫趋光性研究的回顾
2023-11-09 12:15武予清蒋月丽周国涛张改平巩中军
环境昆虫学报 2023年5期
武予清,蒋月丽*,周国涛,张改平,苗 进,巩中军,段 云,李 彤
(1.河南省农业科学院植物保护研究所,河南省农作物病虫害防治重点实验室,农业部华北南部有害生物治理重点实验室,郑州 450002;2. 河南云飞科技发展有限公司,郑州 450000;3 河南省禹州市农业农村局,河南禹州 452570)
我国的成语“飞蛾扑火”,意指无法抵抗灯火的魅力而自取灭亡的昆虫对光的趋性行为,出自隋·姚察等《梁书·到溉传》中的“如飞蛾之赴火,岂焚身之可吝。”。在距今1400年前,昆虫的趋光性已经被人们作为著名的生物现象,在艺术表现形式中,有唐朝诗人张祜(约785年—849年)赠内人“禁门宫树月痕过,媚眼唯看宿鹭窠。斜拔玉钗灯影畔,剔开红焰救飞蛾。”的著名诗句;瓢虫类的甲虫,因其面向太阳的定位行为飞行,人们认为它是太阳神的使者,从而被命名为“天道虫”。昆虫的趋光性现象在日常生活中已有广泛应用,如在农业、林业、制造业、零售业、学术研究、医疗等各种场所,已经利用了光源作为刺激源(Cantelro,1974)。另外,Gilbert(1984)在有关昆虫电击灯诱捕器的报告中,据推测使用了20万个工业设计的光诱捕器。我国自上世纪90年代棉铃虫暴发以来,杀虫灯广泛用于农林害虫的防控。
由于害虫绿色防控和生物多样性保护的需求,从“对环境友好”的观点出发,通过光来预测、预防害虫,以及对益虫、中性昆虫和稀有昆虫的引诱、回避等习性的利用,一直是一种重要的害虫治理技术。
随着对害虫防控从绿色防控走向生态防控的更高需求,进一步开展昆虫趋光性的应用,就需要对昆虫趋光性机理及生态适应性有更深刻的了解,以开发更有效的利用光的广谱性或专化性诱虫技术,因而有必要解释昆虫聚集在光源周围的现象,并对昆虫趋光行为多样性和趋光性机制进行不断揭示和理解。
1 动物趋光性的定义
动物的趋光性(Photaxis)在狭义上被定义为动物空间内光源本身的定位和运动的方式,根据运动的方向性被分类为两种,朝向光源时,称为正趋光性(Positive photaxis)。远离光源时,称为负趋光性(Negative phototaxis)(Campan, 1997)。
但是,要严格区分动物对趋光的现象是否是对光源定位的结果是不容易的,这是因为动物的行动受到光多种方式的影响(弘中 and 針山,2009;Shimoda and Honda,2013),动物可以通过各种行动机制在光源聚集。表面上看,这种情况似乎是动物对光源明亮部分显示出正的趋光性。但是,即使不依赖于这种狭义的趋光性(图1-a),动物也可以通过各种行为机制聚集到光源,动物到达光源附近(图1-b,根据动力学的表观定位)。当动物通过光刺激激发变向运动(Klinokinesis)或变速运动(Orthokinesis)时,通过对光源反应来改变方向转换的频率和移动速度,那么对光源定位能被观察到。另外,光源对周围产生的其他光刺激定位,有时会被观察为定位在光源上(图1-c,对周围光环境的定位:参照后述的马赫带理论)。
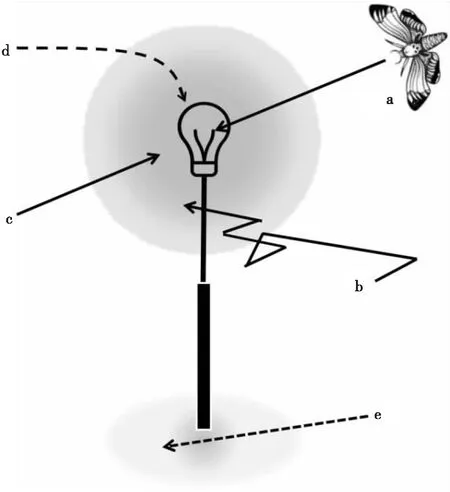
图1 昆虫聚集在光源及附近的多种形式
此外,由于光源附近的活动受到阻碍,从外观上看,动物也有可能聚集在光源附近(图1-d,因活动阻碍的外观上定位)。例如飞行昆虫,已知背光反应(Dorsal light reaction)是其姿势的定位是从背侧接受光,具有这种习性的飞行性昆虫被从下方照射到强光阻碍了姿态定位,所以能观察到落在光源附近的个体。如果是暴露在某个阈值以上的亮度下,会阻碍动物的移动行为,即使是完全没有趋光性的物种,也会在光的周围聚集。以夜蛾类为代表的一部分夜行性昆虫,已知存在着有光行动抑制现象(野村等,1965),这种行动反应会因阻碍行动而引起到光源处集中。而且,由于受到光源附近存在饵料味道的引导等与光刺激完全无关的刺激,昆虫也有可能聚集在光源附近(图1-e,间接定位)。
广义的趋光性多用于通过光依赖性的定位机制结果接近光源的行为,或结果远离光源的行为。在这样的范畴中,根据上述的动力学的表观定位,通过趋暗性(Scototaxis)接近光源周围的行动(马赫带理论)也包含在趋光性中,但是,在应用的观点中,动物是否“直线性、主动地”被吸引到光源本身是重要的要素,由于运动性(Kinesis)的表观定位等广义的趋光性中包含的内容,以及对行动阻碍和间接刺激的反应等与趋光性不同的行动反应,有必要与狭义的趋光性明确区别研究,在处理趋光现象的情况下,详细观察目标动物的光源定位和运动过程,获得其移动轨迹是不可缺少的,并且动物将光源作为刺激源,在描绘曲线的同时或者直线地向光源移动的情况下,狭义趋光性的可能性很高(参照后述的指南针理论,或开放空间理论)。
