
羊草LcZIP1 的铁转运功能鉴定
2023-09-25 03:42:54亢燕王耀辉牛天慧滕哲祁智杨佳
草业学报 2023年9期
亢燕,王耀辉,牛天慧,滕哲,祁智,杨佳
(内蒙古大学牧草与特色作物生物学教育部重点实验室,内蒙古 呼和浩特 010070)
羊草(Leymus chinensis)属于小麦族(Triticeae)赖草属(Leymus),是多年生禾草。羊草是我国重要的乡土植物,广泛分布在我国北方草原,具有较高的饲用价值、生态价值和经济价值。以羊草为主的草场是优质天然放牧场和割草场。羊草的研究始于1974 年,主要集中在种质资源、栽培和生物学特性等方面[1]。牧草矿质营养直接影响牧草和草原家畜的产量和质量,进而影响草原的生产力。然而关于羊草矿质营养吸收的分子调控机理研究较少。影响植物矿质营养吸收主要有两个因素,一是外在因素,即土壤的矿质元素含量。已有研究发现外源施加微肥即锌、铜、硼、钼等可以显著提高羊草干草的质量和产量,增产高达近50%[2];氮结合钾、硫、锌、铜可以促进羊草生长[3];二是内在因素,即植物体内的矿质元素转运蛋白和相关调控因子[4-5]。在羊草中已发现锌转运蛋白LcZNE1,具有锌结合和转运功能,参与响应高锌和低铁条件[4]。
锌铁转运蛋白(zinc-regulated transporter/iron-regulated transporter-like proteins,ZIP)在古菌、细菌、真菌、植物和哺乳动物中广泛存在,负责金属二价阳离子如Zn2+、Fe2+、Cu2+、Mn2+、Ni2+和Cd2+等的吸收和转运[5]。其中Fe、Zn、Cu、Mn 和Ni 是必需微量元素,其作为酶结构的一部分或辅因子介导植物的生长、发育和代谢[6]。Fe 具有氧化还原作用,在光合作用和呼吸作用的电子传递链中起着重要作用,参与氮同化、激素合成、活性氧(reactive oxygen species,ROS)清除以及生物胁迫响应等[6-7]。Zn 是多种酶的组成部分和辅因子,与维持生物膜的结构完整、蛋白质合成以及种子发育等相关[6]。预计有1200 多种蛋白质内包含Zn2+、结合Zn2+或运输Zn2+,这些蛋白包括锌指蛋白、转录因子、氧化还原酶和水解酶等。
目前,ZIP 家族在模式植物和农作物中受到越来越多的关注,如拟南芥(Arabidopsis thaliana)[8-9]、烟草(Nicotiana tabacum)[10]、蒺藜苜蓿(Medicago truncatula)[11]、野生二粒小麦(Triticum turgidumssp. dicoccoides)[12]、大麦(Hordeum vulgare)[13]、谷子(Setaria italica)[14]、水 稻(Oryza sativa)[15-16]、玉米(Zea mays)[17]、大豆(Glycine max)[18]、菜豆(Phaseolus vulgaris)[19]、葡萄(Vitis vinifera)[20]和脐橙(Citrus sinensis)[21]。