
马铃薯杂交后代表型性状的遗传变异分析
2023-09-24 07:40:28李雅萍
种子 2023年7期
李雅萍, 王 舰,2,3,4, 王 芳,2,3,4
(1.青海大学农牧学院, 西宁 810016; 2.青海大学农林科学院, 西宁 810016;3.青海大学,青藏高原生物技术教育部重点实验室, 西宁 810016;4.青海省农林科学院,青海省马铃薯育种重点实验室, 西宁 810016)
马铃薯(SolanumtuberosumL.)是全球继水稻、玉米、小麦后的第四大粮食作物,也是最重要的块茎类作物之一[1]。杂交育种技术是基于双亲的优良性状产生一定的变异群体,并在其中筛选而获得优良品系的过程,是当前马铃薯品种选育的有效途径[2-3]。马铃薯杂交亲本一般选择综合性状优良、育种目标性状需要改良的品种,或者是经过验证,子代能产生优良后代的育种材料,通常要求亲本性状互补[4]。马铃薯亲本受遗传、环境和人为等因素的影响,育种周期长、效率低[5],为了能够在杂交后代中早期鉴定出具有优良性状的子代,快速选育出具有目标性状的新品种,这就需要了解马铃薯性状在后代中的分离和遗传变异规律,为新品种的选育提供参考。
马铃薯杂交后代的分离和遗传变异规律可以从形态学水平和分子水平进行分析。形态学指标是植物分类和品种鉴别的传统方法[2],孙邦升等[6]对国家马铃薯种质资源克山试管苗库中179份鲜食高产马铃薯种质资源的29个质量性状和7个数量性状进行遗传多样性分析,筛选出可以用于早熟育种、薯形和株型的改良以及选育晚熟高淀粉的资源。杨春和齐海英[7]对田间采集的种质资源的11个质量性状和10个数量性状,进行Shannon-Wiener指数、相关性、主成分与聚类分析,认为各个性状都具有较高的遗传多样性,并将种质资源分为中早熟低产型、中早熟高产型、中晚熟高产型、中晚熟低产型四大种质群。余斌等[8]对119份从秘鲁国际马铃薯中心引进的马铃薯材料的10个表型性状进行遗传多样性分析及综合评价,发现生育期具有最丰富的遗传多样性,并筛选出对马铃薯种质资源表型性状综合值具有显著影响的几个指标,这些指标可用于旱作条件下对马铃薯种质资源的综合评价。张凤军等[9]分析大西洋和定薯1号的杂交后代在干旱调控下抗旱相关性状的遗传变异时发现,主茎数、分枝数、单株结薯数等性状为微效多基因控制的数量性状,现蕾期的株高和茎粗、盛花期的株高和茎粗等性状表现为不完全显性,单株结薯数在不同环境中分离特征有差异,干旱条件下表现为不完全显性,正常灌溉时高值为显性,低值为隐性,说明该性状受环境影响较大。在分子水平上国内很多学者已利用SSR、SRAP等标记对马铃薯的遗传多样性进行了大量研究[10-13]。本研究用青薯9号作为母本,分别与大西洋、 品系12-9-33杂交,获得杂交后代群体作为试材,从群体遗传学角度对杂交后代的若干性状进行遗传多样性分析,探究杂交后代主要表型性状的遗传变异规律。
1 材料与方法
1.1 实验材料
供试材料为116份青薯9号(♀)×大西洋(♂)的杂交后代(以下简称组合1)和425份青薯9号(♀)×品系12-9-33(♂)的杂交后代群体(以下简称组合2)。
1.2 实验方法
1.2.1性状的收集
1) 描述型性状的测定