这里,“趋光性”主要处理昆虫光源的正趋光性,并用作“广义趋光性”,包括运动性(Kinesis)的表观定位和周围光环境的定位,把由趋光性引起的昆虫在光源上的集合记述为“趋光现象”。
对于颜色光(光波长)的趋性,一般是白天活动或微弱光环境下(杨小凡等,2021)的昆虫对颜色板(物体反射光,也称为非自发光体)趋性,如蚜虫、粉虱对黄色粘板的趋性,就不在本文讨论。
2 昆虫趋光现象的特征
在理解昆虫的趋光性时,首先要了解迄今为止观察到的趋光性现象。表现出趋光性的昆虫物种所具有的特征,在研究趋光性的适应性意义和行动机制方面能够给予人们重要的启示:引起趋光现象的环境条件、被光源吸引的个体特征等信息,从应用的观点来看这些也是有用的。
2.1 被光源吸引的物种
在为了调查记录生活在某个地区的昆虫种类目录清单中,光诱捕器(Light trap)作为重要的调查方法被广泛使用。在河南省昆虫种类大规模调查中,8 637种昆虫种类中有30%的种类来自光诱捕器(申效诚,2009)。作为对物种数量进行计数的调查,鳞翅目昆虫的光诱捕器的调查在多个类型的地区进行。根据Frank(2006)的总结,即使是限定于大型鳞翅目昆虫,也有数百到千种以上的种类在所有调查中被光吸引诱集,半翅目记录也有118种(Southwoodetal.,2003)。丁建云和张建华(2016)收集到的北京灯下蛾类有27科494种。高月波等(2021)收集到的吉林省灯下蛾类也有17科235种。这些结果表明,夜行性昆虫的趋光性是极为普遍的反应。
夜间聚集在人工光源上的昆虫不限于夜行性的物种。例如,有多个记录是白天活动的昆虫被人工光源所吸引(Heitzman,1965;Chowdhury and Soren,2011),一般认为是夜间被扰乱休息场所而被赶出去的昼行性昆虫在寻找新栖息地的过程中聚集在人工光源上的结果。
光源不仅吸引了飞翔的昆虫种类,还吸引了地表徘徊行走的种类,例如,不飞翔的西印度甘薯象甲Euscepespostfasciatus(鞘翅目Coleoptera象鼻虫科Paramecidae)的成虫被放置在野外的光诱捕器捕获(仲本&澤岻, 2001)。巴西猎蝽Triatomainfestans(半翅目猎蝽科Reduviidae)的幼虫通过步行接近野外的光诱捕器(Vazquez-Prookopecetal., 2006)。据报道,西印度甘薯象甲(Katsukietal., 2012)、赤拟谷盗Triboliumcastaneum(Duehletal.,2011)和几种步甲(Andersen, 1989)等游荡性物种通过步行接近光源。
2.2 影响诱集的环境条件
如果在同一个地方持续采集的光诱捕器,采集到昆虫的数量会随着时间发生很大的变化,这是因为天气等环境条件会影响昆虫的活动性和对光源定位这两个行动要素,光、湿度、风、大气压、雷和闪电这六种气象条件,都会影响昆虫光诱捕器的诱导,由于低温的影响,诱集数降低,这是由于这些因素抑制了昆虫的飞翔行为等活动性(Williams,1940;Hardwick,1972;Persson,1976;McGeachie,1989)和强风(Persson,1976;Mizutani,1984;McGeachie,1989)。圆臀大黾蝽Aquariuspaludum(半翅目Hemiptera黾蝽科Gerridae)试验证实,温度环境对定位行动本身有影响,成虫阶段的高温引起正趋光性,低温引起负趋光性(Harada,1991;原田,1996)。
在考虑昆虫趋光性机制的基础上,直接影响昆虫对光源的定位行为的环境条件也很重要。例如,许多昆虫被用于研究月相、月光量与光诱捕器的诱捕数量之间的关系。很多物种在接近新月期时,诱集数会增加,出现数倍的差异(Williams,1940;Nowinszkyetal., 1979)。同样的倾向,也可以认为是受其他光源引诱的昆虫种类(Eisenbeis,2006)。满月期诱集数下降趋势的原因,是可能在满月前后昆虫的活动性下降(Bowden,1973)和对光源的定位行动可能被满月阻碍。由于在临近满月也采集到很多个体,所以月亮与光源的吸引数之间的关系,并不是因为活动性依月相而增强或抑制,而是因为月亮对光源的定位行为本身产生了某种影响,由于作为背景的黑暗变亮,考虑到作为昆虫被吸引范围的光源的诱集半径有变小的可能性(Bowden and Church,1973;Eisenbeis,2006)。