目前Zn2+和Fe2+转运功能鉴定的主要手段是通过离子转运缺陷酵母突变体功能互补验证,常使用的突变体为Zn2+转运蛋白缺陷突变体(∆zrt1/∆zrt2、∆zrc1/∆cot1)和Fe2+吸收缺陷突变体(∆fet3/∆fet4)。在模式植物拟南芥中发现,AtZIP1、AtZIP2、AtZIP3、AtZIP7、AtZIP11 和AtZIP12 能够使酵母的Zn2+转运蛋白缺陷突变体(∆zrt1/∆zrt2)恢复Zn2+吸收功能,AtZIP7 也能使酵母的Fe2+吸收缺陷突变体(∆fet3/∆fet4)恢复Fe2+吸收功能。这些结果证明了AtZIP 的Zn2+或Fe2+转运功能[8-9]。此外还发现,水稻OsZIP1 能使酵母锌转运蛋白缺失突变体∆zrc1在高锌条件下正常生长,从而证明OsZIP1 具有锌转运功能。已有研究表明,OsZIP1在过量的锌、铜和镉的条件下被诱导表达,表达产物定位于内质网和质膜上,阻止水稻中过量的锌、铜和镉积累[16]。
目前关于羊草ZIP 家族成员的表达、定位和功能的研究鲜有报道。羊草ZIP 家族成员的挖掘可为了解羊草微量元素吸收和转运的机理以及对日后禾本科作物遗传改良提供参考。
1 材料与方法
1.1 植物材料和培养
将野生羊草种子(于2018 年采集自内蒙古和林格尔县)置于无菌水中,4 ℃处理3 d 后,收集饱满种子进行表面消毒(浸入5% NaClO+0.1% Triton X-100 中,置于摇床150 r·min-1震荡2 h 后用无菌水洗3~5 次)。此后将无菌的羊草种子点播于MQA-CK 培养基(5 mmol·L-1KNO3、1 mmol·L-1CaCl2、1 mmol·L-1MgSO4、1 mmol·L-1H3PO4、1/2 MS Fe 盐、1/2 MS 微量、5 mmol·L-14-吗啉乙磺酸、1%蔗糖、1%琼脂糖,pH 5.7[22])。将培养基置于生长室,黑暗中培养5 d 后进行光照培养(光强度:100 μmol·m-2·s-1;光周期:12 h 光照/12 h 黑暗;温度:22~25 ℃)。
1.2 生物信息学分析