参照国标《植物新品种特异性、一致性和稳定性测试指南(马铃薯)》,采用直接观察法评定描述型性状,具体包括植株类型、植株生长习性、茎花青苷显色强度、茎翼波状程度、复叶大小、小叶密集程度、小裂叶数量、复叶绿色程度、复叶主脉花青苷显色强度、小叶联会频率、小叶边缘波状程度、小叶光泽度、块茎形状、芽眼深度、表皮光滑度、薯皮颜色、薯肉颜色、芽眼基部颜色,共18个指标。各个观测性状表达状态和赋值标准见表1。

表1 描述型性状的遗传变异分析Table 1 Analysis of genetic variation of descriptive traits
2) 数值型性状的测定
干物质、蛋白、淀粉、粗纤维、还原糖、Vc等指标通过近红外品质分析仪测定。
单株结薯数:收获后,每个植株上形成的所有块茎数。
单株产量:收获后,每个植株上形成的所有块茎的重量。
1.2.2数据分析
利用Excel和SPSS软件进行描述统计、频率分析以及变异系数的确定,对数值型性状进行质量化处理,然后利用Past4.09软件计算 Shannon-Weaver 遗传多样性指数(H′),以皮尔逊相关系数度量各表型性状间相关方向和相关程度大小,通过主成分分析挑选优株,根据杂种优势理论和相对遗传力分析表型性状的遗传模型,各性状遗传参数分别以变异系数、中亲优势率、超亲优势率、父母本的相对遗传力表示,各参数计算公式如下[14]:

H′=-∑PilnPi;
a1=(Fm-LP)/(HP-LP);
a2=1-a1,
式中:S表示标准差,Fm表示杂交F1代某一性状的平均值,H′表示遗传多样性指数,MP(中亲值)表示亲本性状的平均值,HP表示双亲中较大的亲本值,LP表示双亲中较小的亲本值,a1为大值亲本的相对遗传力,a2为小值亲本的相对遗传力。
2 结果与分析
2.1 后代表型性状的离散特征
由表1可知,组合1杂交后代的18个描述型性状平均多样性指数为1.08,变化范围为0.58~1.57,其中薯肉颜色的遗传多样性指数最高,为1.57,说明薯肉颜色在116个后代中遗传多样性最为广泛,其次为茎部花青苷显色程度,为1.51。18个性状中,有11个性状的多样性指数超过1。组合2的杂交后代中18个描述型性状平均多样性指数为1.17,变化范围为0.76~1.49,其中茎部花青苷显色程度最高,为1.48,其次为块茎形状(1.48),只有2个性状的多样性指数小于1。综合来看,两个组合杂交后代描述型性状都表现出显著的形态多样性,且各性状的遗传多样性指数在两个组合中的排名大致相同。薯肉颜色、茎花青苷显色程度、块茎形状变异丰富度都很高,小叶联会频率和表皮光滑度的遗传多样性较低。8个数值型性状测定结果见表2,两个杂交组合各性状间的差异较大,组合1杂交后代各性状的遗传多样性范围为1.07~1.69,组合2各性状的遗传多样性范围为1.38~1.62,其中淀粉含量变异最为丰富,在两个组合中分别为1.69和1.64,但组合2各性状的遗传多样性总体大于组合1,说明组合2的变异更丰富。

表2 数值型性状的遗传变异分析Table 2 Genetic variation analysis of quantitative traits
