
重金属胁迫下花生根瘤菌的溶磷作用研究
2023-09-26 09:12:48崔永亮雷久城余秀梅程祖强吴航玉张玉婷陈绪玲
资源开发与市场 2023年9期
崔永亮,罗 伟,雷久城,余秀梅,程祖强,吴航玉,张玉婷,陈绪玲,胡 骥
(1.四川省自然资源科学研究院,四川 成都 610041;2.四川省生态环境科学研究院,四川 成都 610041;3.四川农业大学资源学院,四川 成都 610041;4.四川省自然资源科学研究院成都分院,四川 成都 610000)
0 引言
近年来,农田土壤重金属污染问题日渐凸显,重金属对土壤的组成及其功能产生深远的消极影响[1],造成了食品安全问题和生态系统破坏等不良后果[2]。为修复土壤重金属污染,改善土壤生态环境,科学家深入研究了多种土壤重金属修复技术,如物理修复技术、化学修复技术和生物修复技术。然而,传统的化学修复和物理修复技术成本高、效率低,且存在产生二次污染的风险[3]。目前,经济高效、绿色环保的生物修复技术,尤其是微生物—植物联合修复技术,已成为解决土壤重金属污染问题的重要手段。微生物-植物联合修复是一种利用植物根际微生物分泌的活性物质来提高植物对重金属的吸收、累积或固定效率的修复技术。根际微生物可以通过分泌植物激素、维生素,利用根瘤菌固氮等方式促进植物生长,以提高植物生物量的方式增加重金属的总积累量[4]。根瘤菌—豆科植物共生体系是目前修复土壤重金属污染较为有效的方法之一。在土壤重金属浓度过高时,根瘤菌—豆科植物共生体系能形成解毒机制和重金属抗性来抵抗重金属胁迫[5-7],同时固氮作用还可以大幅提高土壤养分促进植物生长[8,9],显著提高作物产量[10],其所固定的氮约占生物固氮总量的65%[11]。邹萌萌的研究表明,白三叶—根瘤菌修复体系在被Cu 污染的土壤中能通过提高白三叶的生物量及其对Cu 的吸收量来提升土壤修复效率[12]。宋修勇的研究发现,接种了根瘤菌的天蓝苜蓿对重金属胁迫的耐受性更强,且结瘤数与固氮酶活均有显著提高[13]。练建军等在使用紫花苜蓿—根瘤菌体系修复钼污染土壤时发现,根瘤菌不仅能促进重金属钼由水溶态向中间过渡态的转变,也能增大土壤残渣态钼的比例,从而促进紫花苜蓿对钼的吸收,同时降低钼对紫花苜蓿的生物毒性,修复效果良好[14]。
在筛选适合开展重金属土壤污染修复的生物材料时,不仅需要考虑植物的重金属富集能力,还需要考虑植物的经济效益。因此,作为油料作物的花生受到了广泛关注。研究表明,花生对土壤中的Cd、Zn、Cu、Cr、Hg、Ni 等重金属具有良好的吸收作用[15-18],且花生果实对Cd的富集能力最强[16]。然而,鲜有研究探讨添加花生根瘤菌是否能提高花生对土壤重金属的耐受能力和富集能力,筛选具有重金属抗性的花生根瘤菌的研究也同样稀缺。
此外,最新研究表明,根瘤菌除固氮外,还具有溶磷、解钾、分泌IAA(indoleacetic acid,吲哚乙酸)等促生功能[19,20]。磷元素是植物生长的必需元素,但土壤中的磷元素都是以难溶于水的化合物的形式存在,难以被作物直接吸收。植物根系和土壤微生物的水解作用能将其转化为植物可利用的形式,但转化过程缓慢,效率低下[21,22]。近年来的研究表明,根瘤菌与共生植物形成的根瘤能提高磷元素的转化效率,显著提高植物生物量[23-26]。