用“zinc”“zinc transporter”或“Zn2+”在野生羊草转录组数据库(本实验室测定)中筛选注释为锌相关的基因,最后筛选到Lc206852。通过植物基因组网站(https://phytozome-next.jgi.doe.gov/)中BLAST 查找Lc206852的同源蛋白,基于Lc206852 及同源蛋白利用https://www.genome.jp/tools-bin/clustalw 进行多重比较并构建系统进化发生树。利用TMHMM(http://www. cbs.dtu.dk/services/TMHMM-2.0/)进行跨膜域预测。
1.3 亚细胞定位
将光照培养7 d 的羊草幼苗速冻研磨,利用TransZol Up Plus RNA Kit(北京全式金生物技术有限公司)提取RNA,使用Hifair Ⅱ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus) (上海翊圣生物科技有限公司)合成cDNA 并去除基因组DNA。利用引物LcZIP1-GFP_F-5′-CGGGGTACCATGGCAGCAGCCA GTCTCAAG-3′和LcZIP1-GFP_R-5′-CCGCTCGAGGGTGCCCATACGGCAAGCATTGAC-3′(加粗字母为限制性内切酶酶切位点,分别为KpnⅠ和XhoⅠ),使用大连宝生物工程有限公司的PrimeSTAR HS DNA 聚合酶从羊草cDNA 中扩增不含终止密码子的Lc206852全长编码序列。通过TA 克隆将PCR 产物连接到pEASYBlunt Simple Cloning Vector(北京全式金生物技术有限公司)后进行测序验证。将测序正确的Lc206852连接到pENTR 3C(Thermo Fisher)的KpnⅠ和XhoⅠ位点,通过Gateway 克隆将不含终止子的Lc206852连接到植物双元表达载体pGWB605[23]的花椰菜花叶病毒(cauliflower mosaic virus,CaMV)35S 启动子下游,绿色荧光蛋白(green fluorescent protein,GFP)编码基因上游,最后将pGWB605-Lc206852转化农杆菌GV3101。将转基因农杆菌和含内质网标记蛋白ER mcherry CD3-959 的农杆菌(购自拟南芥生物资源中心)共同注射到本氏烟草叶片下表皮[4,24]。同时采用聚乙二醇(polyethyleneglycol,PEG)介导法将重组质粒pGWB605-Lc206852转入羊草叶原生质体[4,25]。
采用蔡司LSM 710 共聚焦显微镜(德国)和LSM880 超高分辨率共聚焦显微镜(德国)观察GFP 和mCherry荧光。
1.4 荧光定量PCR
