
橘小实蝇对不同品种苹果的选择偏好及适应性
2023-10-24 05:41李星星周国富骆官雨陈思蓉张金龙陈国华张晓明
中国农业科学 2023年17期
李星星,周国富,骆官雨,陈思蓉,张金龙,陈国华,张晓明
橘小实蝇对不同品种苹果的选择偏好及适应性
李星星,周国富,骆官雨,陈思蓉,张金龙,陈国华,张晓明
云南农业大学植物保护学院/云南生物资源保护与利用国家重点实验室,昆明 650201
【目的】探究橘小实蝇()在6个主栽苹果品种果实上的选择偏好,了解不同品种苹果果实对橘小实蝇生长发育的影响。【方法】以3种早熟品种(华硕、红露、嘎拉)和3种晚熟品种(短枝富士、烟富三号、秦脆)为试验材料,使用硬度计和糖度折光仪测定苹果果实的硬度和糖度;设置完整果实、去皮果实和果实气味3种处理方式,将同成熟批次的同一处理方式果实放入养虫笼内,测定橘小实蝇对不同品种果实的访问数和产卵数;饲喂初孵幼虫不同品种果实,测定其取食后幼虫历期、化蛹率、蛹重等生长发育参数,分析其对不同品种苹果的选择偏好和生长发育适应性。【结果】橘小实蝇在不同品种苹果果实上的访问数和产卵数存在差异,雌虫对果实的不同品种和处理更为敏感。早熟品种的完整果实中,橘小实蝇更偏好选择嘎拉,该品种上的雌虫访问数和诱卵数分别达9.0头和105.0粒/果;晚熟品种的完整果实中,橘小实蝇更偏好选择秦脆,该品种上的雌虫访问数和诱卵数分别达5.5头和64.2粒/果。果实处理方式影响橘小实蝇的选择偏好,除嘎拉外,同一品种果实的不同处理方式中去皮果实上的访问数和产卵数更高;早熟品种中以华硕的去皮果实上诱卵数最高,达201.6粒/果,晚熟品种中以烟富三号的果实气味诱卵数最高,达83.8粒/装置;双因素方差分析发现,品种和果实处理方式会对访问数和诱卵数产生影响。橘小实蝇取食嘎拉后的幼虫历期最短,为11.9 d,蛹期以华硕最短,为9.9 d;化蛹率和羽化率在晚熟品种中更高,橘小实蝇幼虫取食不同品种苹果后的特定年龄-阶段存活率差异主要在2龄阶段,取食华硕和嘎拉的2龄幼虫发育速度最快,3 d时达到特定年龄-阶段存活率峰值。【结论】橘小实蝇对苹果不同品种的选择偏好存在差异,早熟品种中偏好选择嘎拉,晚熟品种中偏好秦脆;果实完整程度和气味对其选择偏好有影响,去皮果实和果实气味上的访问数和诱卵数普遍高于完整果实;橘小实蝇能在苹果果实上完成幼虫期的生长发育,具有在苹果园中繁殖危害的能力,橘小实蝇对苹果果实的选择偏好与后代的生长发育适合度关联性不强,同“选择性与适合度”假说存在差异。苹果园橘小实蝇综合管理中应当加强对嘎拉和秦脆品种的受害情况监测,农事操作中避免损伤果实并及时摘除受损果实。
橘小实蝇;苹果;生长发育;选择偏好;选择适应性
0 引言
【研究意义】橘小实蝇()又名东方果实蝇,隶属双翅目(Diptera)实蝇科(Tetriphitidae)果实蝇属(),是世界性重要农业入侵性有害生物[1]。橘小实蝇主要通过成虫穿刺果皮产卵于果实内,卵孵化后的幼虫进行取食危害,幼虫基本在受害果实内生长发育,不转果危害。其幼虫摄入的营养来源于雌成虫产卵时选择的寄主,因此橘小实蝇亲代的选择偏好和子代生长发育的关系,是研究寄主植物对于橘小实蝇的影响、橘小实蝇与寄主植物协同进化的核心内容[2-3]。开展此研究在一定程度上有利于进一步掌握橘小实蝇在不同寄主上的发生规律,对橘小实蝇防治措施的制定具有重要意义。【前人研究进展】橘小实蝇原产于亚洲热带和亚热带地区[4],现已在我国云南、四川、贵州、广东等省均有分布,其寄主范围广泛,可危害芒果、香蕉、苹果等46科250余种植物[5-9]。橘小实蝇成虫具有较高的活动能力,7 d累计飞行距离最远可达97 km,可在不同寄主间转移危害[10]。不同种寄主的不断出现与成熟为橘小实蝇种群的连续危害提供了条件,使其成为果实生产中的重要害虫。在杨桃、番石榴、柿等产区的调查中橘小实蝇的危害率达40%以上,发生严重时产量损失可达80%以上,甚至绝收[11-13]。橘小实蝇还是果园实蝇科害虫中的优势种,在贵州省安龙县果园调查中占比最高达94.85%[14]。由于橘小实蝇的巨大破坏性,欧洲植物保护组织(EPPO)在2016年将其划分为A1类限定性有害生物[15]。橘小实蝇虽寄主范围广泛,但对于不同寄主会表现出明显不同的产卵偏好性。在番石榴、酸石榴、甜石榴和番茄中,其更喜欢在番石榴上产卵,在番茄上的产卵量最低[16];刘慧等研究发现橘小实蝇对5种寄主果实的产卵选择嗜好顺序为杨桃、香蕉、木瓜、甜橙、番茄[17];黄爱玲等研究了橘小实蝇对14种寄主果实的产卵偏好,发现在不同寄主上的产卵量存在差异[18]。在同一寄主的不同品种中,橘小实蝇雌虫的产卵偏好也存在明显差异;在禄子、光颜、沙子、酸甜和厚皮5种石榴品种上,橘小实蝇偏好在禄子和光颜品种上产卵[19];在4种桃品种上,橘小实蝇的产卵嗜好顺序为川中岛>大久保>岱妃>中蟠11[20];黄慧欣等研究发现,橘小实蝇偏好在普红、白肉桂热心和台湾大红品种火龙果上产卵[21]。除了影响橘小实蝇的产卵选择外,寄主植物果实作为橘小实蝇的食物来源,对其生长发育、存活乃至整个种群的生长动态具有重要影响。袁彬乔等选用石榴、苹果、梨、猕猴桃、桃和枣6种寄主果实作为橘小实蝇幼虫的食料,发现在不同寄主上幼虫的生长情况不同,石榴上的内禀增长率最高[22];以东魁、荸荠种和水梅3个品种杨梅果实为食料的橘小实蝇生长发育参数存在差异,其中以取食荸荠种杨梅的橘小实蝇发育最好,幼虫历期最短、初羽化雌虫体重最高[23]。【本研究切入点】近年来,橘小实蝇在贵州、云南、山东等地危害苹果的报道逐渐增加[24-26]。苹果是世界上贸易量最大的主要水果之一,而我国是苹果生产的第一大国[27]。昭通地区苹果种植面积达2.1万公顷,占云南省苹果种植面积的63%,产量达39.6万吨,苹果产业已成为云南省滇东北、滇西北冷凉山区重要的经济收入来源之一[28]。课题组前期调查发现,橘小实蝇已有在昭通地区苹果上危害的情况,目前尚不清楚橘小实蝇对不同品种苹果果实的选择偏好和生长发育适应性。【拟解决的关键问题】测定橘小实蝇成虫在6个昭通地区主要品种苹果果实上的访问数和产卵数,以及幼虫取食不同品种后的生长发育参数,明确橘小实蝇对不同品种苹果的选择偏好和幼虫取食后的生长发育变化,为评价橘小实蝇对苹果的危害风险和苹果生产中橘小实蝇的综合防治提供理论依据。
1 材料与方法
1.1 材料
1.1.1 供试虫源 试验于2022年在云南农业大学云南生物资源保护与利用国家重点实验室的养虫室内开展。橘小实蝇采自云南省昭通市苏家院镇绿健果蔬商贸有限公司(103.55°E,27.28°N)。于田间采集受害虫果后将其带回养虫室,置于相对湿度10%的沙上,待成虫羽化后进行种类鉴定并放入养虫笼(长×宽×高=38 cm×38 cm×38 cm)中饲养,将酵母浸粉与蔗糖等质量混匀饲喂成虫。待其开始产卵后,提供3 cm厚的香蕉片供其产卵,继代饲养5代以上。幼虫饲料参考陈瑶瑶等方法配置,主要由水、香蕉、玉米面、纤维素、酵母浸粉、白砂糖、少量的维生素(Vc、Vb)、叶酸和苯甲酸钠,按照400﹕250﹕250﹕50﹕50﹕50﹕4﹕3﹕1﹕1的比例使用搅拌机打碎混匀制成[29]。饲养及试验条件:温度(26±1)℃,相对湿度为60%—80%,光周期L﹕D=14 h﹕10 h。
