
一株鱼源致病性嗜水气单胞菌XDMG的全基因组测序及比较基因组分析
2023-09-01 10:42:08郭少华毛会丽刘征权付美媛赵平原马文博李旭东关建义
生物技术通报 2023年8期
郭少华 毛会丽 刘征权 付美媛 赵平原 马文博 李旭东 关建义
(新乡医学院生命科学技术学院, 新乡 453003)
嗜水气单胞菌(Aeromonas hydrophila)属于弧菌科气单胞菌属,是一种条件致病菌,同时也是细菌性败血症的主要病原体,广泛存在于世界各地的各种环境中,如淡水、土壤和水产养殖设施等,是常见的人-兽-鱼共患病病原菌,其不仅能感染草鱼(Ctenopharyngodon idella)、鲢鱼(Hypophthalmichthys molitrix)、罗非鱼(Oreochromis mossambicus)等多种重要经济鱼类,而且还会感染人类,引起腹泻、食物中毒及败血症等多种疾病[1-5]。由于我国水产养殖密度高、过度投喂、环境污染等因素,嗜水气单胞菌引起的细菌性败血症已成为养殖行业的主要问题[6-7]。本实验室在河南新乡某渔场,从自然患病的麦穗鱼(Pseudorasbora parva)体内分离出一株嗜水气单胞菌,命名为XDMG[8], 并对其开展了初步研究。结果表明,嗜水气单胞菌XDMG对健康鲫鱼(Carassius auratus)的半数致死浓度(median lethal dose,LD50)为 1.5×105CFU/mL[9];回归感染试验显示,嗜水气单胞菌XDMG导致银鲫臀鳍、腹鳍、胸鳍的基部及殖泄孔处均出现充血现象, 与细菌性败血症症状一致,感染嗜水气单胞菌XDMG后,发病迅速,存活时间短,说明嗜水气单胞菌XDMG具有较强的致病性[8]。
本研究对嗜水气单胞菌XDMG进行了全基因组测序、基因注释及分析,与4株亲缘关系较近的嗜水气单胞菌[10-13]进行基因组的比较分析,为嗜水气单胞菌基因与蛋白质功能的研究奠定了基础,同时也为嗜水气单胞菌基因工程疫苗的研制奠定了基础。
1 材料与方法
1.1 材料
本试验所用菌株嗜水气单胞菌XDMG来源于自然患病濒死的麦穗鱼体内,现保藏于中国典型培养物保藏中心(保藏号为CCTCC NO: M2013566)。选用LB液体培养基对该菌株进行培养(每 1000 mL含有NaCl 10 g,胰蛋白胨 10 g,酵母提取物 5 g,pH 为 7.2),加入20 g琼脂即为LB固体培养基。
1.2 方法
1.2.1 嗜水气单胞菌XDMG全基因组测序、拼接组装 将嗜水气单胞菌XDMG在LB固体平板上划线分离纯化后,挑取单菌落至LB液体培养基中,在28℃,180 r/mim的条件下进行培养,12 h后取1 mL菌液(1%)接种至100 mL LB液体培养基中,培养16 h之后,采用DNA提取试剂盒提取嗜水气单胞菌XDMG基因组DNA。将提取得到DNA送至上海美吉生物科技有限公司进行全基因组测序。利用软件Newbler[14]对测序得到的原始数据进行组装,经过组装后,对于基因组上仍然存在的缺口,通过设计相应的引物对缺口处的基因序列进行PCR扩增,然后将PCR产物通过Sanger法进行测序,测序结果通过SeqMan[15]与已有的测序结果进行拼接,最终得到完整的基因组序列。
1.2.2 基因功能注释分析 利用Glimmer[16](http://ccb.jhu.edu/software/glimmer/index.shtml)对基因组序列进行开放阅读框(open reading frame, ORF)的预测。