将光照培养7 d 的羊草幼苗移入MQA-CK 液体培养液中,生长14 d 后置于含不同浓度锌和铁的MQA-CK 液体培养液中处理2 d。处理液分别为:对照(MQA-CK)、缺锌(不含Zn2+的MQA,即MQA-Zn2+)、高锌(MQA+150 μmol·L-1Zn2+)、缺铁(不含Fe2+的MQA,即MQA-Fe2+)和高铁(MQA+500 μmol·L-1Fe2+)。参照1.3 分别提取羊草叶和根的总RNA 并合成cDNA。以cDNA 为模板,使用上海翊圣生物科技有限公司的实时荧光定量PCR 试剂盒Hieff UNICON Universal Blue qPCR SYBR Green Master Mix 和TOWER 2.2 荧光定量梯度PCR 仪(德国)进行qRT-PCR 分析,重复3 次。qRT-PCR 扩增引物为LcZIP1-RT_F-5′-CGTCGACACCATCGCCACGG-3′,LcZIP1-RT_R-5′-CACCGAGTGCACTATGATCCCCAGC-3′。以LcACTIN2作为内参,采用2-ΔΔCt算法[4]分析该基因的相对表达量。扩增体系:总体积为20.0 μL,其中Hieff UNICON® Universal Blue qPCR SYBR Green Master Mix(1×):10.0 μL;上游引物(0.2 μmol·L-1):0.4 μL;下游引物(0.2 μmol·L-1):0.4 μL;cDNA(200 ng·μL-1):1 μL;ddH2O:8.2 μL。反应条件:95 ℃预变性2 min;95 ℃变性10 s;60 ℃退火30 s;72 ℃延伸30 s;40 个循环。熔解曲线程序为60~95 ℃,每15 s 升温1 ℃。
1.5 酵母功能互补试验
使用引物LcZIP1-pYUL2_F-5′-GCTCTAGAATGGCAGCAGCCAGTCTCAAGC-3′;LcZIP1-pYUL2_R-5′-CCCAAGCTTTCATGCCCATACGGCAAGCATTGAC-3′(加粗字母分别为XbaⅠ和HindⅢ酶切位点)和PrimeSTAR HS DNA 聚 合 酶(Takara,大 连)将Lc206852克隆入酵母表达载体pYUL2,利 用Yeast Transformation System 2(CLONTECH)转化高锌敏感酵母突变体∆zrc1/∆cot1(BY4741,MATa zrc1::Ura1 cot1HIS,由华中农业大学林会副教授惠赠)。利用引物LcZIP1-pFL61_F-5′-ATTTGCGGCCGCGGTACC ATGGCAGCAGCCAGTCTCAAG-3′和LcZIP1-pFL61_R-5′-ATTTGCGGCCGCTCATGCCCATACGGC AAGCATTG-3′(加粗字母分别为KpnⅠ和NotⅠ酶切位点)扩增Lc206852,将Lc206852克隆入酵母表达载体pFL61。