背景的光强度的重要性是,在光量较低的阴天的夜晚,诱集数增加,林冠遮蔽了来自天空的光,在林内吸引了很多个体,这种观察结果也支持背景光强度是对诱集有影响的(Bowden,1982)。另外,月亮产生的夜空的偏振光可能影响到光源的诱导,Nowinszkyetal.(2012)观察到,毛翅目Trichoptera的石蛾类(Caddisflies)在夜空偏振最大的上弦和下弦月时,光诱捕器的诱集数最大,偏振光不是影响着这些昆虫对光源的定位行动,而是由于利用月球偏振光作为定位的昆虫,在上弦和下弦的月龄期间,增加了飞翔个体数。
另外,有研究表明日照长度对趋光性有影响。黾蝽在14.5 h的长日照条件下饲养的成虫显示出强烈的正趋光性。另一方面,试验证实,在9.5 h的短日照条件下饲养的成虫显示出负趋光性(Harada,1991)。
2.3 诱集个体的特征
多种昆虫,在特定活动周期内的个体很容易被吸引到光源。一些试验证实了活动周期对趋光性的影响,如黄曲条跳甲Phyllotretastriolata趋光性的作用光谱测定,在上午8∶00-12∶00,下午13∶00-18∶00,晚上18∶00-24∶00这三个时间段进行,结果表明,与上午相比,下午和夜间这两个时段,昆虫的趋光性灵敏度低,特别是,明确了对于430 nm以下的波长反应性低(Yangetal., 2003)。
在受光吸引的昆虫中,有时会出现幼虫虫龄和成虫虫龄等特征,有报告显示老熟幼虫具有负趋光性(Olson and Rings,1969;Archer and Musick,1976;Shields and Wyman,1984;Griswold and Trumble,1985)。例如,彩地老虎(黄毛夜蛾)Triphaenapronuba,在评价幼虫的趋光性的试验中,从半边涂满黑漆的盘子上方照射光,计算了盘子的照明部和遮光部中存在的个体数。结果显示,1龄幼虫具有显著的正趋光性,7龄幼虫具有显著的负趋光性,在它们之间的年龄段,观察到中间反应(Madge,1964)。除此之外,赤拟谷盗Triboliumcastaneum和杂拟谷盗Triboliumconfusum,成虫羽化后,2~3日龄有很强的负趋光性倾向,而4~5日龄时正趋光性的倾向很强(Arbogast and Flaherty, 1973)。
在蛾类中,雄蛾被光诱捕器吸引的倾向很强,这是因为它们大多数在自由活动的空间中,雄性在很大范围内飞翔,因而容易被光诱捕器捕获(Robinson and Robinson,1950;Frank,2006)。Robinson and Robinson(1950)在设置的自由空间的光诱捕器中只采集了枯叶蛾科Lasiocampidae的Trichiuracrataegi的雄蛾,在树木附近设置的光诱捕器中,只采集到了雌蛾,也有试验观察到性别差异的报告例子。在调查黑蝇科的红头丽蝇Calliphoraerythrocephala对于50 W卤素灯的趋光性的研究中,性别也存在很大的差异。雌蝇在羽化后第2天显示出强烈的正趋光性后,在第3天之后表现出了急剧增强的负趋光性。另一方面,雄虫在第2天同样表现出正趋光性后,观察到了大约3 d间隔的周期有交替出现正和负趋光性的倾向(Meyer,1978)。卡罗拉多马铃薯甲虫Leptinotarsadecemlineata被570 nm或585 nm的黄色LED强烈地吸引,但在其他波长的光选择试验中发现了性别差异。相对于660 nm 红色光或351 nm 紫外线,雄虫更喜欢白色LED,而雌虫则没有这种选择性倾向。虽然相比红色LED,雌虫更喜欢紫外线和472 nm蓝光,但雄性这种倾向不明显(Otálora-Luna and Dickens, 2011)。
对芬兰的夜蛾科Hydraeciapetasitis进行野外的标记再捕法的调查结果表明,同一种群中特定的个体有的容易被光吸引,意味着物种内有可能存在对光的行动变异(Väisänen and Hublin,1983)。根据趋光性与长距离移动的相关性,估计这种物种内变异的原因之一是与翅型多型的关联。但是,中印玫瑰蚜Sitobionrosaeiformis的有翅型和无翅型的两种形态都观察到了正的趋光性,没有观察到与翅型多型相关的显著趋光性差异(Hajong and Varman,2002)。
3 引起趋光性的光和光源