综上,本研究拟对从典型镉污染区域农田中采集的花生根瘤菌进行研究,筛选出在重金属胁迫下具有较好溶磷能力的菌株,为微生物—植物联合修复技术提供重要的根瘤菌资源。
1 材料与方法
1.1 材料
花生根瘤菌。从镉污染区域农田采集的花生根瘤中分离纯化得到36 株花生根瘤菌,置于60%甘油中保存于-80℃冰箱。
花生根瘤中存在多种细菌,可通过使用YMA培养基对根瘤菌进行分离提纯,其配置方法为酵母粉1.5g/L,甘露醇1g/L、K2HPO40.5g/L、MgSO4·7H2O 0.2g/L、NaCl 0.1g/L、去离子水1 L,pH 7.0。PKO 固体培养基常用于测定微生物是否具有分解无机磷的能力,筛选解磷微生物,其配置方法为葡萄糖10g/L、Ca3(PO4)25.0g/L、(NH4)2SO40.5g/L、NaCl 0.2g/L、MgSO40.03g/L、MnSO40.03g/L、FeSO40.003g/L、KCl 0.2g/L、琼脂20g/L、去离子水1L,pH7.2。PKO 液体培养基常用于对解磷微生物分解无机磷的能力进行定量分析,精确比较不同解磷微生物分解无机磷的能力,其配置方法为葡萄糖10g/L、Ca3(PO4)25.0g/L、(NH4)2SO40.5g/L、NaCl 0.2g/L、MgSO40.03g/L、MnSO40.03g/L、FeSO40.003g/L、KCl 0.2g/L、去离子水1L,pH7.2。
1.2 方法和设备
1.2.1 方法
1.2.2 设备
恒温培养箱,恒温震荡摇床,离心机,分光光度计,高压灭菌锅,超净工作台,ICP-OES等。
1.2.3 菌株活化
在无菌条件下,用接种环蘸取保种于60%甘油中的花生根瘤菌分别划线于YMA 培养基上,28℃培养2d。
1.2.4 菌株溶磷作用测定
根瘤菌溶磷能力测定实验分为定性测定和定量测定[22]。
①定性测定。将根瘤菌接种在PKO 固体培养基上,每个平板采用点接法接3 个点,每点接4μL培养好的菌液,28 ℃下培养7d 后测量菌落直径和周围透明圈直径,并计算有效透明圈直径。②定量测定。称取1.5g 左旋抗坏血酸溶于100mL钼锑混合液[27]。在试管中装入6mL 液体PKO 培养基,灭菌后备用。按1%接种量在液体PKO培养基中接种被测菌株,每株菌设置3 个重复,并以接种无菌水的培养基为对照。在150r/min,28 ℃的环境下振荡培养7d 后,取培养液在8000r/min 的转速下离心10min,取上清液稀释后,用钼锑抗比色法测定上清液中的磷含量。取3mL 上清液,测定pH 值。菌液中磷的含量=(P×V1×TS)/V2(P:根据标曲计算出的磷的浓度;V1:测定OD值所定容的体积;TS:分取倍数;V2:所摇菌液的体积)。
公式(2)中Qd+Qg-Qw为干燥垃圾所需热量,设每千克湿垃圾含水率取50%,每千克湿垃圾需蒸发水分m=0.5 kg,垃圾属于混合物,且具有各向不同性,其比热容较难求得,且垃圾比热容小于水。设干燥1kg垃圾需要热量为q,按公式(3)计算:
1.2.5 根瘤菌的分子鉴定与系统进化分析
采用异硫氰酸胍(GUTC)法提取根瘤菌的DNA进行测序鉴定[22],并用核酸检测仪与琼脂糖凝胶电泳对DNA 的质量进行检测分析。对提取的DNA使用PCR 引物27F:5′-GAGTTTGATCACTGGCTCAG-3′ 和 1492R:TACGGCTACCTTGTTACGACTT-3′进行16S rRNA扩增,得到的扩增产物用1%琼脂糖凝胶电泳检测其质量并进行测序,将测序结果在GenBank 中进行分析,用邻接法在MEGA6.0 中构建16S rRNA系统发育树。
1.2.6 根瘤菌重金属抗性测定
对筛选的菌株进行重金属抗逆性分析。配置YMA液体培养基,在试管中加入5ml 培养基后,分别加入不同浓度梯度的Cu、Ni、Cd、Cr、Zn 五种重金属,浓度梯度按照0、5、10、15、…、70、75、80mg/kg设置。将菌株以1:100 的比例接种到培养基中,以不接种菌株但添加重金属的培养基作为对照组,每组处理设置3 个重复。28℃、150r/min 培养2 d 后用分光光度计分别测定OD600,获得重金属对菌株的致死浓度。
1.2.7 重金属胁迫下花生根瘤菌的溶磷能力测定
根据典型镉污染区域农田重金属抗性测定结果,以5 种重金属的致死浓度为上限,从0mg/kg 开始,分别加入到灭菌后的PKO 液体培养基中,再接入根瘤菌悬液3mL,以不接菌的PKO液体培养基作为对照组,每个处理设置3 个重复。28 ℃的环境下振荡培养7 d后,测定培养液中可溶性磷含量和pH值,分析不同浓度重金属胁迫下根瘤菌溶磷能力及对应pH值的变化。
2 结果及分析
2.1 根瘤菌溶磷能力测定
经过定性测量无机磷固体培养基上出现的透明圈来判断菌株是否拥有溶磷能力。在36 株根瘤菌中,有19 株根瘤菌产生了大小不一的溶磷圈,透明圈直径为0.07—1.47cm(表1)。在这19 株菌株中,只有1 株菌株(HM28)的透明圈直径大于1 cm,有3 株菌株(HW22、HG13、HG16)的透明圈直径在0.5—1cm之间。

表1 花生根瘤菌溶磷能力测定Table 1 Phosphorus solubilization of peanut rhizobium detection
使用钼锑抗比色法进一步定量测定初筛菌株的溶磷能力。通过定性测定筛选出的19 株根瘤菌其溶磷量各不相同,溶磷量为7.54-55.81μg/mL(表1)。有2 株菌株(HM28、HM2)的溶磷量超过50μg/mL。有4 株菌株(HM21、HW42、HW39、HW38)的溶磷量在40-50μg/mL 之间。有2 株菌株(HG8、HM52)的溶 磷 量在30-40μg/mL 之间。菌 株HM28、HM2 和HM21 具有较高由效溶磷能力,分别为55.81μg/mL、52.82μg/mL和48.61μg/mL。
2.2 根瘤菌的分子鉴定
使用测序结果构建系统发育树(图1)。