利用上述方法转化低锌敏感酵母突变体∆zrt1/∆zrt2(ZHY3;MATα ade6 can1 his3 leu2 trp1 ura3 zrt1::LEU2 zrt2::HIS3)和低铁敏感酵母突变体∆fet3/∆fet4(DEY1453;MATa can1his3leu2trp1 ura3∆fet3::HIS3∆fet4::LEU2)(由美国Missouri 大学David Eide 教授惠赠)。将转基因酵母的OD600值调至1.0,然后进行梯度稀释,分别为10-1、10-2、10-3、10-4。取10 μL 不同浓度的菌液点样于含高锌[(YPD 培养基:蛋白胨2 g,酵母提取物1 g,加水定容至95 mL,琼脂粉2 g,121 ℃灭菌20 min,待溶液冷却后加入40 %无菌葡萄糖5 mL)+7 mmol·L-1ZnCl2,pH 6.5]、低锌(YPD 培养基+1 mmol·L-1乙二胺四乙酸+50 μmol·L-1ZnCl2,pH 6.5)和低铁[(SD-Ura 培养基:Minimal SD Base 2.67 g,-Ura DO Supplement 0.077 g 于蒸馏水中充分溶解,定容至100 mL,琼脂粉2 g,121 ℃灭 菌20 min)+50 mmol·L-1MES+50 μmol·L-1红 菲 绕 啉 二 磺 酸 (Bathophenanthrolinedisulfonic acid disodium salt hydrate,BPDS),pH 为5.5]的酵母培养基上。30 °C 暗培养3~4 d 后观察并拍照记录。
2 结果与分析
2.1 Lc206852 同源蛋白系统发育分析和跨膜域分析
通过Phytozome 中的BLAST 分析,发现Lc206852 与拟南芥AtZIP1 同源,因此将其命名为LcZIP1。选取亲缘关系较近的禾本科5 属8 种植物的ZIP1 同源蛋白进行系统发生分析,发现LcZIP1 与禾本科短柄草属(Brachypodiumspp.)的Brachypodium stacei和二穗短柄草(Brachypodium distachyon)亲缘关系最近(图1A)。跨膜域分析发现羊草LcZIP1 含347 个氨基酸,有9 个跨膜结构域(transmembrane domain,TM),其中膜内和膜外各有5 个非跨膜域短肽(图1B)。高粱(Sorghum bicolor)、柳枝稷(Panicum virgatum)、狗尾草(Setaria viridis)、谷子、二穗短柄草和B. stacei含9 个跨膜结构域,羊草LcZIP1 与其结构类似;而哈氏黍(Panicum hallii)与LcZIP1 不同,其只含7 个跨膜域,其中第5 和6 个跨膜域缺失,第4 个跨膜域之后的非细胞质面非跨膜域短肽长度长于LcZIP1(图1C)。ZIP1 在不同的植物中存在一定程度的变异。