昆虫的趋光性是由具有什么特征的光源引起的呢?昆虫通过复眼和单眼接受波长(颜色)、强度、偏振等光的属性,或者光源的大小和形状等视觉属性,可以认为它们是对各种光和光源的视觉属性的行动反应。
从使用了各种各样种类灯的野外调查来看,汞灯吸引了很多种类和个体数(Mikkola,1972;Eisenbeis,2006;Batesetal., 2013)。但是,由于各种灯的光和光源的属性有很大的不同,为了弄清昆虫是由什么样的属性所吸引的,需要试验来控制光和光源的各种属性,观察只有特定的视觉特性发生变化时的行为反应。
3.1 波长
显然,在光的属性中,波长,即颜色属性对趋光性有很大的影响。例如,很多试验昆虫对波长不同的光源有明确的偏好(Weiss,etal., 1943; Weiss,1944,1946),在这些试验中,尽管刺激光的强度和试验昆虫个体的适应状态有所不同,个体的年龄、性别、观察的行动等条件各种各样,但是,对于大多数昆虫来说,330~370 nm的紫外线的诱导性是最高的(Menzel,1979)。
Labhart(1974)通过严格的试验,从几种昆虫中获得了趋光性的作用光谱。在西方蜜蜂Apismellifera中,用各种波长照射固定个体的一侧的复眼,将头部朝向照射方向的反应作为趋光性的指标,得到了趋光性光谱灵敏度曲线(Spectral sensitivity curve),在蓝色区域(438 nm)和绿色区域(534 nm)进行了确认。使用能够自由行走的西方蜜蜂,通过自发的趋光性行为(Spontaneous photactic behavior)选择光源的试验也得到了在紫外线区域具有高峰值的类似结果。
通过高压氙灯的白光和各种光强度的单色光进行趋光性选择的试验,研究黑腹果蝇Drosophilamelanogaster的作用光谱(Schümperli,1973)表明,在低光量条件下0.05 ergs-1·cm-2(1 ergs-1·cm-2=1×10-3J·m-2·s-1)中,在紫外线区(367 nm)和绿色区(504 nm)获得了同样高选择性的双峰性曲线,但是在高光量条件(10 ergs-1·cm-2)下,其对绿色区域的选择性大大降低,被强烈地吸引到紫外线区,在黑腹果蝇中,根据复眼的适应状态,作用光谱也会发生很大的变化。
黏虫Mythimnaseparata在室内条件下对绿光表现出较强的趋性(Kimetal., 2019),但是在野外,绿光很少能诱集到这种蛾子,而紫外灯和探照灯(卤素灯)可以诱集到大量黏虫蛾子。这种蛾子是否像黑腹果蝇一样对绿光的趋性因光量条件变化而变化,是在监测设备研发中需要注意的问题。
3.2 光强度
引起趋光性的最低限度的光强度,在应用上也很重要。在暗箱中进行食物训练的西方蜜蜂,在得到饵料后,为了归巢从暗箱中逃脱而显示出趋光性。此时,537 nm的绿色LED,8.3×107quanta/cm2/s(1 quanta/cm2/s=1 photons·cm-2·s-1)以上的光强度显示反应,根据蜜蜂的小眼的视角和绿色受体的数量计算,这意味着 7只小眼的28个绿色受体是1 s内同时接受10个光学量子就发生反应的、非常高的灵敏度阈值(Menzel and Greggers,1985),强度响应曲线(Intensity-response curve)明确了昆虫的趋光性反应是如何变化的,因此通常光源的光强度较大,昆虫趋光性被认为是很强地激发出的。但是,光强度越大,是否会引起更强力或更准确的趋光性就越值得研究探索。在黑蝇科的一种红头丽蝇C.erythrocephala中,使用10-3~103 lx(1 lx=11 umen·m-2)的卤素灯,对光源的趋光性精度进行了调查。结果,在102 lx中观察到最准确光源的定位,在光强度103 lx以上的情况下,对光源的定位分散(Meyer,1978)。即使是二化螟,在确定光源时也暗示有最佳的光强度,Kaburaki and Iwasa(1933)用矩形暗箱的一端放置白炽灯泡的实验场所,调查了从1.9×10-5lx到17.2 lx的照度变化,1.3~9.2×10-2lx的照度下雌雄虫都显示出最大的抵达率。
3.3 偏振光
水生昆虫以及水生昆虫演化而来的昆虫(如麦红吸浆虫Sitidiplosismodellana)可被直线偏振光诱集(Schwind,1991;蒋月丽等,2016),但这些昆虫的偏振走势主要通过人工控制水面反射光的偏振度来阐明,昆虫也可被偏振照射的光源所吸引,或回避这种光源。