1.1.2 供试寄主 供试寄主按果实成熟期分为早熟和晚熟品种,其中早熟品种为红露、嘎拉、华硕,晚熟品种为秦脆、短枝富士、烟富三号,试验按照成熟期分两批次进行。供试寄主均采自绿健果蔬商贸有限公司,选择果实大小基本一致、果面洁净、无病虫和机械损伤的成熟期苹果供试。
1.1.3 仪器与设备 自制长38 cm、高38 cm、宽38 cm的养虫笼(200目纱网);SZ51体视显微镜,日本奥林巴斯株式会社;三量GY-3水果硬度计,东莞三量量具有限公司;BM-0532手持糖度折光仪,深圳东美测量仪器有限公司;自制采卵器,用高6 cm、直径3 cm的透明塑料瓶,自瓶口向下2 cm使用昆虫针均匀打孔制作,孔间隔0.5 cm;自制幼虫饲养盒,用高3 cm,直径5 cm的塑料盒制成,顶部打孔后粘接80目纱网。
1.2 方法
1.2.1 苹果果实硬度与糖度测定 使用三量GY-3水果硬度计测量果实去皮硬度,具体测量方法参考彭帅等,使用8 mm直径探头,压入深度10 mm,以最大值作为硬度测量值,分别测量每个果实的上、中、下部硬度,计算3个位置硬度的均值代表该果实硬度[30]。每品种选择大小一致的果实各5个进行测定。
使用BM-0532手持糖度折光仪测定果实糖度,具体方法参考李慧敏等,取果实的赤道部位果肉,榨成匀浆经纱布过滤后进行测量[31]。每品种选择大小一致的果实各4个进行测定。
1.2.2 橘小实蝇对不同品种及不同处理方式苹果的访问数与产卵选择 寄主处理参考李貌等方法并有所改进,设置3种处理方法:(1)完整果实:将不同品种的果实使用保鲜膜包裹,仅在果实中部将保鲜膜切开一2 cm×2 cm的正方形开口,备用;(2)去皮果实:将不同品种的果实分别削去中部2 cm×2 cm的果皮,并用保鲜膜包裹,将去皮部位的保鲜膜切开;(3)果实气味:取中部果实8 g,切为小块后放入采卵器中[32]。
试验参照Jaleel等[33]方法设置。将处理好的相同成熟期的3种果实分别放入养虫笼中,果实间隔5 cm以上,以是否具有产卵器区分雌雄虫后每养虫笼中放入10对成虫。连续观察11 h,每1 h观察一次,每次2 min,以11次观察中橘小实蝇落在寄主上的数量之和分别计算雌雄虫的访问数。24 h后将寄主取出,记录产卵数。参照周鹏对橘小实蝇交配和产卵节律的研究,试验时间设置:23:00时使用红光灯放入或取出果实,9:30—19:30进行访问数观察,以避免影响其产卵[34]。每处理设置4次重复。
1.2.3 橘小实蝇幼虫在6种苹果果实上的生长发育适应性 参考胡昌雄等生命表测定方法,将盛有苹果匀浆的自制采卵器放置于处于产卵盛期的橘小实蝇成虫饲养笼中,产卵2 h后将卵冲洗过滤收集,选取大小均匀的卵备用[35]。将卵粒放置于铺有湿润滤纸的培养皿(d=7.5 cm)中,培养皿上覆盖保鲜膜。再将准备好的培养皿放置于养虫室,在温度(26±1)℃、RH(70±10)%条件下饲养。每隔24 h使用体视显微镜观察其孵化情况和存活情况(卵发黑变色或失水干瘪视为死亡),将同一天孵化的幼虫挑出放置于幼虫饲养盒进行标号后单独饲养,每日为幼虫更换不同品种新鲜的苹果块(2 cm×2 cm×2 cm)并检查其存活与发育情况。在幼虫即将化蛹时,在养虫罐底部铺3 cm厚度的湿润细沙,供其化蛹,化蛹后称量蛹重。根据橘小实蝇每日存活与发育情况,计算橘小实蝇各虫态的存活率。每品种设置4次重复,每重复25头初孵幼虫。
1.2.4 果实硬度、糖度与橘小实蝇的访问数、产卵数和生长发育参数的相关性 采用Pearson相关性分析法分别分析各品种果实硬度、糖度与橘小实蝇完整果实上的访问数、完整果实上的产卵数和生长发育参数的相关性。
1.3 数据处理
使用SPSS 25.0进行统计分析,采用单因素方差分析比较取食不同品种果实幼虫的各生长发育参数,应用Tukey法进行差异显著性检验(<0.05);采用双因素方差分析比较不同品种及不同处理方式果实上的访问数和诱卵数,应用Tukey法进行差异显著性检验(<0.05);相关性分析采用双尾检验法进行显著性检验。使用Origin 2021绘图。
2 结果
2.1 不同品种苹果果实的硬度与糖度
晚熟品种果实的硬度均高于早熟品种,以短枝富士的硬度最高,为10.1 kg·cm-2,嘎拉的硬度最低,为6.9 kg·cm-2,其余4个品种果实硬度相近(5,18=24.196,=0.0001)。晚熟品种的糖度均高于早熟品种,秦脆的糖度最高,达18.1oBX,嘎拉的糖度最低,为13.1oBX(5,18=21.175,=0.0001)(图1)。
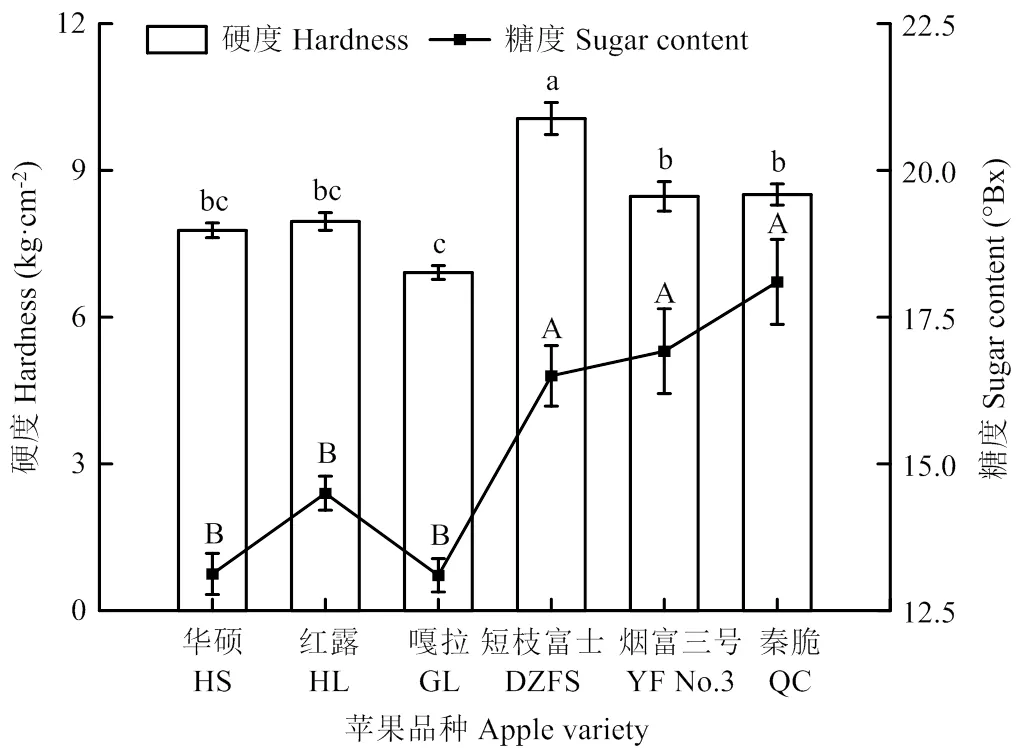
柱上不同小写字母表示不同品种果实间硬度差异显著,折线上方不同大写字母表示不同品种果实间糖度差异显著(Tukey检验法,P<0.05)Different lowercase letters above the column indicate significant differences in hardness among fruits of different varieties, and different uppercase letters above the broken line indicate significant differences in sugar content among fruits of different varieties (Tukey test method, P<0.05)
2.2 橘小实蝇对不同品种及处理方式苹果的访问数与产卵选择
雌虫对苹果品种和果实处理更为敏感,在红露和华硕果实上,均以去皮果实的访问数最高,分别达12.8和11.8头(红露:2,9=25.083,=0.001;华硕:2,9=34.778,=0.0005);完整果实中偏好访问嘎拉,访问数达9.0头,去皮果实和果实气味处理中偏好访问华硕,访问数分别达11.8和11.7头(完整果实:2,9=5.583,=0.043;去皮果实:2,9=14.778,=0.005;果实气味:2,9=5.583,=0.043)。雄虫访问数仅在红露的不同处理果实上存在差异,以完整果实的访问数最高,为9.0头(2,9=7.960,=0.021)。同一品种的同一处理方式果实上,雌雄虫之间的访问数略有差异,绝大部分处理以雌虫访问数更高,在红露去皮果实和果实气味、华硕果实气味上差异显著,分别达12.8、9.8和11.7头(红露去皮果实:1,4=66.125,=0.001;红露果实气味:1,4=16.000,=0.016;华硕果实气味:1,4=45.125,=0.003)(表1)。