变异系数可以体现出该性状是否具有遗传稳定性,变异系数越大,说明性状的离散程度越大,其遗传背景越丰富,越具有选择优株的潜力[15],由表1可见,组合1的18个描述型性状的变异系数平均值为35%,范围在26%~94%之间,组合2的18个描述型性状的变异系数平均值为38%,范围在21%~86%之间,说明这两个组合的杂交后代都具有极广泛的变异性。复叶绿色程度的变异在两个组合中都表现为最低,主脉花青苷显色程度在两个组合中都表现为最高。在8个数值型性状中,单株结薯数、单株产量、还原糖含量变异较丰富,蛋白、干物质、粗纤维、淀粉、Vc的变异系数都很低,基本小于10%。
亲本表现及杂交后代描述型性状的分布情况如表1所示,两个组合的亲本有3个性状表现完全一致,分别是中间型的植株类型、小叶联会频率低、芽眼浅。所有杂交后代的茎花青苷显色程度、茎翼波状程度、复叶主脉花青苷显色程度均以介于双亲过渡型数目居多,复叶绿色程度、小叶联会频率均与父本或母本相同。虽然两个组合有共同的母本,但某些性状在不同组合后代中的分离规律并不相同,例如,组合2的薯肉颜色、薯皮颜色、块茎形状以及组合1的小叶光泽度表现为与父本或母本一致,而组合1的薯肉颜色、薯皮颜色、块茎形状以及组合2的小叶光泽度大部分都处于双亲之间,或偏向于父本,或偏向于母本。茎花青苷显色程度、蛋白含量、淀粉含量在两个组合中的平均值都介于高亲值与中亲值之间,说明这些性状表现为不完全显性。小叶联会频率、单株结薯数、干物质、芽眼深度的平均值高于高亲值,说明这些性状高值为显性,低值为隐性。
数值型性状的分布如表2所示,同一性状在不同组合中的分离规律基本相同,杂交后代的单株结薯数高于高亲值,说明单株结薯数高值为显性,低值为隐性,其余性状大部分都处于双亲之间,为不完全显性,其中单株产量和还原糖接近于低值亲本,干物质、蛋白、淀粉、Vc、粗纤维含量接近于高值亲本。由图1可知,干物质、蛋白、淀粉、粗纤维基本都呈良好的正态分布趋势,说明这些性状可能都是由多个基因控制的数量性状。

注:A为组合1;B为组合2。图1 数值型性状的分布频率Fig.1 Distribution frequency of quantitative traits
2.2 相关性分析
马铃薯的产量和品质不是由单一的某个性状决定的,它受到多种生长形态的影响,如单株结薯数、株高,茎粗等,且不同的性状对产量与品质的影响程度也不尽相同。相关性分析可以研究各性状之间的相关方向以及相关程度。由图2可知,2个组合杂交后代26个性状之间的相关性呈现高度的复杂性。组合1中地上部分各性状之间、地下部分各性状之间、以及地下部分与地上部分性状之间都呈现复杂的相关性,例如小叶光泽度与茎翼波状程度、复叶大小、小叶密集程度呈显著正相关,与小裂叶数量、复叶绿色程度极显著正相关,与复叶主脉花青苷显著负相关;干物质含量与粗纤维极显著负相关,与Vc、淀粉、蛋白含量极显著正相关,单株产量与干物质、蛋白、淀粉极显著负相关;复叶主脉花青苷显色程度与表皮光滑度显著正相关,与表皮颜色、单株产量极显著正相关,与干物质、淀粉显著负相关,与蛋白含量极显著负相关,植株生长习性与粗纤维、Vc含量、薯肉颜色显著负相关。