白纹大凤蝶Papilioaegeus的雌蝶,通过偏振滤光器将偏振方向改变后的两个光源垂直竖立,选择其作为产卵基质,对于水平方向偏光的光源,显示出活跃的产卵行为(Kelber,1999;Kelberetal., 2001)。另外,据报导埃及伊蚊Aedesaegypti雌蚊,可产生对趋向光源偏振光。这个试验是在透明水槽的正下方配置了15 W的黑灯和20 W的白热卤素灯,通过更换放置在水槽和光源之间的扩散板和偏振滤光器的顺序,产生偏振光和非偏振光照射,对水槽内偏振光和非偏振光选择,发现雌虫更喜欢水平线性偏振光照射的水槽中产下更多的卵(Bernáthetal., 2012)。
在野外的调查中,观察到与非偏振光与垂直线性偏振光相比,双翅目摇蚊科更容易被水平线性偏振光吸引(Danthanarayana and Dashper, 1986)。
另外,在使用1 000 W水银灯的调查中,观察到偏振光源诱捕器中捕获到了更多双翅目和鳞翅目个体数量(Kovrov and Monchadskiy,1963)。使用42 W U形荧光灯的偏振和非偏振光分别作为光源的诱捕器,捕获了鞘翅目的115种昆虫,其中8种昆虫被偏振光源捕获到了更多的个体(Szentkirályietal., 2005)。
3.4 光源的大小
关于光源大小对昆虫趋光性影响的研究出乎意料地少。在以欧洲玉米螟Ostrinianubilalis为对象的野外诱集试验中,使用了以750 ft-cd(1 ft-cd=10.76 lumen·m-2)的光强度250 W的氙气光源。将圆形发光面的直径从50.80 mm改变到143.76 mm,面积最大8倍的4个光源尺寸的诱捕器,统计的诱捕虫数,随着发光面的尺寸增加了8倍,诱捕虫数增加了约5倍(Ficht and Hienton,1941)。另外,关于市售的电击光诱捕器的21种型号,家蝇Muscadomestica的捕获率相关的光源亮度、紫外线的反射面积、光源瓦数、诱捕器形状、颜色、尺寸等6个因素进行重回归分析,明确了光源的亮度、紫外线的反射面积对捕获率有很大的影响(Pickens and Thimijan,1986)。
3.5 光源的形状和图案
在趋光性对形状的选择性研究中,都使用色板等反射光。另一方面,在作为直接光的光源中,关于形状和图案是否有选择性,以及是否有与反射光同样的选择性等还不太清楚,今后进行的研究有望有应用性进展。
在使用反射光的试验中,柑橘蓟马Sciertothripscitri对三角形、方形和椭圆形的(Morenoetal., 1984)、3种果蝇对方形和圆形的(Hill and Hooper,1984)、西花蓟马Frankliniellaoccidentalis对圆形的(Mainali and Lim,2010)、烟粉虱Bemisiatabaci对三角形的(Kim and Lim,2011)颜色板具有很强地趋性。但是双翅目科蝇类(Bradbury and Bennett,1974)和芫菁科的Hycleasapcicornis(Lebesaetal., 2011)的色板试验中,报告显示对光源形状的选择没有偏好。另外,在西方蜜蜂选择各种黑白图案的试验中,蜜蜂显示出生来就喜欢放射状对称的图案(Lehreretal., 1995)。
3.6 光源的频闪
人类能感知光源频闪(或频振)的极限闪烁频率大约为50 Hz,超过这个界限以上的频闪在一起看起来像连续的光。已知一些飞行昆虫频闪频率高,使得人类无法察觉(Autrum,1950,1958;Syms and Goodman,1987)。在家蝇的测试中,选择平均光强度和光谱一致的闪烁频率100 Hz 15 W的紫外线荧光灯和无闪烁的荧光灯,结果表明75%的个体被100 Hz频闪的灯吸引,光源的频闪表现出高的诱集效果。与无闪烁光源相比,在使用13 W白色小型荧光灯的试验中,粉虱科的烟粉虱B型、温室白粉虱Trialeurodesvaporariorum,纹翅粉虱Trialeurodesabutilonea(Chuetal., 2004)和热带家蚊Culexquinquefasciatus以及家蝇(Chuetal., 2006)频率为120 Hz的闪烁光源吸引了更多昆虫个体。无闪烁光源也有显著吸引的倾向(Chuetal.,2006)。
猫栉首蚤Ctenocephalidesfelis对频闪的反应室内试验中,使用510~550 nm的LED灯的用市售光诱捕器,频闪光诱捕器显示捕获数量高出连续光源5倍(Mülleretal., 2011)。在半翅目花蝽属Oriustristicolor的试验中,观察到了照明下的步行距离和转向频率。与用直流电源和电容器降低闪烁的100 W白炽灯泡相比,在闪烁频率为120 Hz的白炽灯泡下,小花蝽的步行距离显著变长,步行中的转向频率变少(Shields, 1980)。