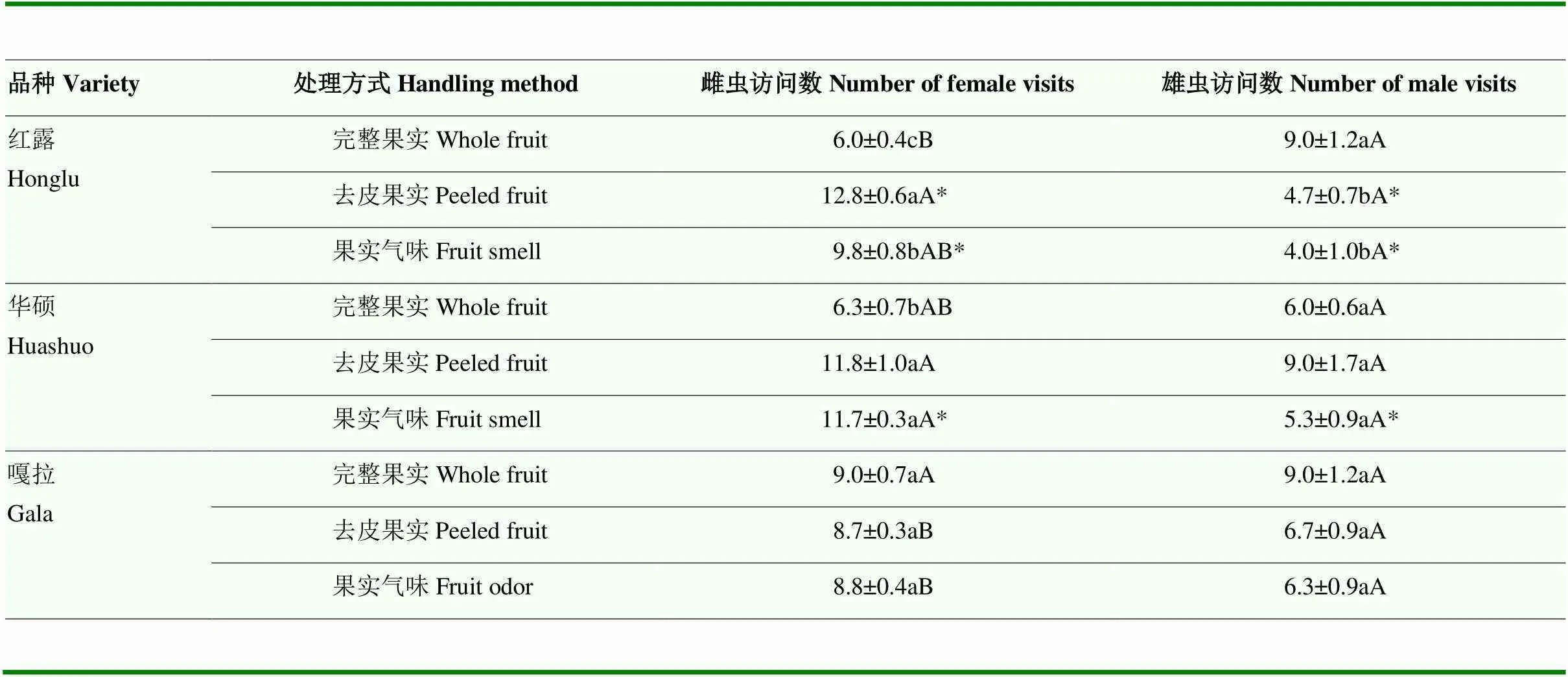
表1 橘小实蝇在不同处理方式的3种早熟苹果上的访问数
数据后不同小写字母表示该性别成虫在同品种、不同处理方式果实上访问数差异显著;不同大写字母表示在该性别成虫在同一处理方式、不同品种果实上访问数差异显著;*表示同一处理方式、同品种果实上雌雄虫的访问数差异显著(Tukey检验法,<0.05)。表2同
Different lowercase letters after the data indicate that the number of visits of the same sex adults is significantly different under the same variety and different fruit handling methods; different uppercase letters indicate the number of visits of the same sex adults is significantly different under different varieties and same fruit handling method; * indicates that the number of visits of male and female adults is significantly different under the same variety and same fruit handling method (Tukey test,<0.05). The same as Table 2
雌虫在秦脆不同处理方式果实上的访问数存在显著差异,以去皮果实上的访问数最高,达9.7头(2,9=6.300,=0.034);不同品种的果实气味上雌虫访问数以秦脆最高,达5.7头,短枝富士最低,为2.8头(2,9=9.125,=0.015)。雄虫在秦脆去皮果实上的访问数最高,为11.3头,完整果实上最低(2,9=37.500,=0.0004)。同一品种的同一处理方式果实上雌雄虫的访问数以短枝富士的去皮果实差异最显著,雌虫访问数更高(1,4=8,=0.047)(表2)。
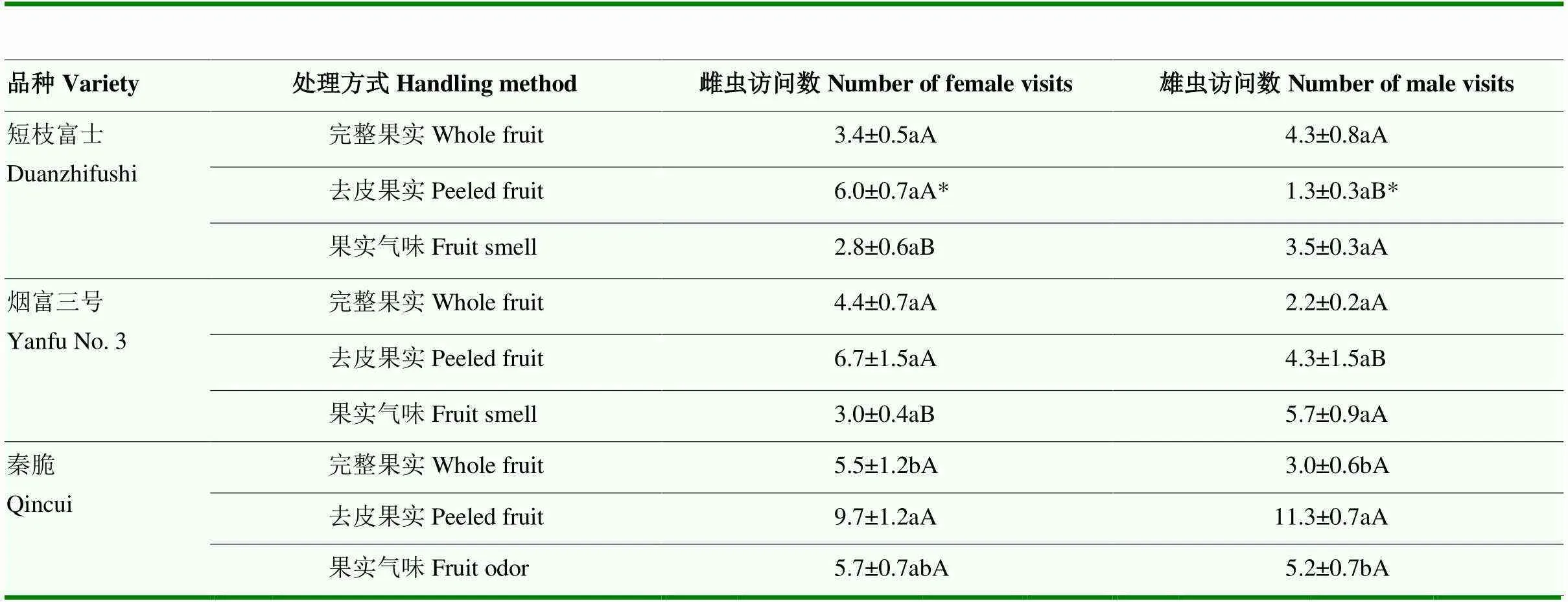
表2 橘小实蝇在不同处理方式的3种晚熟苹果上的访问数
早熟品种对雌虫访问数有显著的交互效应(<0.05),晚熟品种和处理方式单因素对成虫访问数均有极显著的交互作用(<0.01)(表3)。
早熟品种中,华硕以去皮果实上诱卵数最高、红露以果实气味诱卵数最高、嘎拉则以完整果实诱卵数最高,其诱卵数分别达201.6、166.2、105.0粒/果(华硕:2,9=143.519,=0.0001;红露:2,9=90.436,= 0.0001;嘎拉:2,9=10.501,=0.002);完整果实中以嘎拉诱卵数最高、去皮果实以华硕诱卵数最高、果实气味则以红露诱卵数最高(完整果实:2,9=15.054,=0.0005;去皮果实:2,9=129.774,=0.0001;果实气味:2,9=92.227,=0.0001)。晚熟品种中,短枝富士以去皮果实和果实气味诱卵数更高,烟富三号以果实气味诱卵数更高(短枝富士:2,9=61.671,=0.0001;烟富三号:2,9=47.502,=0.0001);完整果实以秦脆的诱卵数最高,达64.2粒/果;去皮果实上秦脆和短枝富士的诱卵数高于烟富三号,在果实气味诱卵数中烟富三号和秦脆的诱卵数高于短枝富士(完整果实:2,9=36.657,=0.0001;去皮果实:2,9=6.499,=0.012;果实气味:2,9=13.248,=0.0009)(图2)。

表3 苹果品种、果实处理方式与橘小实蝇访问数的双因素方差分析
表中数据为值,误差项省略The data in the table arevalues, error terms omitted。*:显著影响significant impact (<0.05);**:极显著影响extremely significant impact (<0.01);NS:相关性不显著Correlation is not significant

柱上不同小写字母表示同品种苹果的不同处理方式果实上诱卵数差异显著,不同大写字母表示同一处理方式的不同品种果实上诱卵数差异显著(Tukey检验法,P<0.05)Different lowercase letters on the column indicate that the number of eggs on the fruits of different treatments of the same variety of M. pumila is significantly different. Different uppercase letters indicate the number of eggs on the fruits of different varieties with the same treatment is significantly different (Tukey test, P<0.05)

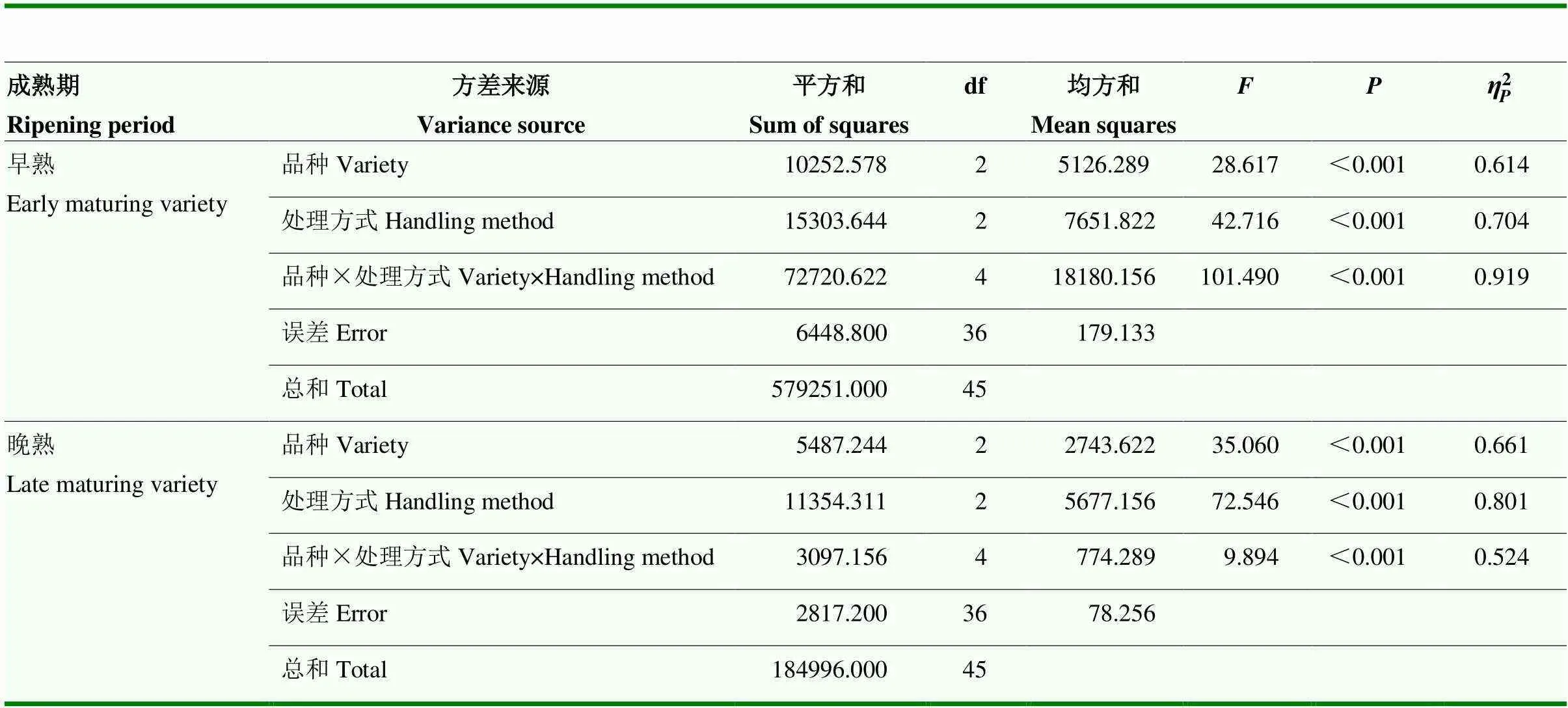
表4 苹果品种、果实处理方式与诱卵数的双因素方差分析
2.3 橘小实蝇幼虫在6种苹果果实上的生长发育适应性
橘小实蝇的幼虫历期在嘎拉果实上最短,为11.9 d,在短枝富士上的历期最长,为13.1 d(5,377=2.88,=0.01)。以烟富三号果实为幼虫期食物的蛹重最重,达12.9 mg,以红露果实为幼虫期食物的蛹重最轻,为11.4 mg(5,352=5.35,=0.0001)。以不同品种果实为食物的橘小实蝇蛹期差异显著,烟富三号>秦脆>短枝富士>红露>嘎拉>华硕(5,353=103.64,=0.0001)。取食短枝富士和秦脆的橘小实蝇羽化率最高,分别达100%和98.7%,显著高于取食嘎拉的羽化率(5,18=3.86,=0.01)(表5)。
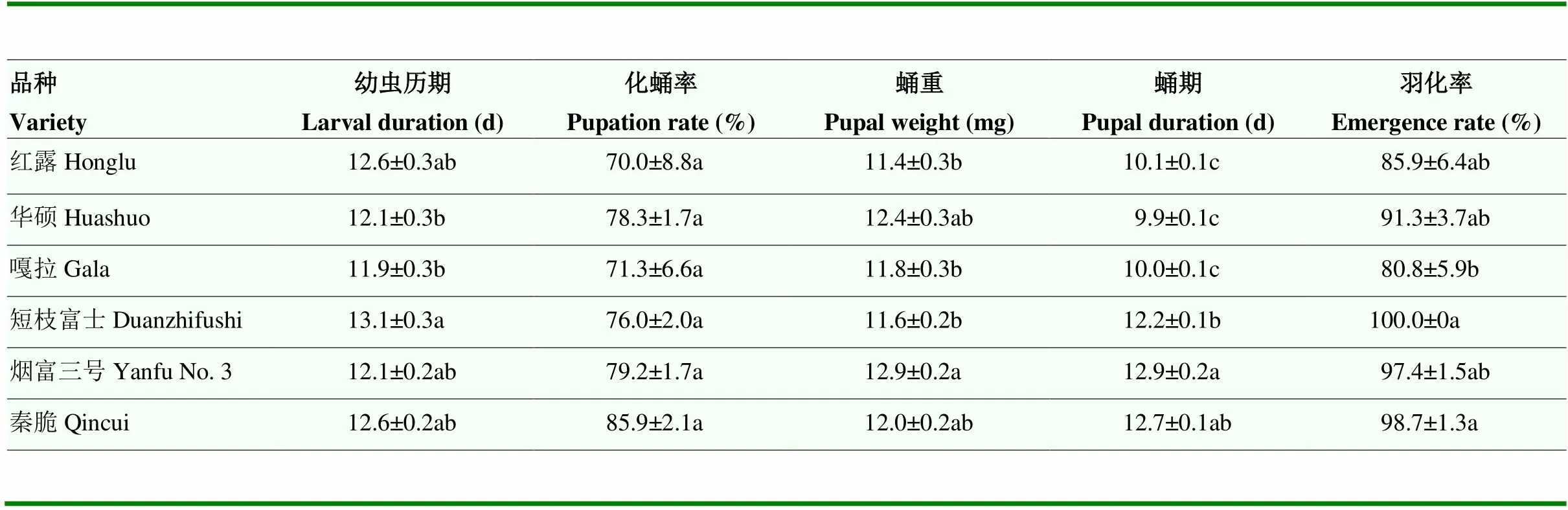
表5 橘小实蝇在不同品种苹果上的生长发育参数
同列数据后不同小写字母表示在不同品种上的生长发育参数差异显著(Tukey检验法,<0.05)
Different lowercase letters after the data in the same column indicate significant differences in growth and development parameters on different varieties (Tukey test,<0.05)