图1 高效溶磷花生根瘤菌16S rRNA基因的系统发育树Figure 1 Phylogenetic tree of 16S rRNA gene of effective phosphorus-soluble peanut rhizobium
由图1 可知,溶磷能力较好的3 株根瘤菌均属于根瘤菌属(Rhizobium),但这3 种根瘤菌在系统发育树上处于两个分支。其中,HM2 和HM21 与标准菌株Rhizobium altiplani BR 10423T 在同一个分支上,因此将两种菌株初步鉴定为Rhizobium altiplani;HM28 与标准菌株Rhizobium metallidurans ChimEc 512T在同一个分支上,因此将其初步鉴定为Rhizobium metallidurans。
2.3 根瘤菌重金属抗性测定
具有较好溶磷作用的根瘤菌的重金属(Cu、Ni、Cd、Cr、Zn)抗性测定结果如图1 所示。HM28 对Cu、Ni、Cd、Cr、Zn 的 致 死 浓 度 分 别 为30mg/kg、40mg/kg、10mg/kg、25mg/kg、50mg/kg,因 此 选 取0—30mg/kg、0—40mg/kg、0—10mg/kg、0—25mg/kg、0—50mg/kg作为浓度区间分别进行Cu、Ni、Cd、Cr、Zn胁迫下的溶磷作用测定。HM2 对Cu、Ni、Cd、Cr、Zn的致死浓度分别为50mg/kg、30mg/kg、10mg/kg、25mg/kg、20mg/kg,因此选取0—50mg/kg、0—30mg/kg、0—10mg/kg、0—25mg/kg、0—20mg/kg 作为浓度区间分别进行Cu、Ni、Cd、Cr、Zn 胁迫下的溶磷作用测定。HM21 对Cu、Ni、Cd、Cr、Zn 的致死浓度分 别 为50mg/kg、50mg/kg、50mg/kg、50mg/kg、50mg/kg,因此选取0—50mg/kg、0—50mg/kg、0—50mg/kg、0—50mg/kg、0—50mg/kg 作为浓度区间分别进行Cu、Ni、Cd、Cr、Zn胁迫下溶磷作用测定。
2.4 重金属胁迫下花生根瘤菌的溶磷能力测定
钼锑抗比色法测定溶磷量的实验结果显示,3株根瘤菌在5 种重金属的胁迫下,随着重金属浓度逐渐增加,活菌数降低,pH值逐渐升高,其溶磷作用均降低(图2—4)。

图2 不同重金属胁迫下HM28 的溶磷能力及pH值变化曲线Figure 2 Phosphorus solubilization and pH change curves of HM28 under different heavy metal stress
由图2 可知,菌株HM28 在Cu 的胁迫下,菌株在0—12mg/kg 的浓度之间溶磷量下降较快,在12—18mg/kg 的 浓度 之 间 几 乎 无 变化,在18—30mg/kg的浓度之间下降较缓慢,在浓度为30mg/kg时,没有溶磷作用;在Ni的胁迫下,菌株在0—8mg/kg和16—24mg/kg的浓度之间溶磷量变化比较小,在8—16mg/kg 和24—40mg/kg 的浓度之间下降较快,在浓度为40mg/kg 