光源闪烁间隔较长的试验中,拟步行虫科Tenebrionidae的杂拟谷盗T.confusum和赤拟谷盗T.castaneum(Herbst)的成虫,以及皮蠹科Dermestidae黑绒皮蠹Attagenusmegatomad的幼虫,在以1~15 000次/min的频率使闪光灯发光的情况下,在100~200次/min的频闪频率以下,光源对拟步行虫科的成虫的诱导性显著降低,相反黑绒皮蠹的幼虫,在100次/min的频率以上,显示出负趋光性的个体数的比例上升了(Soderstrom,1968)。
3.7 光源的高度
从野外(Ficht and Hienton,1941;Herms,1947;Frost,1958;Braverman and Linley,1993)和室内(Mboeraetal., 1998)的研究中得到了光陷阱的高度影响昆虫的结论,短库蠓Culicoidesbrevitarss被白炽灯泡的诱捕器所吸引,而将18 W的灯泡分别设置在2 m、4 m、6 m的高度时,许多短库蠓C.brevitarss被从地表引到2 m的光诱捕器(Murray,1987);也有报告说光源的高度不影响诱捕性(Keever and Cline,1983)。在调查这些光诱捕器的高度和诱捕率的关系时,可能是原本昆虫飞翔的高度不同,或者是光源的视认性不同(Herczig and Mészáros,1994)。另一方面,根据昆虫用哪个仰角捕捉光源,就可以得出诱虫数量变化可能结果(Pickens and Thimijan,1986),光源的仰角属性,今后还有进一步验证的价值。
3.8 光源的发光方向
在不是点光源的情况下,有人指出发光面的方向(Orientation)可能影响诱集数量。在向家蝇Muscadomestica开展40 W黑光灯源的室内试验中,将光源的长轴水平放置,是垂直配置的2.6倍引诱效果(Pickens and Thimijan,1986)。
4 昆虫趋光性机制和适应性意义
昆虫如何接受光和光源的属性、处理信息,并通过运动接近光源,这种趋光性的行动机制,以及接近光源的性质对昆虫有什么好处等等,关于趋光性的适应意义,迄今为止提出了许多假说,其中一些经过各种验证而被称为理论。在关于趋光性的假设中,现在最有力的是指南针/罗盘理论、马赫带理论、以及开放空间理论。但是,无论哪一种理论,都不能充分说明广泛物种中的趋光现象和迄今为止观察到的趋光性特征。
4.1 罗盘理论
以鳞翅目蛾类昆虫为主,很多昆虫将天空的自然光源作为长距离移动时罗盘的基准点(compass reference)来使用(Buddenbrock,1917;Baker,1978;Wehner,1984)。在长距离移动时,昆虫将月亮和太阳作为确定方向的视觉目标,将灯火等夜间人造光源误认为月亮和太阳,从而引起对光源的外观上的引诱。罗盘理论认为,通过保持一定的定位角度来固定体轴,昆虫是直线移动的。在这种情况下,如果误认人造光源为太阳或月亮的昆虫对相对接近的人造光源作为记号,则定位角度(α),昆虫为了保持相对于记号的定位角度而不断修正体轴,以光源为中心螺旋状飞翔(Baker and Sadovy,1978);据预测,昆虫会在火焰和人造光源的周围以螺旋的形式飞行,这种罗盘理论能更好地说明昆虫会在火焰和人造光源的周围飞行的事实,这是Nature杂志刊登的Baker and Sadovy(1978)对欧洲地老虎Noctuapronuba引诱光源特征的报道。月球表面大小的视直径为0.518°,在试验中使用的125 W水银灯和月亮大致相同大小的地方,蛾类被吸引走了,他们认为人造光源很可能被误认为月亮。
通过罗盘理论来解释多种昆虫的趋光现象,还存在几个疑点。其中一个是维持定位角度的问题。在罗盘理论中,图2的定位角度α像这样,相对于光源,以个体、个体群、物种等为单位,分别具有特定的定位角度。定位角度越是锐角,昆虫就越被人造光源直线吸引,90°时,昆虫在保持与光源等距离的同时在周围旋转,在90°以上的情况下,可以从光源飞出去(Boyadzhiev,1999),个体应该显示相同的定位角度。但是,当被人工光源吸引,观察在光源周围旋转的昆虫时,发现同一个体在很短的时间内多次反转旋转方向。这种定位角度短时间内的变化,指南针理论解释不出来,以自然光源的定位角度和对人造光源表示的定位角度相同为前提,唯一验证这一点的研究是Sotthibandhu and Baker(1979),同样以欧洲地老虎为材料,以月亮为指南针的基准点使这头蛾子飞行——使用室外悬挂飞行装置,对月球和125 W的水银灯进行了飞翔试验,向以20°到+1°的定位角度定位的6头蛾提供了人工光源,结果表明每头蛾在光源上的定位角度几乎和月球显示的相同。