橘小实蝇在不同品种苹果上的特定年龄-阶段存活率趋势基本相同。1龄幼虫存活率均随时间的增加逐渐降低,除取食嘎拉外的橘小实蝇在5 d左右存活率均降至较低水平,而取食嘎拉的橘小实蝇1龄幼虫存活率保持在较高水平,其1龄幼虫历期较长。2龄幼虫存活率随时间的增加先增加后降低,在10 d左右均降为0,但取食不同品种苹果的发育进程不一致。取食华硕和嘎拉的2龄幼虫发育最快,在3 d时便达到存活率峰值,两者的峰值水平不一致,取食华硕的2龄幼虫特定年龄-阶段存活率峰值达80.3%,取食嘎拉的仅为43.9%。取食红露、短枝富士、烟富三号、秦脆的2龄幼虫发育略缓慢,在4 d时达到存活率峰值。取食6种品种苹果的橘小实蝇幼虫的3龄阶段发育较为一致,在7—8 d进入3龄存活率峰值,在10—11 d开始下降,在15 d后降至较低水平。进入蛹期后,达到峰值的进程相似,但峰值水平不一致,取食秦脆的橘小实蝇蛹存活率最低。20—21 d时取食红露、华硕、嘎拉的橘小实蝇蛹存活率开始下降,23 d时取食秦脆、短枝富士和烟富三号的橘小实蝇蛹存活率开始下降。蛹存活率下降后,蛹开始羽化,成虫存活率开始逐渐上升,取食红露、华硕、嘎拉的橘小实蝇于25 d左右达到成虫存活率峰值,取食短枝富士、烟富三号、秦脆的橘小实蝇于30 d左右达到成虫存活率峰值(图3)。
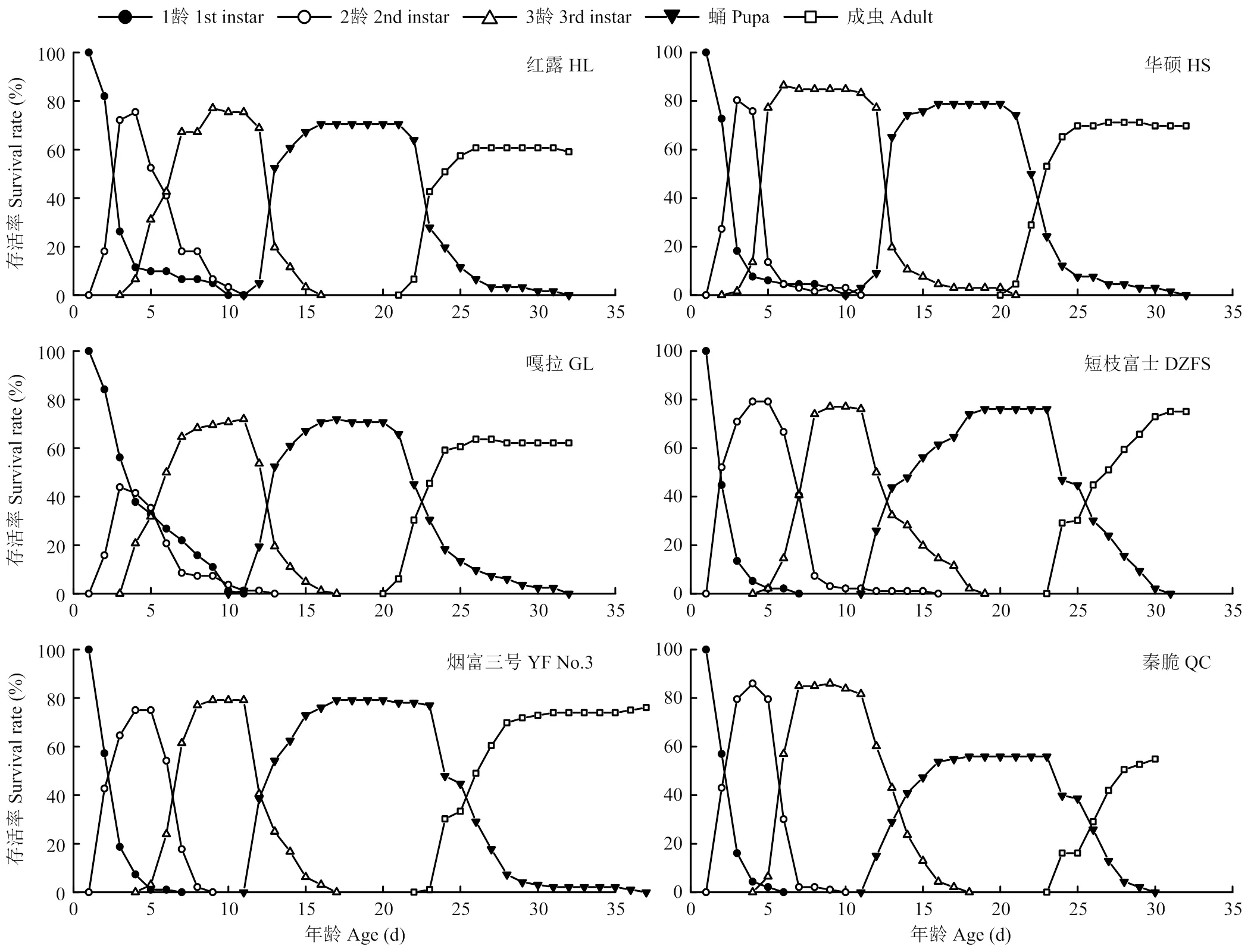
图3 橘小实蝇在不同品种苹果上的特定年龄-阶段存活率
2.4 苹果果实硬度、糖度与橘小实蝇访问数、产卵率和生长发育参数的相关性
不同品种的果实硬度、糖度与橘小实蝇选择偏好相关的雌雄虫访问数、诱卵数均呈负相关,在硬度更大、糖度更高的品种上橘小实蝇的选择性越低:其中硬度与雌虫访问数、雄虫访问数、诱卵数呈显著负相关,相关系数分别为-0.494、-0.427和-0.512,即硬度越大的果实上访问数和诱卵数越低;糖度与雌虫访问数和诱卵数呈极显著负相关,相关系数分别为-0.634和-0.810。不同品种果实硬度、糖度与橘小实蝇生长发育相关的幼虫历期、化蛹率、蛹重、蛹期、化蛹率均呈正相关:其中硬度与幼虫历期呈显著正相关,与化蛹率、蛹期和羽化率呈极显著正相关,相关系数分别为0.433、0.582、0.928、0.670;糖度与幼虫历期、蛹期和羽化率均呈极显著正相关,相关系数分别为0.653、0.681、0.665。选择偏好相关指标同生长发育参数之间基本呈负相关关系:雌虫访问数同蛹期和羽化率间分别呈极显著负相关和显著负相关关系,相关系数分别为-0.556和-0.411;雄虫访问数同蛹重和蛹期之间均呈显著负相关,相关系数为-0.459和-0.424;诱卵数与蛹期和羽化率之间均呈极显著负相关。选择偏好相关指标间均为正相关关系,其中雌、雄虫访问数同诱卵数间分别呈极显著和显著水平,即访问数越多的苹果品种上橘小实蝇诱卵数越多。生长发育参数中幼虫历期、化蛹率、蛹期、羽化率相互间均为正相关,其中蛹期同化蛹率间呈显著正相关、化蛹率同羽化率呈正相关、蛹期同羽化率呈正相关,即蛹期越长化蛹率和羽化率越高(表6)。
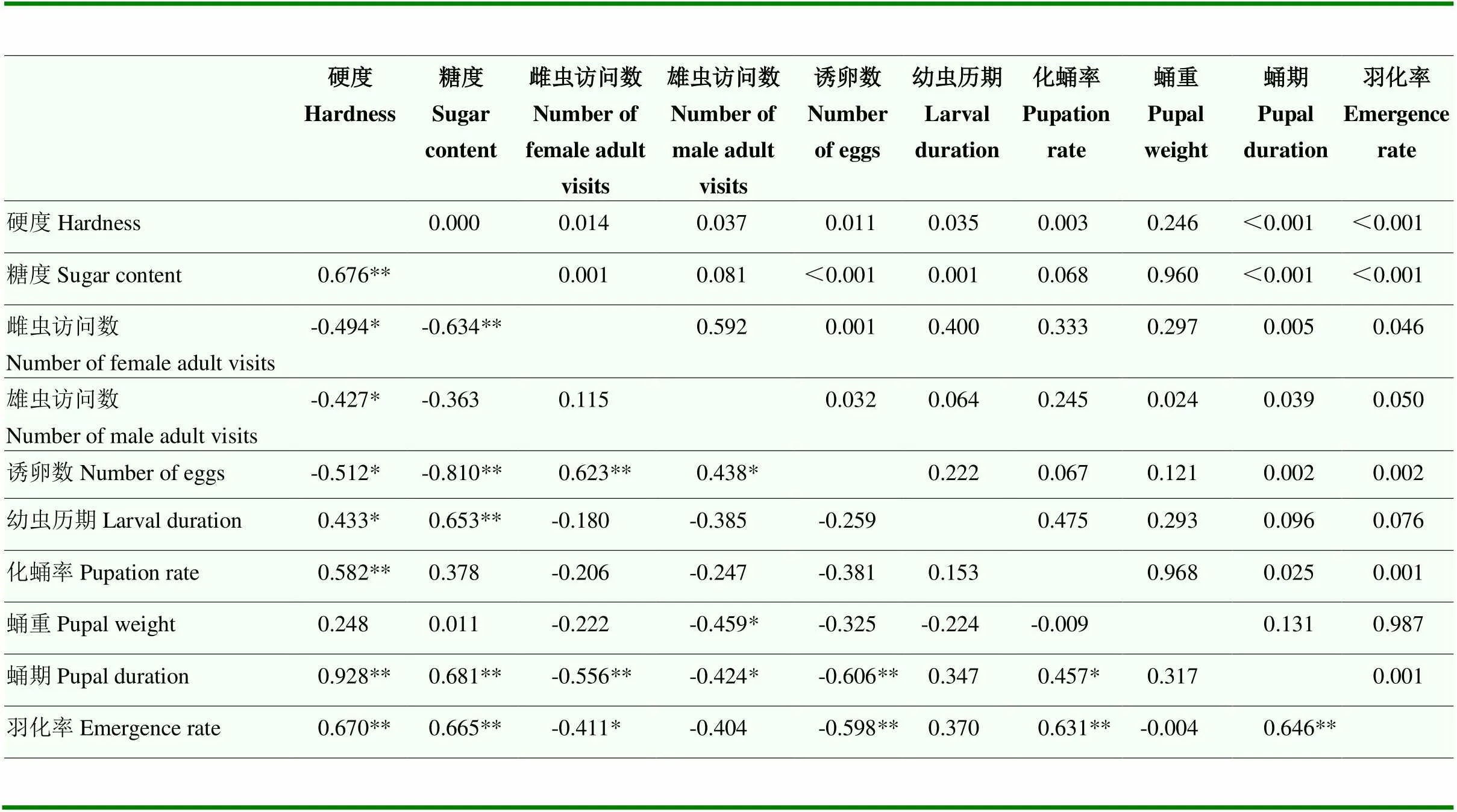
表6 苹果果实硬度、糖度与橘小实蝇取食率、访问数、诱卵数和生长发育参数的相关性
对角线以下数据表示相关系数,对角线以上数据表示相关系数的显著水平Data in the table below diagonal are correlation coefficients, and data above diagonal are significant levels of the correlation coefficients。*:显著水平相关significant correlation (<0.05);**:极显著水平相关extremely significant correlation (<0.01)
3 讨论
3.1 寄主理化性质对橘小实蝇选择偏好的影响
橘小实蝇对苹果果实的选择偏好在不同品种间存在差异:在华硕和秦脆品种去皮果实上的访问数和诱卵数最高,双因素方差分析发现苹果品种和处理方式以及双因素均对诱卵数有极显著的影响,说明苹果品种、果实完整程度、果实气味均为影响橘小实蝇产卵选择的重要因素。橘小实蝇对不同品种的选择偏好不同的现象与已有的多食性实蝇研究结果类似,如黄慧欣等研究了橘小实蝇对5个品种火龙果果实的产卵选择,发现其偏好在普红品种上产卵[21];项前等研究表明,橘小实蝇在不同品种石榴上的产卵存在偏好[36]。植食性昆虫对寄主植物的识别及对不同寄主植物的选择过程中,主要依靠自身的嗅觉、触觉、机械感受和其他可能的信号,因此果实自身的理化性质差异可能对橘小实蝇选择偏好有着重要影响[37-38]。不同品种果实硬度差异可能是产生选择偏好差异的主要原因,蔡子坚等通过通径分析和决策系数计算的方法发现果实硬度是影响橘小实蝇产卵偏好和危害严重度的主决策性状[39];黄爱玲等研究发现,越接近成熟、硬度越低的番石榴上橘小实蝇的产卵率越高[18];李媛等研究表明,橘小实蝇的产卵选择受不同成熟时期三华李果实硬度的影响[40]。本研究结果与已有研究结果一致,硬度最低的品种嘎拉和秦脆上诱卵数最高,果实硬度和诱卵数呈显著负相关关系。不同品种寄主果实的挥发性物质种类和含量存在差异,也会影响橘小实蝇的产卵偏好。如金菊等研究了莲雾的果实挥发物,发现不同品种的莲雾挥发物成分不尽相同,同种物质的含量在品种间也有较大差异,导致对橘小实蝇的诱集效果亦有差异[41];Jayanthi等研究发现2种品种芒果的果实挥发物组成不同,影响了对橘小实蝇的引诱效果[42];张璇等研究发现,草莓果实挥发物的组成不同,对黑腹果蝇()和斑翅果蝇()的吸引效果存在差异[43]。
3.2 苹果果实的完整程度对橘小实蝇选择偏好的影响
与完整果实相比去皮果实上的访问数和诱卵数有所升高,在华硕、短枝富士、烟富三号去皮果实的诱卵数显著高于完整果实,这可能是果皮受损情况下所挥发的烯类等植物挥发物质比正常果实挥发量多,而这些物质可能是吸引橘小实蝇的关键物质[44]。另外,果皮受损后橘小实蝇产卵管刺穿产卵变得更加容易,如UNAHAWUTTI等研究发现,橘小实蝇仅在泰国山竹果皮有裂纹或机械损伤时才能穿刺产卵[45];项前等研究发现,石榴裂果比正常果更能吸引橘小实蝇产卵[36];李貌等研究也发现在去果皮条件下,瓜实蝇()和南亚实蝇()的产卵量显著高于完整果实上的产卵量[32]。红露和烟富三号果实气味的诱卵数显著高于其余两种果实处理的诱卵数,而在其余品种上不同处理方式的诱卵数无明显规律。橘小实蝇落在寄主表面进行产卵活动时,会伴随腹部收缩和上下移动“敲击”果面,还会通过口器舐吸寄主汁液,即橘小实蝇进行产卵选择时会结合果实硬度、汁液、气味等多个因素[46]。不同品种苹果果实的理化性质差异可能导致了不同处理方式下占主要影响地位的因素不一致,呈现出诱卵数在不同品种上大小顺序不一致的现象。在红露和烟富三号上诱卵数均呈现完整果实<去皮果实<果实气味的现象,这两种果实中的某种挥发物质可能对橘小实蝇具有特殊的吸引作用,去皮和切块放入采卵器的处理方法使该挥发物质散发量逐渐增加,但具体是何种物质有待于进一步研究。