时,没有溶磷作用;在Cd 的胁迫下,菌株在0—4mg/kg 的浓度之间溶磷量下降较缓慢,在4—6mg/kg 浓度之间几乎没有变化,在6—10mg/kg的浓度之间下降较快,在浓度为10mg/kg时,没有溶磷作用;在Cr 的胁迫下,菌株在0—10mg/kg 的浓度之间溶磷量下降较快,在10—25mg/kg的浓度之间下降较缓慢,在浓度为25mg/kg时,失去溶磷作用;在Zn的胁迫下,菌株在0—20 mg/kg 的浓度之间溶磷量下降较快,在20—50mg/kg的浓度之间溶磷量下降较缓慢,在50mg/kg浓度时,失去溶磷作用。
从图3 可知,菌株HM2 在Cu的胁迫下,菌株在0—20mg/kg 的浓度之间溶磷量下降较快,在20—50mg/kg浓度之间下降较缓慢;在Ni 的胁迫下,菌株在0—6mg/kg的浓度之间溶磷量几乎无变化,在6—30mg/kg的浓度之间下降较快,在浓度为30mg/kg时,失去溶磷作用;在Cd 的胁迫下,菌株在0—8mg/kg 的浓度之间溶磷量下降较快,在浓度为10mg/kg时,失去溶磷作用;在Cr的胁迫下,菌株在0—5mg/kg 的浓度之间溶磷量下降较快,在5—25mg/kg之间下降较缓慢,在浓度为25mg/kg时,失去溶磷作用;在Zn 的胁迫下,菌株在0—12mg/kg的浓度之间溶磷量下降较快,在12—20mg/kg 之间变化不大,在浓度为20mg/kg时,失去溶磷作用。

图3 不同重金属胁迫下HM2 的溶磷能力及pH值变化曲线Figure 3 Phosphorus solubilization and pH change curves of HM2 under different heavy metal stress
从图4 可知,菌株HM21 在Cu 的胁迫下,菌株在0—50mg/kg的浓度之间溶磷量都下降较快,在浓度为50mg/kg时,失去溶磷作用;在Ni的胁迫下,菌株在0—10mg/kg 和40—50mg/kg 的浓度之间溶磷量变化较快,在10—40mg/kg 的浓度之间变化较小,在浓度为50mg/kg 时,失去溶磷作用;在Cd 的胁迫下,菌株在0—10mg/kg 的浓度之间溶磷量几乎无变化,在10—50mg/kg 浓度之间下降较快,在浓度为50mg/kg时,失去溶磷作用;在Cr的胁迫下,菌株在0—20mg/kg 的浓度之间溶磷量下降较快,在20—50mg/kg 之间下降缓慢,在浓度为50mg/kg时,失去溶磷作用;在Zn 的胁迫下,菌株在0—50mg/kg 的浓度之间溶磷量下降较快,在浓度为50mg/kg以上时,失去溶磷作用。
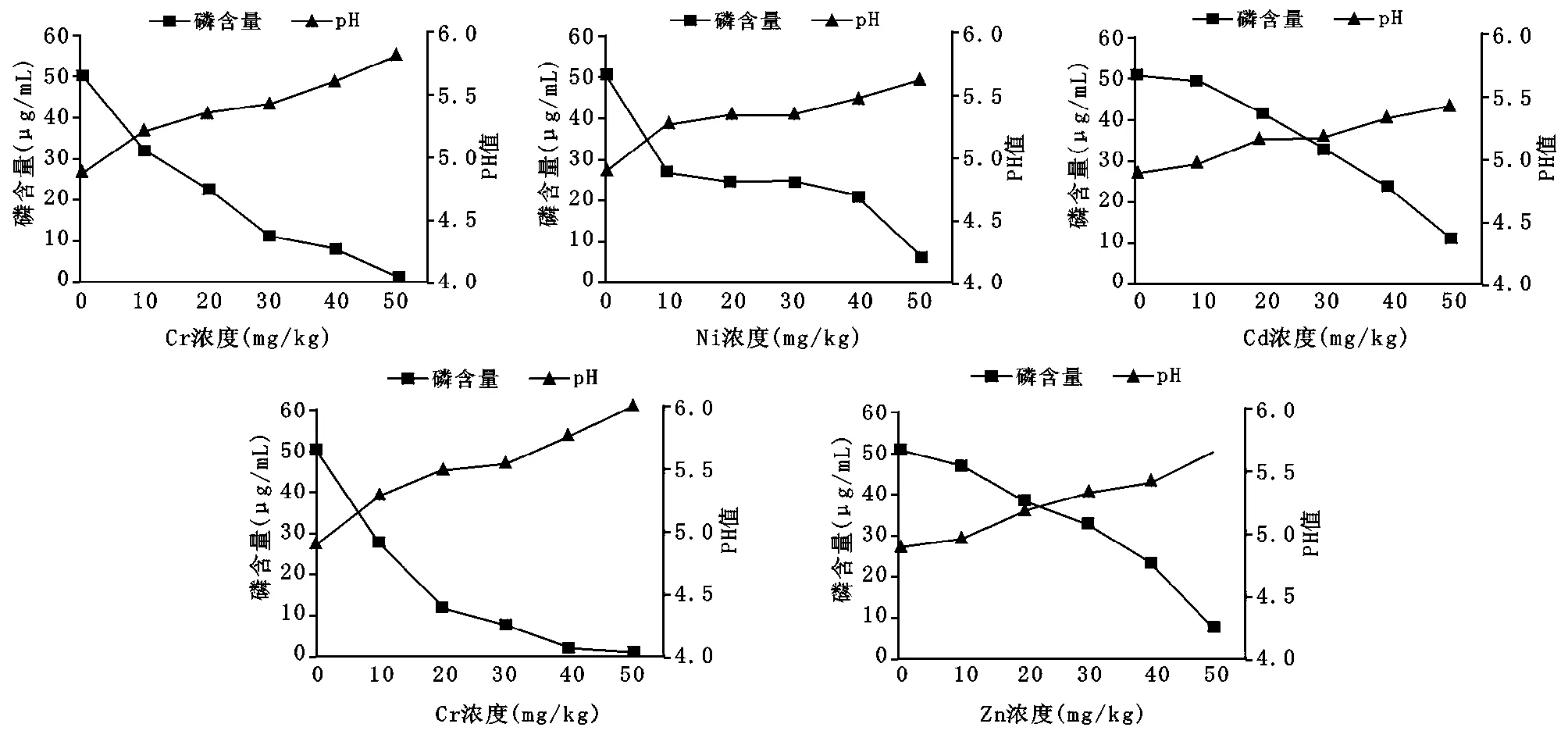
图4 不同重金属胁迫下HM21 的溶磷能力及pH值变化曲线Figure 4 Phosphorus solubilization and pH change curves of HM21 under different heavy metal stress
3 讨论