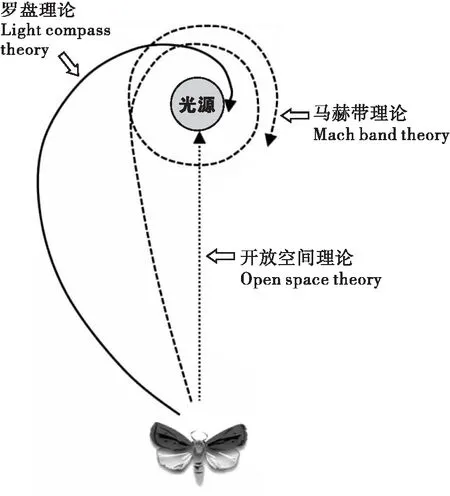
图2 蛾子向光飞行轨迹的三种定向理论(仿Spencer et al.,1997)
4.2 马赫带理论
在马赫带(Mach band)理论中,昆虫被吸引到光源附近,是因错觉引起的逃避到最黑暗方向的定位行为,明暗对比现象的一种马赫带(Mach bands)和切夫勒错觉(Chevreul illusion),在人的视觉中也能观察到,在不同的亮度之间对比度更明显,是可以作为轮廓捕捉的现象,与明度高的部分接触的黑暗部分,在心理上比实际要阴暗。引起明暗对比的神经机构,是由节肢动物的鲎复眼发现的,被称为侧抑制(Hartlineetal., 1956)。昆虫也有侧抑制机构,所以和人类一样,与明度高的区域接触的黑暗区域被认为是更暗的。因此,光源明亮部分的旁边的黑暗比周围的黑暗更加暗,打算逃避在黑暗中的昆虫,相反地在光源周围直线定位。
Hsiao(1973)利用巧妙的实验装置,分析了美洲棉铃虫Helicoverpazae对光源的移动轨迹,将蛾子固定在实验池水面的发泡苯乙烯小船上,通过翅膀扇动来记录游船的轨迹,定量分析了蛾子的移动轨迹和到达位置。对于悬挂在水面正上方的8 W黑光灯,蛾子直线移动到光源正下方,并在其周围旋转。另一方面,在实验池的壁面配置了15 W的黑光,在形成垂直明暗边缘的情况下,蛾子相对于边缘大约直线移动,并且很多蛾子到达明暗边缘的正侧面的暗部,一部分蛾子到达光源侧的明亮部分。同时Hsiao表示,在野外垂直设置15 W的黑光灯,在其旁边配置涂有粘接剂的黑色撞击板,观察并记录粉纹夜蛾Trichoplusiani、美洲棉铃虫、甜菜夜蛾Spodopteraexigua到达距离光源多远的位置。结果表明,无论在哪种情况下,在离光源约15~45 cm的位置,都观察到了黏着虫数的最高值。
4.3 开放空间理论
狭义的趋光性也包含罗盘定位理论,但是,趋光性表面意思是直接定位到光源明亮的部分,属于“真正的趋光性”。认为昆虫定位在光源明亮部位的假说被称为开放空间理论(图2),昆虫认为人工光源是开放空间的指示而被捕捉(Mazokhin-Porshnyyakov,1960)。落叶之下或林冠覆盖的树林内的昆虫在逃离它们的封闭空间时,必须朝向天空等明亮的部分飞去。被人工光源吸引的昆虫是以开放空间为目标移动的,这是真趋光理论解释。
美国白蛾Hyphantriacunea羽化后不久的成虫,被观察到了向树冠上空开放的夜空飞翔的行为,被指出是由于趋光性而定位在微光的夜空中(Hidaka,1972)。长小蠹Platypusquercivorus(Igetaetal.,2003)和罗汉松蛀皮天牛Hirticlytuscomosus(Sato,2005)等森林的钻蛀性昆虫类,已经确认成虫通过趋光性,以定位在森林内存在的间距和林地边缘等开放空间。
4.4 其他假设
由于人工光源的红外线成分与某种蛾类雌性激素的红外辐射相似,所以有人提出生物天线假说,蛾类会误以为光是交配对象的雌性而被吸引(Callahan,1977),一方面,Robinson and Robinson(1950)主张,蛾类在光源周围飞来飞去的行为是被光源捕捉到的行为,解释为通常的行动受到阻碍而引起向光源侧弯曲的反应(Robinson,1952)。但是,这些假说可以说明一些昆虫的物种趋光性,而不足以说明广泛的昆虫趋光性现象,包括这个假说的前提在内,试验验证是不够的。另外,也有昆虫趋光行为的光胁迫假说,受光源照射后个别物种的一些生理响应或某些基因表达以及生命过程的影响(桑文等,2016;Duan等,2017)。
5 昆虫向人工光源移动的轨迹
在阐明趋光性的行动机制上,与到达光源的位置同样重要的是昆虫对光源的飞翔或步行轨迹及其特征,这些是不同的(Spenceretal.,1997)。在罗盘理论中,昆虫在光源上画螺旋状逐渐接近;在马赫带理论中,昆虫在光源的旁边一直定位;在开放空间理论中描绘直线移动到光源中心的轨迹(图2)。但是,由于定量观察困难等原因,不仅在野外观察,在室内试验中也几乎没有正确记录昆虫向光源移动轨迹的研究,Hsiao(1973)使用上述的小船的美洲棉铃虫蛾的试验,比较了对烟草天蛾Manducasexta观察到的定位角度。与5 W 白炽灯泡相比,烟草天蛾的平均定位角度接近-0.22°±2.7°和0°,显示了真正的趋光理论所预期的向光源中心的直线移动。