3.3 取食不同品种苹果果实的橘小实蝇生长发育差异与适合度评价
橘小实蝇幼虫取食不同品种苹果后的生长发育参数差异显著、特定年龄-阶段存活率略有差异,即不同品种苹果果实的理化性质不仅影响橘小实蝇成虫的选择偏好,还会影响幼虫的生长发育。研究表明,相同饲养条件下取食不同种类寄主的昆虫,其生长发育参数会有所差异。如郭腾达等研究发现,不同品种苹果中的淀粉和可滴定酸对橘小实蝇幼虫体重有显著影响[46];多食性害虫甜菜夜蛾()在芹菜、反枝苋、分葱和青菜上的生长发育参数存在显著差异[47];橘小实蝇在以石榴、苹果、梨等6种寄主果实为食料时,其生长发育参数和种群生命表参数差异显著[22]。在实蝇寄主适合度评价中,采用何种参数作为主要评价依据存在多种观点。有观点采用历期进行评价,认为发育历期短的寄主更适合实蝇,更快的完成生长可以在果实成熟期快速增殖,提前孵化的幼虫在竞争中也更占优势[48];也有研究者采用蛹重进行评价,认为蛹重体现了幼虫期物质的积累,蛹重高的昆虫羽化后成虫的体重更大、耐受不良环境的能力更强[17]。本研究结果显示,以发育历期来看华硕上的蛹期更短、嘎拉上的幼虫历期更短,以蛹重来看烟富三号上的蛹重最高,显然单一指标评价实蝇的寄主适合度可能并不能得出准确、统一的结论。因此,有研究者提出通过统计羽化后的雌虫数与单雌产卵数,并计算净增殖率能更好地反映寄主适合度差异[49]。
目前已有的选择偏好和适合度研究中,以“选择性与适合度”假说为主流的理论,该假说认为雌成虫会偏向于将卵产于适合幼虫生长发育的寄主上[50]。本研究中,早熟和晚熟品种中完整果实上诱卵量分别以嘎拉和秦脆最高,而以此为食料的幼虫生长发育并非最好,将各品种完整果实上的访问数、诱卵数与生长发育指标进行相关性分析,结果显示访问数、诱卵数和蛹期呈负相关,诱卵数与羽化率亦为负相关。有研究发现昆虫选择偏好和后代的生长发育具有正相关性,支持了“选择性与适合度”假说的观点[19,51];也有研究与本研究相似,不支持“选择性与适合度”假说,如刘慧等研究发现番石榴实蝇()和橘小实蝇均偏好在番石榴上产卵,但取食番石榴的幼虫发育的蛹重却明显偏低,木瓜是两种实蝇均不喜好产卵的寄主,但在木瓜上的发育历期却很短[17];张灿等研究表明,木瓜实蝇()在番茄上不产卵,但取食番茄后的幼虫发育的蛹重比其他嗜好寄主更重[52];何超等研究也发现瓜实蝇和南亚实蝇对6种寄主果实产卵选择性和幼虫生长发育适应性相关性不显著[53]。有观点认为,多食性昆虫的产卵选择性和生长发育的适应性之间没有一定的相关性,或者说相比寡食性昆虫多食性昆虫的选择适应性关联更弱[54-55]。苹果是橘小实蝇的非嗜好寄主且在苹果上生长发育情况较差,苹果果实特性(如红露和烟富三号的挥发物质)可能增加了橘小实蝇的访问数和诱卵数,但其他物质含量并不利于橘小实蝇的生长发育,导致本研究中选择性与适应性呈负相关。总之,植食性昆虫的寄主选择适应性仍是一个备受关注、充满争议的问题。如何运用选择偏好指标和生长发育参数,科学合理地评价植食性昆虫的选择适合度还需进一步研究。
4 结论
橘小实蝇对不同品种和处理方式的苹果果实具有不同程度的选择偏好,早熟品种中偏好选择嘎拉,晚熟品种中偏好秦脆,去皮果实和果实气味上的访问数和产卵数普遍高于完整果实。橘小实蝇能在苹果果实上完成幼虫期的生长发育,具有在苹果园中繁殖危害的能力。在果蔬选育中可以将果实硬度纳入选育指标;在果园管理中应及时摘除病果、萎蔫果,清除落果,避免果实硬度降低后橘小实蝇产卵;农事操作时应避免对果实造成机械损伤,定期检查果实被鸟类取食情况,减少田间果皮受损果实数量,降低橘小实蝇通过受损果产卵的机率。
[1] 谢琦, 张润杰. 桔小实蝇生物学特点及其防治研究概述. 生态科学, 2005, 24(1): 52-56.
XIE Q, ZHANG R J. Study advance on biology and ecology of(Hendel) and its control. Ecologic Science, 2005, 24(1): 52-56. (in Chinese)
[2] Costa A M, Amorim F, Anjos-Duarte C S, Joachim- Bravo I S. Influence of different tropical fruits on biological and behavioral aspects of the Mediterranean fruit fly(Wiedemann) (Diptera, Tephritidae). Revista Brasileira De Entomologia, 2011, 55(3): 355-360.
[3] Cunningham J P. Can mechanism help explain insect host choice?. Journal of evolutionary biology, 2012, 25(2): 244-251.
[4] 黄素青, 韩日畴. 桔小实蝇的研究进展. 昆虫知识, 2005, 42(5): 479-484.
HUANG S Q, HAN R C. Advance in the research on the quarantine pest. Chinese Bulletin of Entomology, 2005, 42(5): 479-484. (in Chinese)
[5] 闫振华, 张金龙, 方薛交, 朱文禄, 张文华, 吴革瑛, 张玲, 陶玫, 陈国华. 云南蒙自桔小实蝇寄主种类和枇杷园内桔小实蝇成虫种群动态及其影响因子. 生态学杂志, 2016, 35(11): 3037-3044.
YAN Z H, ZHANG J L, FANG X J, ZHU W L, ZHANG W H, WU G Y, ZHANG L, TAO M, CHEN G H. Host species of the insect pestin Mengzi, Yunnan, and population dynamics ofand its related environmental factors in loquat garden. Chinese Journal of Ecology, 2016, 35(11): 3037-3044. (in Chinese)
[6] 朱俐遐, 李桂珍, 尧美英, 黄丹. 攀枝花地区杧果园桔小实蝇成虫种群动态规律. 中国南方果树, 2019, 48(2): 30-33.