组合2的相关性更倾向于地上部分与地上部分、地下部分与地下部分之间的相关性。例如植株类型与除复叶大小、小叶密集程度、小叶联会频率以外的所有地上部分的性状之间极显著相关,茎花青苷显色程度与除复叶大小、小叶密集程度以外的所有地上部分的性状极显著正相关。粗纤维与干物质、蛋白、淀粉、Vc含量极显著负相关,Vc含量与干物质、蛋白、淀粉含量极显著正相关,表皮颜色与芽眼基部颜色、薯肉颜色极显著正相关,单株产量与单株结薯数极显著正相关。也有个别地上部分与地下部分相关的性状,如茎花青苷显色程度、复叶主脉花青苷显色程度分别与表皮颜色和芽眼基部颜色极显著正相关,干物质与茎翼波状程度极显著负相关。
2.3 优良单株的筛选
2.3.1优良单株的初选
据《加工用马铃薯流通规范(SB/T 10968—2013)》中影响马铃薯加工品质的关键因子:干物质、淀粉、还原糖含量,对541个杂交单株进行初步筛选,筛选的标准如下:20%≤干物质含量≤25%;淀粉含量≥16%;还原糖含量≤0.25%,共筛选出70个单株,其中11株来自于组合1,59株来自于组合2。
2.3.2主成分分析
少数几个性状表现最优的单株不一定所有的性状都表现优良,为了选出综合性状最优的单株,对经过初步筛选后的70个优株的 26个性状进行主成分分析。由表3可知,前3个主成分的特征值均大于 1,且累积贡献率达 79.42%,表明这3个主成分可以反映出70个单株26个性状79.42% 的信息。第一主成分反映地上部分的形态、长势等方面的信息。其中,对第一主成分贡献率较大的性状是茎花青苷显色程度、植株生长习性、小叶光泽度、茎翼波状程度、复叶绿色程度,累积贡献率为62.62%;对第二主成分贡献率较大为复叶有关性状、块茎芽眼以及表皮性状,贡献率为12.72%;对第三主成分贡献率较大的为产量方面的性状,累积贡献率为3.88%。
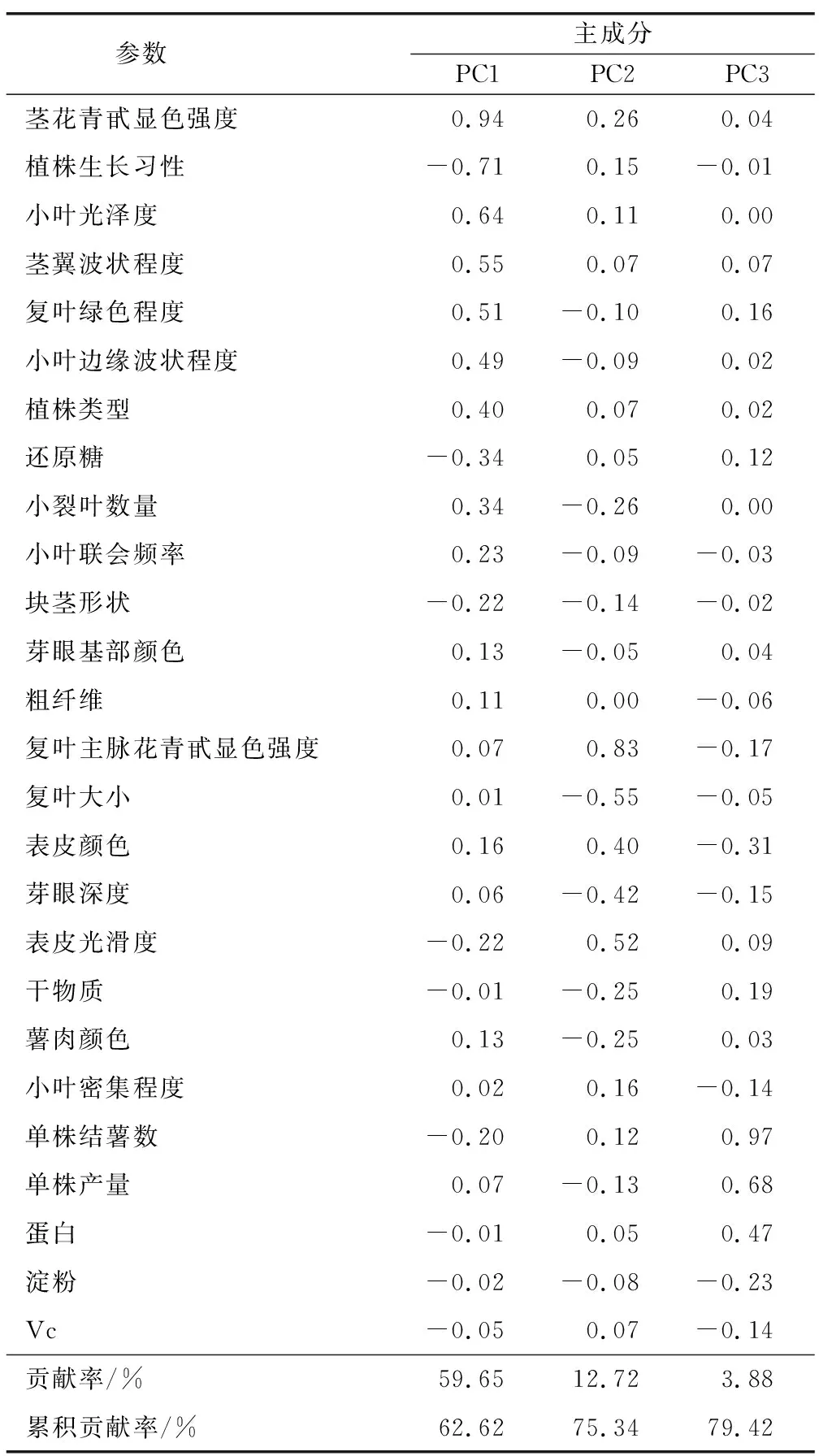
表3 主成分旋转成分矩阵Table 3 Principal component rotation component matrix
为进一步分析各个杂交后代的综合得分情况,利用提取的3个主成分计算出综合主成分值,对各单株进行综合评价比较,排名前10的单株见表4。排名为第五和第六的单株来自于组合1,其余8个单株都来自于组合2。根据综合得分值以及主成分得分值,可以得出不同植株的优劣以及各优株的突出性状。综合排名第一的2-348的PC1、PC2、PC3值比较高,说明该植株无论是地上部分的长势还是块茎的产量以及外形,表现都很突出。排名第二、三、六、七、八的植株PC1值都很高,说明这5个单株地上部分性状优异。排名第四、八、九、十的植株PC2值较高,另外排名第八的PC3值也较高,说明该单株产量也较好。
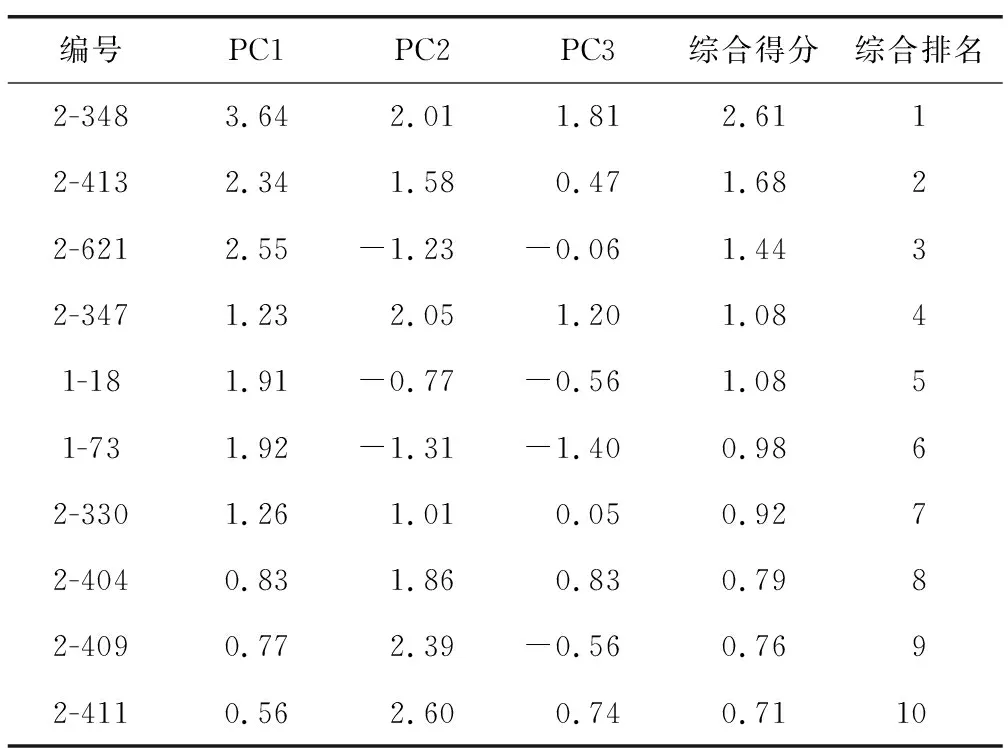
表4 综合评价结果Table 4 Comprehensive evaluation results
2.4 杂交后代表型遗传距离与聚类分析