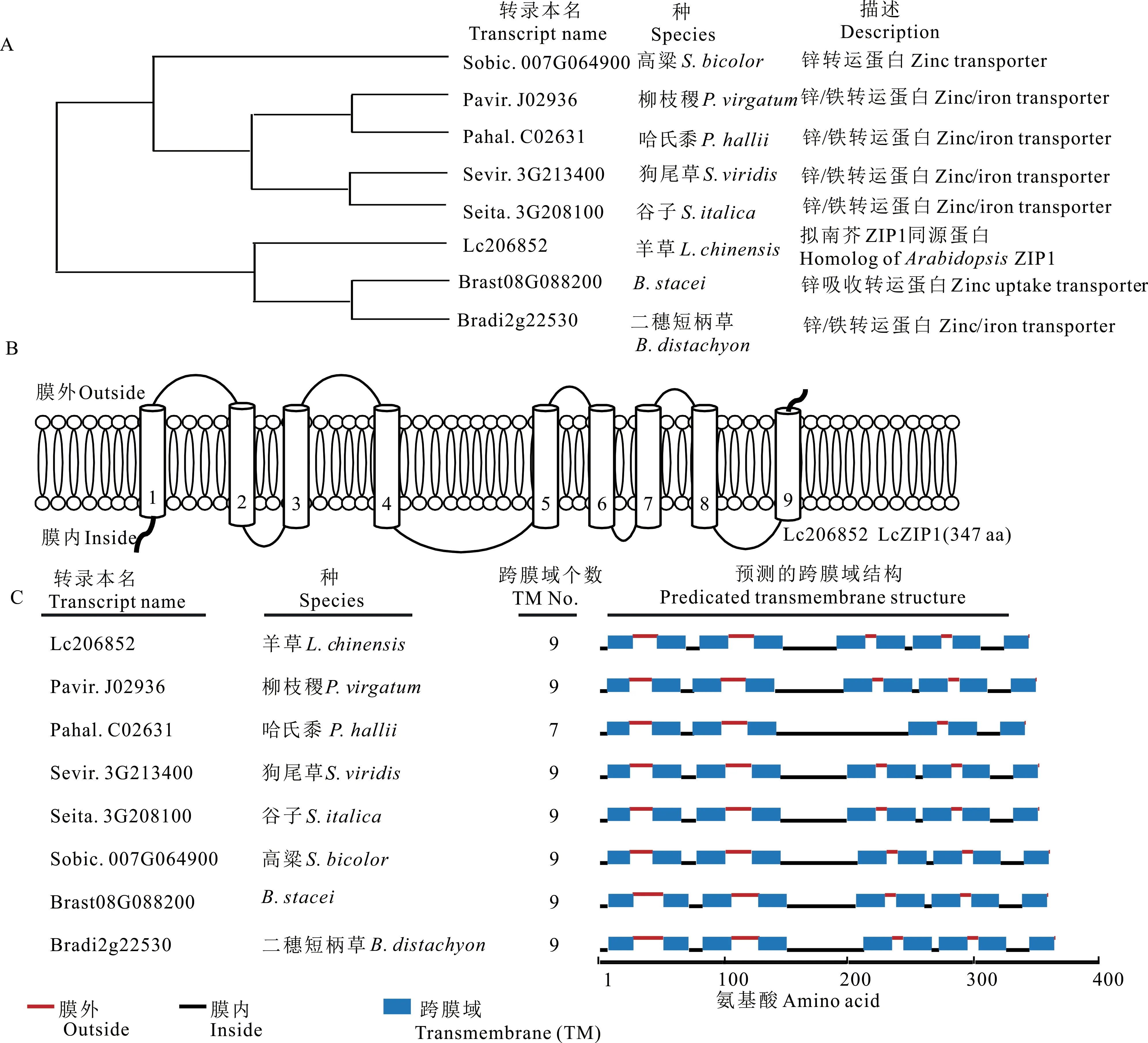
图1 LcZIP1 同源蛋白系统进化发生分析和跨膜域Fig.1 Phylogenetic analysis and transmembrane domains of LcZIP1 homologues
2.2 LcZIP1 亚细胞定位于内质网
观察烟草下表皮细胞异源共表达LcZIP1-GFP和内质网标记蛋白发现,LcZIP1 的绿色荧光呈网状结构,内质网标记蛋白的红色荧光同样呈网状结构,并且绿色荧光和红色荧光融合,呈黄色荧光,表明在烟草细胞中LcZIP1 定位于内质网(图2A)。观察LcZIP1和内质网标记蛋白在羊草叶肉细胞原生质体中的共表达也发现,LcZIP1 的绿色荧光也和内质网标记蛋白的红色荧光融合(图2B),表明LcZIP1 定位于羊草叶肉细胞的内质网上。
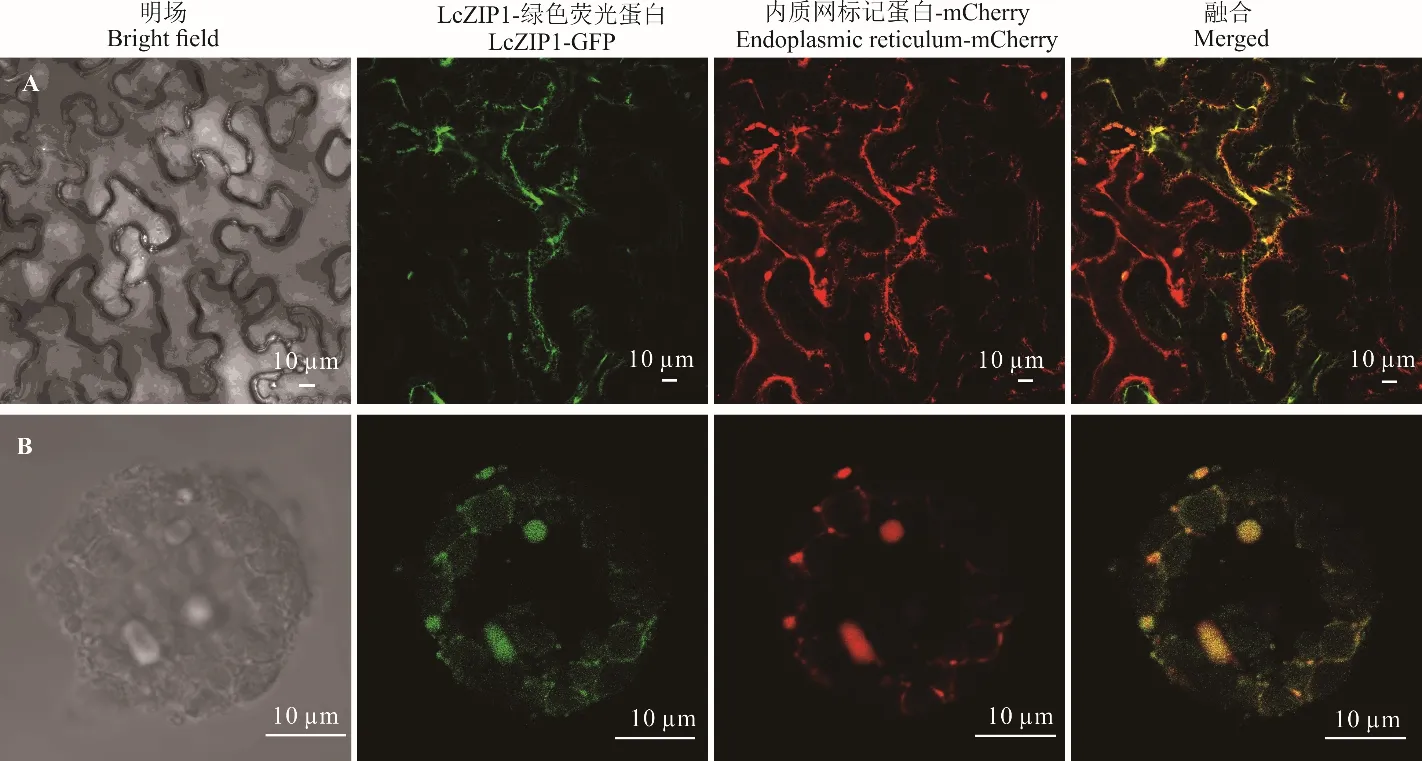
图2 LcZIP1 定位于内质网Fig. 2 LcZIP1 was located in endoplasmic reticulum
2.3 LcZIP1 响应高锌、低铁和高铁条件
实时荧光定量PCR 结果表明,LcZIP1在羊草根和叶中均有表达。在缺锌条件下,LcZIP1在根和叶中的表达量无显著差异。在高锌、缺铁和高铁条件下,叶中LcZIP1的表达量无显著变化,但根中LcZIP1的表达量均显著上调(图3)。在高铁条件下,根中的表达量虽然上调但显著低于缺铁条件。结果表明,羊草根细胞中的LcZIP1参与对高锌、低铁和高铁条件的响应。
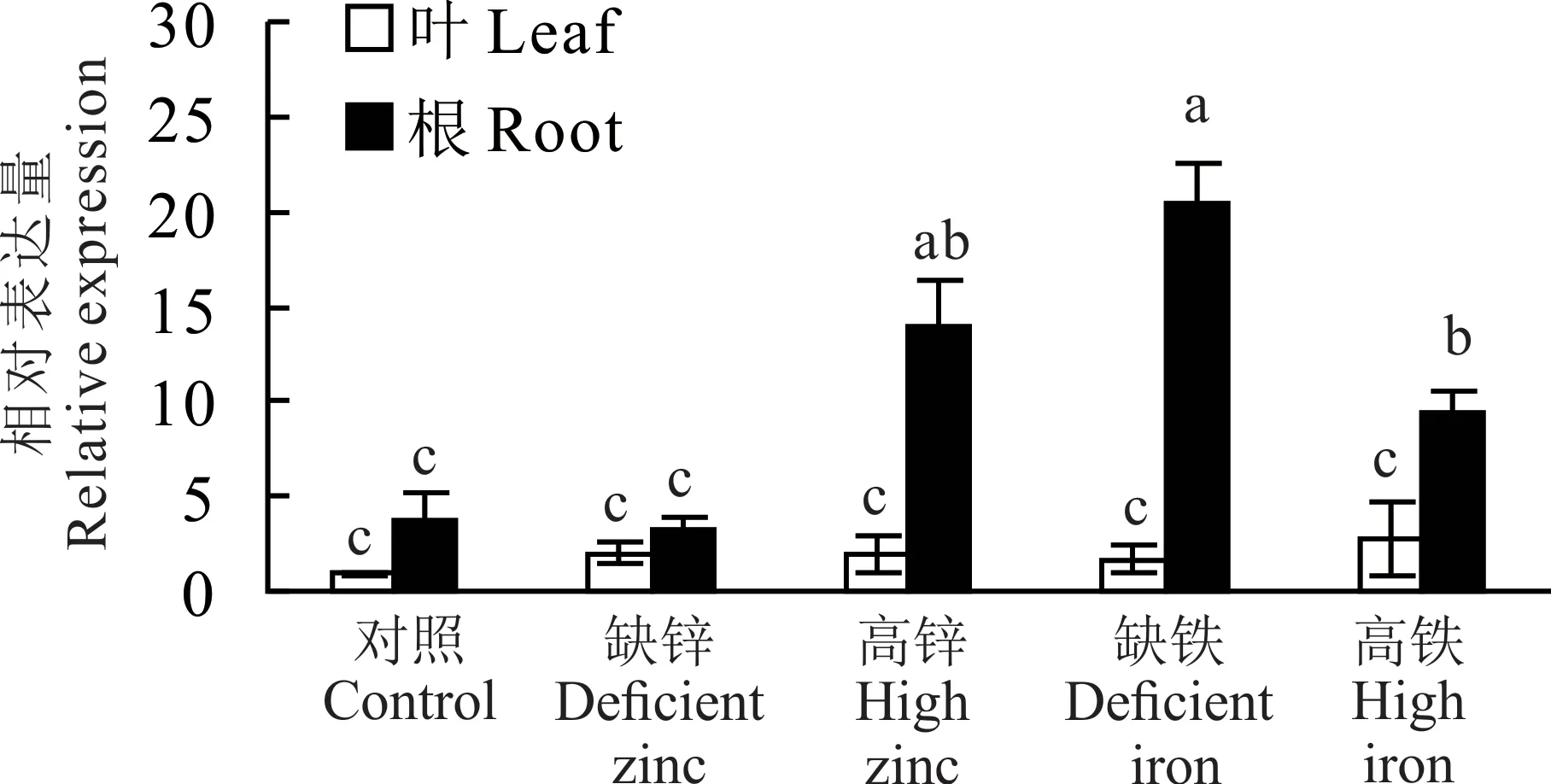
图3 不同Zn2+和Fe2+浓度条件下LcZIP1 的相对表达量Fig. 3 Relative expression of LcZIP1 under different Zn2+and Fe2+ concentration
2.4 LcZIP1 具有Fe2+转运功能
在低铁条件下,LcZIP1能够恢复低铁敏感酵母突变体∆fet3/∆fet4的生长表型,与已报道的阳性对照AtZIP7表型一致(图4A)。据报道,AtZIP7可以使∆fet3/∆fet4在低铁条件下恢复生长[8],表明其具有铁转运功能。