ZHU L X, LI G Z, YAO M Y, HUANG D. Population dynamics ofadult inorchards in Panzhihua area. South China Fruits, 2019, 48(2): 30-33. (in Chinese)
[7] 王涛, 任艳玲, 王珂佳, 杨茂发, 张润志. 兴义桔小实蝇发生动态及影响因子研究. 南方农业学报, 2015, 46(5): 806-812.
WANG T, REN Y L, WANG K J, YANG M F, ZHANG R Z. Occurrence dynamics and influencing factor ofin Xingyi city, Guizhou province. Journal of Southern Agriculture, 2015, 46(5): 806-812. (in Chinese)
[8] 苏湘宁, 李传瑛, 廖章轩, 黄少华, 刘伟玲, 章玉苹. 基于性信息诱捕监测广东桔小实蝇和瓜实蝇发生动态及其对2种常用农药的抗药性评价. 环境昆虫学报, 2023, 45(1): 226-232.
SU X N, LI C Y, LIAO Z X, HUANG S H, LIU W L, ZHANG Y P. Monitoring occurrence dynamics based on sexpheromone trapping and evaluation of two commonly used pesticides resistance ofandin the field of Guangdong. Journal of Environmental Entomology, 2023, 45(1): 226-232. (in Chinese)
[9] AKBAR S A, NABI S U, MANSOOR S, KHAN K A. Morpho- molecular identification and a new host report of(Hendel) from the Kashmir valley (India). International Journal of Tropical Insect Science, 2020, 40(2): 315-325.
[10] 陈鹏, 叶辉, 母其爱. 基于荧光标记的怒江流域桔小实蝇()的迁移扩散. 生态学报, 2007, 27(6): 2468-2476.
CHEN P, YE H, MU Q A. Migration and dispersal of the oriental fruit flyin regions of Nujiang River based on fluorescence mark. Acta Ecologica Sinica, 2007, 27(6): 2468-2476. (in Chinese)
[11] 童伴玲, 黄亦英, 任惠, 陈冠南, 王宏. 茂名市桔小实蝇发生概况及防控对策. 农业与技术, 2020, 40(16): 34-36.
TONG B L, HUANG Y Y, REN H, CHEN G N, WANG H. Overview of the occurrence ofin Maoming City and its prevention and control measures. Agriculture and Technology, 2020, 40(16): 34-36. (in Chinese)
[12] 雷艳梅, 廖冬晴, 黄树长, 谢荔元. 广西番石榴园桔小实蝇发生情况调查及套袋防治试验. 中国热带农业, 2007(2): 50-51.
LEI Y M, LIAO D Q, HUANG S C, XIE L Y. Investigation of occurrence and bagging control experiment ofin guava orchard, Guangxi. China Tropical Agriculture, 2007(2): 50-51. (in Chinese)
[13] 巫辅民, 王小云, 郑霞林, 陆温. 广西柿主产区桔小实蝇的转主为害规律研究. 中国南方果树, 2021, 50(1): 134-140.
WU F M, WANG X Y, ZHENG X L, LU W. Study on the law of turning the main hosts ofin the main producing area of Guangxi. South China Fruits, 2021, 50(1): 134-140. (in Chinese)
[14] 叶照春, 杨德辉, 金剑雪, 李定超, 李鸿波, 程英, 周宇航, 李凤良. 贵州省安龙县杧果园实蝇科害虫种群消长动态分析. 中国南方果树, 2021, 50(4): 87-90, 95.
YE Z C, YANG D H, JIN J X, LI D C, LI H B, CHENG Y, ZHOU Y H, LI F L. Dynamic analysis of population growth and decline of fruit fly pests in theorchards of Guizhou Province. South China Fruits, 2021, 50(4): 87-90, 95. (in Chinese)
[15] Shen G M, Wang X N, Dou W, WANG J J. Biochemical and molecular characterisation of acetylcholinesterase in four field populations of(Hendel) (Diptera: Tephritidae). Pest Management Science, 2012, 68(12): 1553-1563.
[16] 袁盛勇, 肖春, 孔琼, 陈斌, 李正跃, 高永红. 桔小实蝇的产卵选择性. 江西农业大学学报, 2005, 27(1): 81-84.
YUAN S Y, XIAO C, KONG Q, CHEN B, LI Z Y, GAO Y H. Oviposition preference ofHendel. Acta Agriculturae Universitatis Jiangxiensis, 2005, 27(1): 81-84. (in Chinese)
[17] 刘慧, 侯柏华, 张灿, 何日荣, 梁帆, 郭明昉, 武目涛, 赵菊鹏, 马骏. 桔小实蝇和番石榴实蝇对6种寄主果实的产卵选择适应性. 生态学报, 2014, 34(9): 2274-2281.