为了解马铃薯种质资源遗传差异和亲缘关系,本研究利用聚类分析对3个亲本以及70个初步筛选的单株进行分类。对杂交后代中70个初步选择的优株以及3个亲本进行系统聚类,3个亲本与70个单株在遗传距离10.2处分为五大类群(图3),这五大类群的各性状的平均值见表5,植株类型都接近于中间型,生长习性介于直立与半直立之间,小叶密集程度中等,复叶绿色较深,小叶联会频率低,边缘波状程度弱,块茎形状接近于卵圆形,芽眼深度在浅与中等之间,蛋白含量都在2.5%左右,粗纤维含量在1%左右。第Ⅰ类群包括22个单株,占总数的30.14%,其中一个为母本青薯9号,21个个体来自组合2,特点是茎花青甙显色程度强,茎翼波状程度介于中等到强之间,小裂叶数量较多,小叶光泽度较强,表皮光滑,淀粉含量高,还原糖含量低;第Ⅱ类群包括21个单株,占总数的28.8%,2个个体来自组合1,17个个体来自组合2,另外2个为两个组合的父本:大西洋和品系12-9-33,该类群特点是复叶较大,表皮颜色为中等黄色,第Ⅲ类群包括22个单株,占总数的30.14%,其中4个来自组合1,18个来自组合2;第Ⅳ类群包括4个单株,占总体的5.5%,3个来自组合1,1个来自组合2,特点是复叶主脉花青苷显色最强,表皮颜色为部分黄色,干物质含量高(21.85%),Vc含量高(13.38%)。第Ⅴ类群包括4个单株,占总数的5.5%,2个来自组合1,2个来自组合2,特点是芽眼基部颜色为黄色,薯肉颜色为中等黄色,单株结薯数多(28个),单株产量高(2.18 kg)。

注:Q9为青薯9号;DXY为大西洋;9-33为品系12-9-33。图3 70个初步选择的优株以及3个亲本的聚类分析 Fig.3 Clustering analysis of 70 preliminary selected superior strains and 3 parents
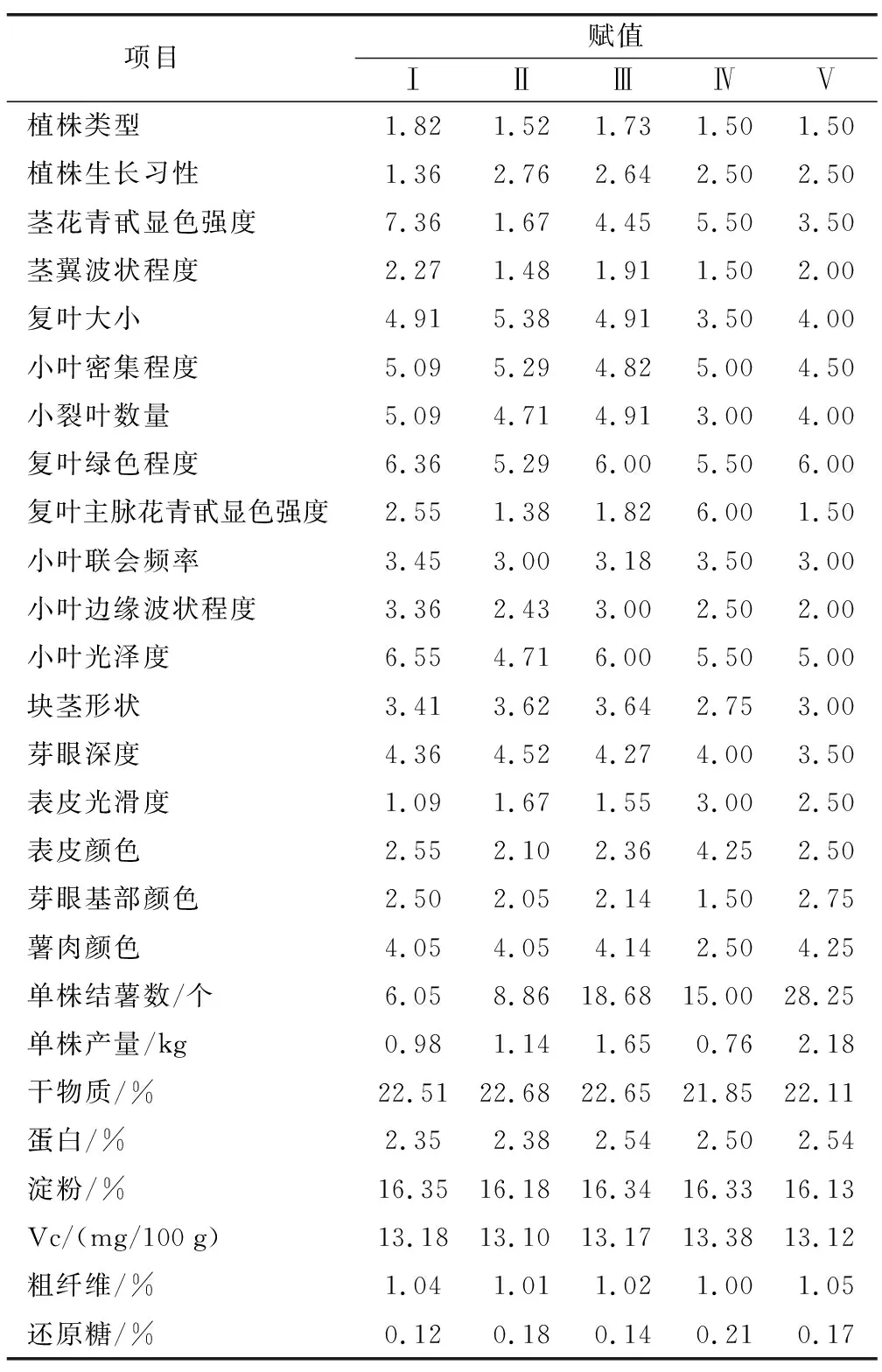
表5 不同类群表型性状赋值Table 5 Assignment values of phenotypic traits in different groups
第Ⅰ类群在茎花青苷显色程度、茎翼波状程度、小叶光泽度方面与第Ⅱ类群差异大,在小裂叶数量、表皮光滑度、还原糖含量上与第Ⅳ类群差异大,在单株结薯数、淀粉含量方面与第Ⅴ类群差异大;第Ⅱ类群在复叶主脉花青苷显色程度、薯皮颜色、Vc含量方面与第Ⅳ类群差别大,在芽眼基部颜色、薯肉颜色、干物质含量方面与第Ⅴ类群差异大。两个组合的亲本分属不同的类别,说明各组合的亲本性状差异较大,符合杂交育种中亲本选配的原则。
2.5 亲本相对遗传力和后代杂种优势
由表6可知,从杂交后代的整体遗传方向来看,组合1有7个性状亲代一致,分别是植株类型、复叶大小、小裂叶数量、小叶联会频率、芽眼深度、表皮光滑度、植株生长习性,有8个性状表现为正向显性,分别是茎花青苷显色程度、经翼波状程度、小叶边缘波状程度、表皮颜色、薯肉颜色、蛋白含量、淀粉含量、Vc含量,有4个性状表现为负向显性,分别是复叶绿色程度、复叶主脉花青苷显色强度、小叶光泽度、块茎形状。从父母本相对 F1代遗传力强弱来看,父本在复叶绿色程度、主脉花青苷显色程度、叶边缘波状程度、小叶光泽度、块茎形状、芽眼基部颜色、单株结薯数、单株产量、粗纤维含量这些性状中相对遗传力强于母本,亲代一致的7个性状无法得到父母本相对遗传力,其余性状母本相对遗传力强于父本。