图4 LcZIP1 在酵母突变体∆fet3/∆fet4、∆zrt1/∆zrt2 和∆zrc1/∆cot1 中的功能互补验证Fig. 4 Complementation analysis of Saccharomyces cerevisiae mutants (∆zrt1/∆zrt2, ∆fet3/∆fet4 and ∆zrc1/∆cot1) with LcZIP1
在低锌条件下,LcZIP1不能恢复低锌敏感酵母突变体∆zrt1/∆zrt2(锌吸收缺陷酵母突变体)的生长表型(图4B),与已报道的阳性对照AtZIP7(该基因具有锌转运功能)不同,因为AtZIP7可以使∆zrt1/∆zrt2在低锌条件下恢复正常生长。
在高锌条件下,LcZIP1不能增强∆zrc1/∆cot1对高锌的抗性,所以不能在高锌培养基上生长(图4C),与已报道的阳性对照MtMTP3(该基因具有锌转运功能)不同。MtMTP3能够增强∆zrc1/∆cot1对高锌的抗性,在高锌培养基上可正常生长[26]。
以上酵母功能互补试验表明,LcZIP1具有Fe2+转运功能,无Zn2+转运功能。
3 讨论
前期研究表明植物ZIP 家族成员由309~476 个氨基酸组成,含8 个跨膜域,氨基端和羧基端位于质膜外,第3 和4 跨膜域之间的短肽长度变异度大,命名为变异区(variable region),该区富含组氨酸,被认为是潜在的金属结合域[27]。后来发现,水稻OsZIP1 有9 个跨膜域,氨基端位于膜内,羧基端位于膜外[16],但OsZIP1 的锌结合域尚未明确;蒺藜苜蓿6 个ZIP 家族成员(MtZIP1, 3, 4~7)由350~372 个氨基酸组成,有8 个跨膜域,第3 和4 跨膜域之间也为富含组氨酸的短肽[11]。本研究发现羊草LcZIP1 由347 个氨基酸组成,含9 个跨膜域,氨基端位于膜内,羧基端位于膜外,与OsZIP1 相似。在高粱、柳枝稷、狗尾草、谷子、哈氏黍、二穗短柄草和B. stacei的ZIP 中除哈氏黍ZIP 有7 个跨膜域,其余都具有9 个跨膜域。另外这些ZIP 的第4和5 跨膜域之间短肽长度变异度大,为54~108 个氨基酸(图1C)。羊草LcZIP1 第4 和5 跨膜域之间短肽长44 个氨基酸,序列为QRAAHAKKAAAVVGADV EATPAHHAHGVSAAIASSSDAQLIRHR,含5 个组氨酸,其他非跨膜域不含组氨酸。表明这段短肽可能代表金属结合域,但需进一步证明。此外还发现,以上8 种禾本科植物(包括羊草在内)中,除哈氏黍外,其余植物的ZIP1 都具有9 个跨膜域。另外,8 种植物ZIP1 的氨基端和羧基端都分别位于膜内和膜外(图1)。