LIU H, HOU B H, ZHANG C, HE R R, LIANG F, GUO M F, WU M T, ZHAO J P, MA J. Oviposition preference and offspring performance of the oriental fruit flyand guava fruit fly(Diptera: Tephritidae) on six host fruits. Acta Ecologica Sinica, 2014, 34(9): 2274-2281. (in Chinese)
[18] 黄爱玲, 王小云, 陆温, 郑霞林. 橘小实蝇对不同寄主果实的产卵偏好及其与寄主果实理化性质的相关性. 植物保护学报, 2022, 49(2): 595-602.
HUANG A L, WANG X Y, LU W, ZHENG X L. Ovipositional preference of oriental fruit flyto different host fruits and its correlation with physical and chemical properties of host fruits. Journal of Plant Protection, 2022, 49(2): 595-602. (in Chinese)
[19] 李貌, 方薛交, 张金龙, 肖春, 闫振华, 刘娜, 陈国华, 张晓明. 桔小实蝇对5个石榴品种的产卵选择适应性. 南方农业学报, 2021, 52(12): 3408-3414.
LI M, FANG X J, ZHANG J L, XIAO C, YAN Z H, LIU N, CHEN G H, ZHANG X M. Oviposition preference of(Hendel) to five varieties of. Journal of Southern Agriculture, 2021, 52(12): 3408-3414. (in Chinese)
[20] 郭腾达, 宫庆涛, 孙瑞红, 武海斌, 姜莉莉. 橘小实蝇对桃果实的产卵选择. 中国农学通报, 2022, 38(4): 92-98.
GUO T D, GONG Q T, SUN R H, WU H B, JIANG L L. Oviposition preference ofon peach. Chinese Agricultural Science Bulletin, 2022, 38(4): 92-98. (in Chinese)
[21] 黄慧欣, 李媛, 黄爱玲, 王小云, 郑霞林, 陆温. 橘小实蝇对5个品种火龙果果实的产卵选择. 果树学报, 2021, 38(3): 394-402.
HUANG H X, LI Y, HUANG A L, WANG X Y, ZHENG X L, LU W. Oviposition preference of(Hendel) to five varieties of pitaya. Journal of Fruit Science, 2021, 38(3): 394-402. (in Chinese)
[22] 袁彬乔, 赵向杰, 张金勇, 黄天祥, 涂洪涛. 落叶果树对橘小实蝇生长发育和繁殖的影响. 果树学报, 2022, 39(12): 2397-2405.
YUAN B Q, ZHAO X J, ZHANG J Y, HUANG T X, TU H T. Effects of deciduous fruit species on the growth, development and reproduction of. Journal of Fruit Science, 2022, 39(12): 2397-2405. (in Chinese)
[23] 周宇翔, 商晗武, 蔡冲, 陈玲. 桔小实蝇在不同杨梅品种上的生长发育特性研究. 植物检疫, 2011, 25(6): 21-24.
ZHOU Y X, SHANG H W, CAI C, CHEN L. Biology of the oriental fruit fly () over three varieties of red bayberry () cultivars. Plant Quarantine, 2011, 25(6): 21-24. (in Chinese)
[24] 梁家燕, 贺海雄, 罗全丽, 何忠雪, 潘晓莲, 蒙仕鹏, 朱雄. 贵州山区苹果园橘小实蝇发生为害规律及绿色防控技术. 中国农技推广, 2018, 34(8): 59-60.
LIANG J Y, HE H X, LUO Q L, HE Z X, PAN X L, MENG S P, ZHU X. Occurrence and damage law ofin apple orchards of Guizhou mountainous areas and its green prevention and control technology. China Agricultural Technology Extension, 2018, 34(8): 59-60. (in Chinese)
[25] 朱学松, 段钟情, 陈平, 潘嫦艳, 封谷祥, 刘凤娥, 李燕. 曲靖市桔小实蝇为害情况及发生规律. 中国南方果树, 2022, 51(6): 184-187.
ZHU X S, DUAN Z Q, CHEN P, PAN C Y, FENG G X, LIU F E, LI Y. The damage and occurrence regularity ofin Qujing City. South China Fruits, 2022, 51(6): 184-187. (in Chinese)
[26] 宋来庆, 刘美英, 赵玲玲, 唐岩, 孙燕霞, 刘学卿, 张学勇, 姜中武. 桔小实蝇在烟台果树产区的监测与防控. 烟台果树, 2019(2): 36-37.
SONG L Q, LIU M Y, ZHAO L L, TANG Y, SUN Y X, LIU X Q, ZHANG X Y, JIANG Z W. Monitoring and prevention and control ofin Yantai fruit tree production area. Yantai fruits, 2019(2): 36-37. (in Chinese)
[27] 陈学森, 韩明玉, 苏桂林, 刘凤之, 过国南, 姜远茂, 毛志泉, 彭福田, 束怀瑞. 当今世界苹果产业发展趋势及我国苹果产业优质高效发展意见. 果树学报, 2010, 27(4): 598-604.
CHEN X S, HAN M Y, SU G L, LIU F Z, GUO G N, JIANG Y M, MAO Z Q, PENG F T, SHU H R. Discussion on today’s world apple industry trends and the suggestions on sustainable and efficient development of apple industry in China. Journal of Fruit Science, 2010, 27(4): 598-604. (in Chinese)
[28] 李云国, 蔡兆翔, 马静, 王顺富, 邓芳, 马钧, 孔宝华. 昭通高原苹果产业发展现状及可持续发展对策研究. 云南农业大学学报(社会科学), 2015, 9(5): 69-73.
LI Y G, CAI Z X, MA J, WANG S F, DENG F, MA J, KONG B H. Study on the current situation of the development and countermeasures for sustainable development of apple industry in Zhaotong. Journal of Yunnan Agricultural University (Social Science), 2015, 9(5): 69-73. (in Chinese)
[29] 陈瑶瑶, 古枫, 钟国华, 易欣.在桔小实蝇求偶和交配行为中的作用. 昆虫学报, 2020, 63(8): 924-931.
CHEN Y Y, GU F, ZHONG G H, YI X. Role ofin courtship and mating behaviors in(Diptera: Tephritidae). Acta Entomologica Sinica, 2020, 63(8): 924-931. (in Chinese)
[30] 彭帅, 郑丽霞, 吴伟坚. 瓜实蝇对寄主植物的产卵选择性. 环境昆虫学报, 2013, 35(2): 273-276.
PENG S, ZHENG L X, WU W J. Host plant oviposition preference of(Coquillett) (Diptera: Tephritidae). Journal of Environmental Entomology, 2013, 35(2): 273-276. (in Chinese)
[31] 李慧敏, 郑洁新, 曾凯芳, 邓丽莉. 不同采收成熟度对蜜奈夏橙果实营养品质的影响. 食品工业科技, doi: 10.13386/j.issn1002-0306. 2023020014.
LI H M, ZHENG J X, ZENG K F, DENG L L. Effect of different harvest maturity on the nutritional quality of ‘Midknight’ valencia orange fruit. Science and Technology of Food Industry, doi: 10.13386/ j.issn1002-0306.2023020014. (in Chinese)
[32] 李貌, 张金龙, 闫振华, 刘娜, 黄禹禹, 陈国华, 张晓明. 不同寄主对瓜实蝇和南亚实蝇生长发育和产卵选择的影响. 植物保护, 2022, 48(3): 151-158.
LI M, ZHANG J L, YAN Z H, LIU N, HUANG Y Y, CHEN G H, ZHANG X M. Effects of different hosts on growth, development and oviposition selection ofand. Plant Protection, 2022, 48(3): 151-158. (in Chinese)
[33] Jaleel W, Yin J, Wang D, HE Y, LU L, SHI H. Using two-sex life tables to determine fitness parameters of fourspecies (Diptera: Tephritidae) reared on a semi-artificial diet. Bulletin of Entomological Research, 2018, 108(6): 707-714.
[34] 周鹏. 桔小实蝇生殖行为研究[D]. 贵阳: 贵州大学, 2016.
ZHOU P. Reproductive behavior of(Hendel)[D]. Guiyang: Guizhou University, 2016. (in Chinese)
[35] 胡昌雄, 范苇, 张倩, 陈国华, 殷红慧, 徐天养, 杨进波, 杨航, 吴道慧, 张晓明. 基于两性生命表和年龄-阶段捕食率的南方小花蝽对西花蓟马的控制作用. 中国农业科学, 2021, 54(13): 2769-2780. doi: 10.3864/j.issn.0578-1752.2021.13.007.
HU C X, FAN W, ZHANG Q, CHEN G H, YIN H H, XU T Y, YANG J B, YANG H, WU D H, ZHANG X M. Control effect ofonbased on the two-sex life table and the age-stage-specific predation rate. Scientia Agricultura Sinica, 2021, 54(13): 2769-2780. doi: 10.3864/j.issn.0578-1752.2021.13.007. (in Chinese)
[36] 项前, 杨卫平, 于金鑫, 瞿佳宝, 曾鑫年, 吴红霞, 刘家莉. 桔小实蝇对石榴的产卵偏好性研究. 环境昆虫学报, 2022, 44(5): 1308-1318.
XIANG Q, YANG W P, YU J X, QU J B, ZENG X N, WU H X, LIU J L. Oviposition preference of the oriental fruit flyon pomegranate. Journal of Environmental Entomology, 2022, 44(5): 1308-1318. (in Chinese)
[37] Jallow M, Zalucki M. Within- and between-population variation in host-plant preference and specificity in Australian(Hubner) (Lepidoptera: Noctuidae). Australian Journal of Zoology, 1996, 44(5): 503-519.
[38] 董子舒, 张玉静, 段云博, 郑霞林, 陆温. 植食性昆虫产卵寄主选择影响因素及机制的研究进展. 南方农业学报, 2017, 48(5): 837-843.
DONG Z S, ZHANG Y J, DUAN Y B, ZHENG X L, LU W. Influencing factors and selection mechanisms of phytophagous insects for oviposition host plants. Journal of Southern Agriculture, 2017, 48(5): 837-843. (in Chinese)
[39] 蔡子坚, 胡菡青, 韦晓霞, 陈瑾, 林雄杰, 范国成, 吴如健. 杨桃果实性状与橘小实蝇危害严重度的关系. 福建农业学报, 2012, 27(12): 1298-1302.
CAI Z J, HU H Q, WEI X X, CHEN J, LIN X J, FAN G C, WU R J. Traits and damage of carambola fruit by. Fujian Journal of Agricultural Sciences, 2012, 27(12): 1298-1302. (in Chinese)
[40] 李媛, 黄爱玲, 黄慧欣, 王小云, 郑霞林, 陆温. 橘小实蝇对三华李果实的产卵选择. 南方农业学报, 2020, 51(2): 319-326.