组合2有7个性状亲代一致,分别是植株类型、小叶密集程度、小叶边缘波状程度、块茎形状、芽眼深度、单株结薯数、小叶联会频率,有5个性状表现为正向显性,分别是茎花青苷显色程度、复叶绿色、小叶光泽度、蛋白含量、淀粉含量,有9个性状表现为负向显性;从父母本相对 F1代遗传力强弱来看,父本在茎翼波状程度、复叶大小、复叶主脉花青苷绿色程度、表皮颜色、芽眼基部颜色、单株产量、粗纤维中相对遗传力强于母本,亲代一致的7个性状无法得到父母本相对遗传力,其余性状母本相对遗传力强于父本。

表6 杂交后代表型性状的杂种优势表现Table 6 Heterosis of phenotypic traits in hybrid offspring
组合1杂交后代表型性状大部分表现为正向的杂种优势,有8个性状平均值均超出亲本相应的观测值,表现为正向的超亲优势,其中单株结薯数最显著,其次为芽眼深度,8个性状表现为正向的中亲优势,其中叶边缘波状程度最显著,其次为表皮颜色和薯肉颜色,有5个性状表现为负向超亲优势,小叶密集程度最为显著,5个性状表现为负向中亲优势,单株产量最为显著。组合2杂交后代表型性状大部分表现为负向的杂种优势,并且以负向中亲优势为主,有10个性状以复叶主脉花青苷显色程度最为显著,其次为小叶光泽度,有6个性状表现为负向的超亲优势,其中单表皮光滑度最显著,其次为单株产量、植株类型,4个性状表现为正向的中亲优势,茎花青苷显色程度最显著,6个性状为正向超亲优势,边缘波状程度最显著,其次为单株结薯数、芽眼深度。
3 讨 论
植物表型多样性是遗传多样性和环境多样性的综合体现,对表型性状的研究可以从整体上了解研究对象的多样性程度[16]。利用表型性状来分析植物的遗传多样性能快速了解植物的遗传变异水平,分析其遗传变异规律,预测后代性状[17]。本研究用青薯9号×大西洋和青薯9号×品系12-9-33两个组合的杂交后代为研究材料,对地上部分以及与块茎相关的26个性状通过变异系数和遗传多样性分析,发现这两个组合的杂交后代都具有丰富的遗传多样性,尤其在薯肉颜色、块茎形状、茎花青苷显色程度、淀粉含量、干物质、蛋白等性状中突出,这与赵勤娜等[18]的研究基本一致。以上几个性状的遗传多样性丰富,在一定程度上表明这些性状受环境和遗传控制的影响较大,同时也说明这两个性状变异度较大,遗传改良潜力大,便于新品种的筛选,而蛋白、粗纤维、干物质、Vc的变异程度相对较低,性状较为稳定,受外界环境影响较小。在杂交后代材料中,数值型性状的遗传多样性指数整体高于描述型性状,这与数量性状变异呈连续性的特点是一致的。此外,研究中出现个别性状的变异系数和遗传多样性指数的变化趋势并不一致,如淀粉、蛋白、Vc等的遗传多样性指数较高,但变异系数却很低的情况,这在罗文斌等[19]的研究中也有发现,表明这两个指标的变化趋势并不完全一致。