目前已发现植物ZIP 家族可以转运Fe2+、Zn2+、Cu2+、Mn2+、Ni2+或Cd2+。这些元素都属于重金属,其中绝大多数(Fe2+、Zn2+、Cu2+、Ni2+和Mn2+)还属植物生长必需微量元素,因此对该家族成员的挖掘有助于了解Fe2+、Zn2+、Cu2+、Mn2+、Ni2+和Cd2+对促进农作物生长或富集土壤重金属的分子机制,也可为农作物的改良和土壤重金属污染防治提供依据。羊草是内蒙古草原的重要建群种,抗逆性强,不仅是优质的天然牧草还是重要的生态草。羊草ZIP 家族成员的挖掘和功能鉴定,对于了解羊草对必需重金属(Fe2+、Zn2+、Cu2+、Mn2+、Ni2+)和非必需重金属(Cd2+)吸收和转运的分子调控机理具有重要的理论意义,也为牧草和其他农作物的改良和被重金属污染的土壤生态修复提供参考。已有研究表明将拟南芥AtZIP1在非洲重要主食木薯(Manihot esculenta)中过表达可显著提高木薯块根的锌含量[28]。将拟南芥ZIP 家族成员IRT1和编码铁蛋白(ferritin)的基因FER1在木薯中共表达,田间试验数据表明转基因木薯产量不受影响,但Fe 的积累水平提高了7~18 倍,Zn 提高了3~10 倍。该研究为改善全球人口的营养提供了助力[29]。有些ZIP 成员具有金属解毒的功能,如OsZIP1 定位于内质网和质膜上,在过量的锌、铜和镉的条件下,OsZIP1被诱导表达,因此可阻止Zn、Cu 和Cd 在水稻中过量积累[16]。
本研究发现LcZIP1 定位于内质网,高锌、高铁和缺铁条件下,根中的LcZIP1表达上调;酵母功能互补试验发现,LcZIP1 参与Fe2+转运,但其是将Fe2+转运至内质网进行贮存还是排出内质网尚需进行深入研究。另外,现已报道ZIP 家族成员在拟南芥、水稻、小麦、大麦和菜豆中分别有15、17、14、12 和23 个[15],羊草ZIP 家族中有多少成员还需进一步的研究。
猜你喜欢
解放军医学杂志(2021年12期)2022-01-18 03:53:24
现代畜牧科技(2021年7期)2021-07-28 06:40:30
现代畜牧科技(2021年4期)2021-07-21 06:13:04
现代临床医学(2021年1期)2021-01-26 00:55:52
安徽医科大学学报(2016年12期)2017-01-15 14:21:55
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
现代畜牧科技(2016年9期)2016-02-21 12:25:04
中国当代医药(2015年33期)2015-03-01 02:09:08
山东医药(2015年40期)2015-02-28 14:28:45