从镉污染区域的农田土壤中分离出的36 株根瘤菌,有19 株具有溶磷能力。也有部分研究者发现微生物的溶磷能力不仅受菌株自身特质的影响,也会受培养环境影响[23,26]。有相关研究表明,同一种菌株对不同的难溶性磷酸盐的溶解能力有所差异。李静等研究发现的产红青霉菌R3 对Ca3(PO4)2的溶解能力最高达328.79 mg/L,对AlPO4和FePO4的最高溶解量分别为95.99mg/L 和75.39mg/L,溶磷量Ca3(PO4)2>AlPO4>FePO4[28]。但本研究得到的仅为实验室特定培养条件下的测定结果,筛选出的菌株是否具有较好的田间溶磷能力需要进行进一步的深入研究。
在含有不同浓度梯度重金属的液体培养基中HM28、HM2、HM21 溶磷能力变化的测定结果表明,3 株根瘤菌的溶磷能力会随着重金属浓度的增高而降低,且根瘤菌的溶磷量和pH 值变化存在一定关系。研究表明,溶磷微生物通过分泌有机酸等物质分解难溶性矿物磷,导致培养液pH降低,但不同菌株分泌的有机酸种类和含量差异较大,所以不同菌株在培养时测得的pH值也不相同[25,29]。
严警等发现不同根瘤菌与不同豆科植物共生会因为两者基因的兼容性存在差异而影响促生能力[30]。一些根瘤菌可以与广泛的宿主共生,但一些根瘤菌只具有专一宿主。不同区域的环境,如气候、土壤等自然环境条件也会影响共生关系的建立。综上所述,根瘤菌与宿主建立共生关系是根瘤菌、植物和环境三者相互作用的结果。因此,已筛选的菌株与花生的共生作用是否能够实际应用于重金属污染土壤环境,还需要进行多品种、多地的应用研究,并阐明其互作机理。
4 结论
经过定性和定量测定,来自典型镉污染区域农田的36 株花生根瘤菌中,有52.8%的菌株具有溶磷能力,并且对不同重金属有不同程度的抗性,且随着重金属浓度增加,溶磷作用减弱,pH值逐渐升高,说明重金属对根瘤菌的溶磷作用有抑制作用。本研究获得了3 株具有较好溶磷能力的根瘤菌HM28,HM2 和HM21,其中HM28 和HM2 的溶磷量超过50μg/ml。此外,HM28 经初步鉴定为Rhizobium metallidurans,HM2 和HM21 经初步鉴定为Rhizobium altiplani。本研究筛选获得的3 株根瘤菌为重金属污染土壤的生物修复提供了可利用的菌种资源。
猜你喜欢
儿童时代·快乐苗苗(2022年10期)2022-12-09 08:53:44
园艺与种苗(2021年1期)2021-03-16 03:01:08
西藏农业科技(2019年3期)2019-11-04 00:35:08
甘肃农业大学学报(2019年2期)2019-06-01 01:41:42
山西农业大学学报(自然科学版)(2019年1期)2019-03-02 02:09:46
儿童时代·快乐苗苗(2016年4期)2016-11-07 01:09:26
广州化工(2016年8期)2016-09-02 00:48:01
河南农业科学(2016年5期)2016-07-24 16:58:11
学苑创造·A版(2016年7期)2016-07-06 18:11:05
儿童时代·快乐苗苗(2015年8期)2015-11-12 23:16:31