在性状的频率分析中,本研究发现父母本植株类型均为中间型、联会频率都较低、芽眼浅,在两对杂交组合的 F1代群体中,大部分后代表现都与父母本相同,但也出现了少数不同于亲本的后代,推测这些性状受多基因调控,可能与基因的累加效应有关。金黎平[20]通过对重要农艺性状的遗传效应值分析,发现干物质狭义遗传率低,说明基因累加作用对干物质含量的改良极为重要,且该性状受微效多基因控制。单株结薯数具有极显著的加性、显性和加性环境互作效应,说明基因累加和互作效应对单株结薯数很重要,基因累加作用受环境的影响。本研究8个数值型性状中,干物质、淀粉、蛋白、粗纤维的分布呈现连续正态分布趋势,说明这些性状是由微效多基因控制的数量性状,单株结薯数、单株产量、Vc、还原糖呈现偏正态分布,说明这些性状可能受到某几个主效基因的影响。
植物性状的相关性分析可以研究变量间的密切程度,马铃薯的经济器官是块茎、块茎产量与品质的形成与地上部形态建成等密切相关,地上部分的各性状之间也相互制约[21]。两个组合中单株结薯数和单株产量都呈极显著正相关,且在两个组合中二者都具有较高的相关系数(组合1为0.53,组合2为0.61),这与张荣华等[22]的研究结果一致。王鹏伟[23]在19个马铃薯品种的重要性状及遗传多样性分析中发现,单株产量与干物质、淀粉含量、蛋白质含量呈显著正相关,这与本研究中组合1 性状之间的相关性一致。本研究中,复叶主脉上表面花青苷显色程度与茎花青苷显色强度、薯皮颜色呈极显著正相关,表明营养生长与生殖生长之间存在一定的遗传关系,这与韩志刚等[24]的研究基本一致。因此,在马铃薯的品种选育中,可以通过目标块茎性状与其相关的地上部分性状之间的相关性预测该单株块茎性状的表现,便于亲本的选配,达到定向育种的目的。本研究还发现,组合1中单株产量与干物质、蛋白、Vc含量极显著负相关的状态,说明单产的提高会抑制一些营养物质的储存,这种产量性状与品质性状呈极显著负相关的现象在叶玉珍[25]的研究中也同样出现了,原因可能是在马铃薯性状改良过程中过于注重产量而忽略了品质。
挑选优株是马铃薯育种工作必不可少的环节,本研究根据几个品质性状初步筛选出70个单株,但少数几个指标并不能说明个体的好坏,应将所有指标进行综合评价,挑选出综合性状优良的个体,用于下一步的育种工作[26]。主成分分析能够较好地解释群体方差的主要来源,从而获得解释方差的重要性状并简化研究性状,以便于更好地研究群体。张帆等[27]对20个马铃薯品种的6个性状进行主成分分析,结果表明,供试20个马铃薯品种的品质指标可以用4个主成分来表示,各成分代表性指标分别为蛋白质含量、淀粉含量、可溶性糖含量、还原糖含量。魏康等[28]应用主成分分析法,对反映马铃薯全粉-小麦粉混合面团质构品质的6个指标进行降维分析,对混合面团的质构品质进行综合评定,提取2个主成分,其中主成分1为“硬度因子”,主成分2代表为“弹性因子”。主成分分析还广泛地应用于核桃[29]、大豆[30]、茶树[31]、水稻[32]、小麦[33]等。本研究采用主成分分析法进行马铃薯杂交后代综合性状的评价研究,共提取了3个主成分,这3个主成分可以解释原始信息的79.42%,第一主成分反映地上部分的信息,第二主成分反映块茎表皮的信息,第三主成分反映块茎的产量方面的信息,接着通过70个单株的综合评价值选出了排名前10 的综合品质表现优良的单株,其中包括组合1的2个优良单株和组合2的8个优良单株。通过优良单株的排名以及主成分得分值,也可以得知各个单株的优势性状,进而在之后的改良目标性状的育种工作中得以利用。