在这些研究之外的移动轨迹的报告可以说是非常片段的。当远离光源的位置时,某种昆虫会直线地或向Z字形光源移动,在其他物种中也观察到无视光源直接飞行(Robinson and Robinson,1950;Stewartetal., 1969)。在观察到从远距离到光源的接近行为时,根据试验条件和物种,报告了各种各样的结果。例如,Baker and Sadovy(1978)认为,夜蛾科昆虫对两种光源的反应仅在以光源为中心的3 m范围内观察到,因此认为距离过短。也有人认为这是由于试验方法不恰当(Bowden,1982)。此外,125 W的汞灯对昆虫诱导光源的半径为50 m~700 m(Bowden and Morris,1975;Bowden,1982)。
观察到接近光源的昆虫在光源周围表现出复杂的行为,但是以下没有特定的倾向。Robinson and Robinson(1950)指出,光源周围昆虫的飞翔行为,一是在特定的距离内接近光源,一部分围绕光源旋转后,立即飞走的行为;另一种是,它是一种边摇摇晃晃地飞翔边接近光源,然后边以陡峭状的曲线一边加速接近光源,然后穿过光源边缘再次接近光源的反复行动。据报道,受野外光源(诱捕器)吸引的烟草天蛾在接近光源的过程中会在地面上降落或碰撞(Hartstacketal., 1968)。通过对光源周围飞翔的蛾类飞行轨迹进行影像分析,在接近光源时,随着蛾类角速度的急剧增加,会出现迷失定位方向的行为。同样的观察,飞行速度通常在接近光源时是最小的,在离开光源时是最大的(Muirhead-Tomson,1991)。但在观察烟草天蛾的室内试验中得到了相反的飞行速度倾向。以0.4 m/s的速度接近光源的天蛾在从光源飞离时,会以0.29 m/s的速度降低飞行速度(Spenceretal., 1997)。
昆虫相对于太阳和月亮等自然光源的定位行动的研究例子很多(Wehner,1984),但是记录了面向光源行动的移动轨迹的例子不多,盲蝽科Miridae的可可褐盲蝽Distantiellatheobroma在野外释放试验,释放时间是在黎明的时候,观察到了太阳在低高度位置上可可褐盲蝽直线飞行的倾向。另一方面,在太阳来到头顶的正午,观察到了可可褐盲蝽螺旋飞翔和水平曲折飞翔的复杂移动轨迹。
6 结语
在光源周围聚集的昆虫中,有多少是以趋光性为原因聚集起来的?那么,什么样的趋光性行为机制将昆虫引导到光源上呢?我们还在寻找答案的阶段,对趋光性的理解更加困难。
到目前为止,昆虫趋光性的研究,即对于放置在野外或实验室内的人工光源的昆虫诱导和回避的研究,大多着眼于光源本身的吸引。但是,很多人都直观地联想到的趋光性,昆虫定位在光源明亮的部分,我们已经清楚地认识到,仅凭目前的趋光理论是无法解释广泛的趋光现象的。我们需要针对昆虫对人造光源的到达位置和移动途径或轨迹,以许多物种为对象,进一步进行野外的调查研究,在受控实验室内,应仔细验证在野外观察到的到达位置和移动轨迹是否可再现,如Shields(1989)指出的,昆虫和人类的眼睛有很大的不同,需要严格控制用于实验的人工光源的光和光源的属性,可以容易控制波长和光强度等光属性、大小和形状等光源的视觉属性的LED照明的普及,将促进今后昆虫对光的行为反应进行研究。
对昆虫的趋光性和趋光性现象的进一步理解,将会改进和开发包括更高诱虫性的光诱捕器在内的视觉诱捕器,基于在粘着式光诱捕器、灯光采集、水盘诱捕器等各种领域中使用的丰富多彩的视觉诱捕器(Shimada,2018),是一条新形式的专性诱虫性光源开发之路。而且,更加让我们感到对生物行为的深奥之处的惊叹,那就是,在我们身边的生物现象中还残留着一个巨大的谜团,再一次被趋光现象所吸引,并警示我们要开展更加深入的研究。
猜你喜欢
玩具世界(2022年3期)2022-09-20
导航定位与授时(2020年5期)2020-09-23
甘肃教育(2020年2期)2020-09-11
铁道通信信号(2020年9期)2020-02-06
小学生优秀作文(低年级)(2018年9期)2018-09-10
知识经济·中国直销(2018年3期)2018-04-12
小学生必读(低年级版)(2018年11期)2018-03-13
数学小灵通·3-4年级(2017年3期)2017-04-16
上海农业学报(2017年3期)2017-04-10
学习月刊(2015年1期)2015-07-11