LI Y, HUANG A L, HUANG H X, WANG X Y, ZHENG X L, LU W. Oviposition selectivity of(Hendel) to. Journal of Southern Agriculture, 2020, 51(2): 319-326. (in Chinese)
[41] 金菊, 阮赞誉, 黄珍富, 赖贵炎, 黄颂颂, 范骁凌. 莲雾果实挥发物对橘小实蝇的引诱作用. 华南农业大学学报, 2015, 36(3): 71-77.
JIN J, RUAN Z Y, HUANG Z F, LAI G Y, HUANG S S, FAN X L. Attractions of volatiles from wax-apple fruit to the oriental fruit fly. Journal of South China Agricultural University, 2015, 36(3): 71-77. (in Chinese)
[42] KAMALA JAYANTHI P d, Woodcock C M, Caulfield J, CAULFIELD J, BIRKETT M A, BRUCE T J A. Isolation and identification of host cues from mango,, that attract gravid female oriental fruit fly,. Journal of Chemical Ecology, 2012, 38(4): 361-369.
[43] 张璇, 毕亚銮, 方琦, 汪芳, 叶恭银. 两种果蝇对不同成熟期草莓果实及其挥发物的趋性行为分析. 中国生物防治学报, doi: 10.16409/j.cnki.2095-039x.2022.11.016.
ZHANG X, BI Y L, FANG Q, WANG F, YE G Y. Tropism behavior analysis of twospecies to fruits and volatiles of strawberry at different mature stages. Chinese Journal of Biological Control, doi: 10.16409/j.cnki.2095-039x.2022.11.016. (in Chinese)
[44] 王冰, 李慧敏, 操海群, 王桂荣. 挥发性化合物介导的植物-植食性昆虫-天敌三级营养级互作机制及应用. 中国农业科学, 2021, 54(8): 1653-1672. doi: 10.3864/j.issn.0578-1752.2021.08.007.
WANG B, LI H M, CAO H Q, WANG G R. Mechanisms and applications of plant-herbivore-natural enemy tritrophic interactions mediated by volatile organic compounds. Scientia Agricultura Sinica, 2021, 54(8): 1653-1672. doi: 10.3864/j.issn.0578-1752.2021.08.007. (in Chinese)
[45] UNAHAWUTTI U, INTARAKUMHENG R, OONTHONGLANG P, PHANKUM S, FOLLETT P A. Nonhost status of mangosteen toand(Diptera: Tephritidae) in Thailand. Journal of Economic Entomology, 2014, 107(4): 1355-1361.
[46] 郭腾达, 姜莉莉, 孙瑞红, 宫庆涛, 叶保华. 橘小实蝇危害3种苹果的风险评估. 果树学报, 2021, 38(2): 231-241.
GUO T D, JIANG L L, SUN R H, GONG Q T, YE B H. Risk evaluation of three apple cultivars affected byJournal of Fruit Science, 2021, 38(2): 231-241. (in Chinese)
[47] 王凯, 程禹铭, 孙清明, 杨益众, 苏宏华. 甜菜夜蛾在不同寄主植物上的适合度及寄主选择. 中国生物防治学报, 2023, 39(2): 346-354.
WANG K, CHENG Y M, SUN Q M, YANG Y Z, SU H H. Effect of different host plants on the growth and development of(Hübner) and its host selection. Chinese Journal of Biological Control, 2023, 39(2): 346-354. (in Chinese)
[48] 张智英, 赵波, 张亮, 梁华娟. 番石榴实蝇寄主选择性试验. 应用昆虫学报, 2011, 48(2): 359-363.
ZHANG Z Y, ZHAO B, ZHANG L, LIANG H J. The host preference experiment of.Chinese Journal of Applied Entomology, 2011, 48(2): 359-363. (in Chinese)
[49] 陈萍, 刘欢, 侯茂林. 寄主转换对稻纵卷叶螟寄主种群适合度及生命表参数的影响. 昆虫学报, 2021, 64(3): 400-408.
CHEN P, LIU H, HOU M L. Effects of host switch on the performance and life-table parameters of host populations of(Lepidoptera: Pyralidae).Acta Entomologica Sinica, 2021, 64(3): 400-408. (in Chinese)
[50] Mayhew P J. Adaptive patterns of host-plant selection by phytophagous insects. OIKOS, 1997, 79(3): 417-418.
[51] Sicsú P R, Macedo R H, Sujii E R. Lady beetle oviposition site choices: Maternal effects on offspring performance. Florida Entomologist, 2020, 103(2): 228.
[52] 张灿, 刘慧, 梁帆, 胡学难, 赵菊鹏, 马骏. 木瓜实蝇对七种寄主果实的选择性. 环境昆虫学报, 2012, 34(3): 283-288.
ZHANG C, LIU H, LIANG F, HU X N, ZHAO J P, MA J. Oviposition selection and larval suitability ofDrew & Hancock (Diptera: Tephritidae) on seven kind of fruits.Journal of Environmental Entomology, 2012, 34(3): 283-288. (in Chinese)
[53] 何超, 孔琼, 袁盛勇, 沈登荣, 谢昆, 王传铭. 南瓜实蝇与瓜实蝇成虫在4种寄主上的产卵竞争. 江西农业大学学报, 2019, 41(3): 484-490.
HE C, KONG Q, YUAN S Y, SHEN D R, XIE K, WANG C M. Interspecific competition betweenandon four host plants. Acta Agriculturae Universitatis Jiangxiensis, 2019, 41(3): 484-490. (in Chinese)
[54] Gripenberg S, Mayhew P J, Parnell M, Roslin T. A meta-analysis of preference-performance relationships in phytophagous insects. Ecology Letters, 2010, 13(3): 383-393.
[55] Liu Z D, Scheirs J, Heckel D G. Trade-offs of host use between generalist and specialistsibling species: adult oviposition and larval performance. Oecologia, 2012, 168(2): 459-469.
selection preference and adaptability oftodifferent varieties of
LI XingXing, ZHOU GuoFu, LUO GuanYu, CHEN SiRong, ZHANG JinLong, CHEN GuoHua*, ZHANG XiaoMing*
College of Plant Protection, Yunnan Agricultural University/National Key Laboratory for Conservation and Utilization of Bio-resources in Yunnan, Kunming 650201
【Objective】This study aims to investigate the preference offor fruits of six mainvarieties and to understand the effects of different varieties ofon the growth and development of.【Method】Three early maturing varieties (Huashuo, Honglu, Gala) and three late maturing varieties (Duanzhifushi, Yanfu No. 3, Qincui) were used as test materials. The hardness and sugar content offruit were determined by hardness tester and a brix refractometer. Three types of fruit handling methods were employed: whole fruit, peeled fruit and fruit odor. Fruits of the same ripening batch of the same handling method were placed in rearing cages, to measure the numbers of visits and laid eggs byon fruits of different varieties were determined. Feeding different varieties of fruits to the first hatching larvae, the growth and development parameters of larval duration, pupation rate, pupal weight and other parameters after feeding were determined. The selection preference and development adaptability ofto different varieties ofwere analyzed.【Result】There were differences in the numbers of visits and laid eggs ofon different varieties of fruits. The female adults were more sensitive to different varietiesandfruit handling methods.preferred to choose Gala among whole fruits of the early maturing varieties, which had 9.0 female visits and 105.0 eggs per fruit, respectively. While among the late maturing varieties,showed a stronger preference for Qincui, and the numbers of female visits and laid eggs on this variety reached to 5.5 and 64.2 eggs per fruit, respectively. The fruit handling methods influenced the selection preference of. With the exception of Gala, the numbers of visits and laid eggs on peeled fruits were higher in different fruit handling methods of the same variety. Among the early maturing varieties, Huashuo had held the highest number of laid eggs on the peeled fruit, reached to 201.6 eggs per fruit, and among the late maturing varieties, Yanfu No. 3 had held the highest number of laid eggs on fruit odor, reached to 83.8 eggs per device. Two-way ANOVA found that variety and fruit handling method had an impact on the number of visits and laying eggs. Larvae ofhad held the shortest period of 11.9 d in Gala and the shortest pupal stage in Huashuo of 9.9 d after feeding on different fruits. Pupation and emergence rates were higher in late maturing varieties. The difference in the age-stage-specific survival rate of the larvae that fed on different varieties was mainly in the 2nd instar stage, and the 2nd instar larvae which fed on Huashuo and Gala had held fastest development rate and peaked at the 3rd day.【Conclusion】There were differences in the selection preference ofon differentvarieties. Gala was preferred in early maturing varieties, while Qincui was preferred in late maturing varieties. Fruit integrity and fruit odor had an effect on the selection preference. The numbers of visits and laid eggs on peeled fruit and fruit odor were generally higher than those on whole fruit.can finish the growth and development of larvae onfruit, and it has the ability to reproduce damage in apple orchards. The correlation between the preference offorfruit and the fitness of offspring’s growth and development is not strong, which is different from the hypothesis of “selectivity and fitness”. In the comprehensive management ofin apple orchards, the damage monitoring of Gala and Qincui varieties should be strengthened, and the damaged fruits should be avoided and removed in time in agricultural operations.
; apple (); growth and development; selection preference;selective adaptability
10.3864/j.issn.0578-1752.2023.17.010
2023-05-17;
2023-06-23
云南省重大科技专项(202102AE090006)、云南省中青年学术技术带头人后备人才项目(202105AC160071)、“云南省高层次人才培养支持计划”青年拔尖人才专项(YNWRNBJ2020291)、“云南省昭阳区苹果产业科技特派团”项目(202104BI090028)
李星星,E-mail:lixxstarr@163.com。通信作者陈国华,E-mail:chenghkm@126.com。通信作者张晓明,E-mail:zxmalex@126.com
(责任编辑 岳梅)
猜你喜欢
今日农业(2022年13期)2022-09-15
农产品加工(2017年11期)2017-07-31
妇女生活(2017年4期)2017-04-08
食品科学(2017年5期)2017-03-31
中国糖料(2016年1期)2016-12-01
时代农机(2016年6期)2016-12-01
蔬菜(2016年8期)2016-10-10
湖南农业(2016年3期)2016-06-05
湖南农业(2016年3期)2016-06-05
分析化学(2015年6期)2015-06-18