在系统聚类时,73个单株被分为五大的类群,其中第Ⅰ、Ⅳ类群的单株在块茎的品质方面表现较好,根据SB/T 10968—2013《加工用马铃薯流通规范》,这两类群的块茎适用于马铃薯加工,例如炸片、炸条,第Ⅴ类群的单株在产量方面表现优异。两个组合的父母本都被分在了两个不同的类群,说明两个组合父母本的性状差异较明显,可在杂交育种及特异资源培育方面加以利用。
杂种优势是指通过杂交方式产生的后代在生长势、抗逆性、繁殖力、适应性、产量以及品质等性状上的表现优于双亲的现象[14]。有效利用杂种优势不但可以提高作物产量,还可改良品质以及增强作物的抗逆性、抗病性,自杂种优势被提出以来,已在很多粮食作物中得到了广泛应用,并取得了巨大的成功。本研究中,2个组合的杂交后代26个表型性状的中亲优势率分别为-45%~47.65%和-29.35%~25.35%。表现为负向中亲优势的性状包括组合1中的复叶绿色程度等,组合2中的复叶主脉花青苷显色程度、小叶光泽度等,这些性状存在具有显性效应的杂种优势。同时,本研究还发现杂交后代中有的性状平均值低于亲中值,如组合1中的植株类型、复叶大小、小叶密集程度、单株产量等,组合2中的植株类型、生长习性、复叶大小等表现出明显的性状衰退现象,杂种优势衰退的现象可能会导致杂种优势的利用受到限制,但依然可以从后代中选择其他性状显著高于双亲的优良单株,从而培育成新品种。因此,在马铃薯杂交育种中,应对此类性状在杂交后代中的衰退表现给予充分估计,以达到选育理想杂交品种的目的。除此之外,两个组合中都存在表现为负向超显性的性状,说明这些性状的遗传特性较复杂,有等位基因间的显性效应、非等位基因间的上位效应或者环境因素的影响在发挥重要作用。
杂交育种是品种改良和种质创新的重要方式,但周期漫长,且需要花费大量的人力物力。加强遗传规律的研究,有助于发展早期鉴定,缩短育种周期,促进成果产出。筛选性状时,可以综合父母本相对遗传力差异、性状本身遗传力强弱等选配杂交亲本,预测后代各性状的表现。本研究从形态学水平分析了马铃薯杂交后代的地上部分有关性状以及与块茎相关的一些性状的遗传多样性,可为马铃薯杂交后代后续筛选奠定基础,也为马铃薯育种过程中进一步改良提供了依据。在此基础上,还可以考虑将其与基因组学结合起来,开展杂交后代早期鉴定,可节约时间和成本,加速育种进程,提高种质创新与利用效率。
猜你喜欢
High Technology Letters(2021年4期)2022-01-09 02:08:16
今日农业(2021年15期)2021-11-26 03:30:27
河北果树(2020年4期)2020-11-26 06:05:00
海峡姐妹(2019年1期)2019-03-23 02:42:40
中成药(2018年9期)2018-10-09 07:18:46
现代园艺(2017年19期)2018-01-19 02:50:01
东北林业大学学报(2016年6期)2016-07-15 10:12:53
九江学院学报(自然科学版)(2015年1期)2015-11-12 03:33:20
云南中医学院学报(2015年2期)2015-07-31 18:11:58
武夷学院学报(2014年5期)2014-07-19 10:08:27