通过序列比对工具(basic local alignment search tool, BLAST)在NCBI非冗余蛋白(NR)、注释蛋白质序列(Swiss-Prot, https://web.expasy.org/docs/swissprot_guideline.html)数据库[17]、蛋白质家族(Pfam,http://pfam.xfam.org/)数据库[18]、京都基因和基因组百科全书[19](KEGG, http://www.genome.jp/kegg/)、基因本体(GO, http://www.geneontology.org/)、碳水化合物活性酶数据库[20](CAZy, http://www.cazy.org/)、毒力因子数据库[21](VFDB, http://www.mgc.ac.cn/VFs/)、综合的抗生素抗性基因数据库[22](CARD, http://arpcard.Mcmaster.ca)、病原与宿主互作数据库[23](PHI)对ORF翻译的氨基酸进行注释。蛋白质同源群(COG)通过EggNOG[24](http://eggnog.embl.de/)数据库进行注释,在转运蛋白分类数据库[25-26](TCDB,http://www.tcdb.org/)进行转运蛋白的分析,使用Tmhmm软件[27](http://www.cbs.dtu.dk/services/TMHMM/)获得基因组中跨膜蛋白相关的基因,利用 tRNAscan-SE v2.0[28]软件(http://trna.ucsc.edu/software/)和Barrnap软件(https://github.com/tseemann/barrnap)分别对基因组进行tRNA 与rRNA 的预测。使用Tandem Repeats Finder软件[29]进行串联重复序列预测,可以得到重复序列的位置、重复次数、核酸组成等信息。利用IslandViewer[30]进行基因组岛预测。利用Minced[31]进行CRISPRCas预测。
1.2.3 系统发育和基因组比较 基于保守单拷贝的看家基因采用MEGA 6.0软件选择NJ(neighborjoining)法构建系统进化树。用DNAMAN软件进行核酸及氨基酸序列差异性分析。
2 结果
2.1 嗜水气单胞菌XDMG基因组拼接
嗜水气单胞菌XDMG经全基因组测序共获得5355 403个有效反应,基因组覆盖度为98.27%。经过测序数据质控和序列组装之后, 共计获得重叠群(contigs)47个,共形成47个基因组骨架(scaffolds)(表 1)。
2.2 嗜水气单胞菌XDMG基因组的特性
嗜水气单胞菌XDMG基因组是大小为4992 026 bp的环状染色体(图1),GC含量为60.84%, 该基因组包含了4935个编码基因。在嗜水气单胞菌XDMG中共发现99个tRNA 序列,转运20种氨基酸,每种氨基酸至少对应1个tRNA基因,其中异亮氨酸对应的tRNA数最高,在嗜水气单胞菌XDMG的基因组中12个tRNA在转运异亮氨酸。同时也找到3个rRNA 序列,23S rRNA、16S rRNA 和5S rRNA各1个。
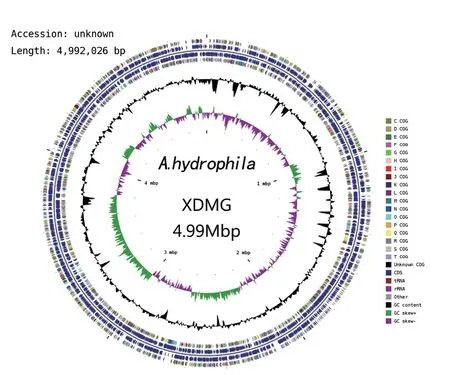
图1 嗜水气单胞菌XDMG全基因组的圆形图谱Fig. 1 Circular mapping of the whole genome of A. hydrophila XDMG
基于31个看家基因(DnaG,Frr,InfC,NusA,Pgk,PyrG,RplA,RplB,RplC,RplD,RplE,RplF,RplK,RplL,RplM,RplN,RplP,RplS,RplT,RpmA,RpoB,RpsB,RpsC,RpsE,RpsI,RpsJ,RpsK,RpsM,RpsS,SmpB,Tsf)利用MEGA 6.0软件对XDMG和19株在种属水平上最接近的菌株进行系统发育树分析(图2)。通过树状图发现,嗜水气单胞菌XDMG与国内株嗜水气单胞菌J-1和美国来源的嗜水气单胞菌AL09_71具有较近的亲缘关系。因此,本研究将嗜水气单胞菌XDMG基因组与其他4株亲缘关系的嗜水气单胞菌基因组的一般特征进行了比较分析(表2)。通过对比发现嗜水气单胞菌XDMG的rRNA与tRNA的数量明显低于其他4个菌株。
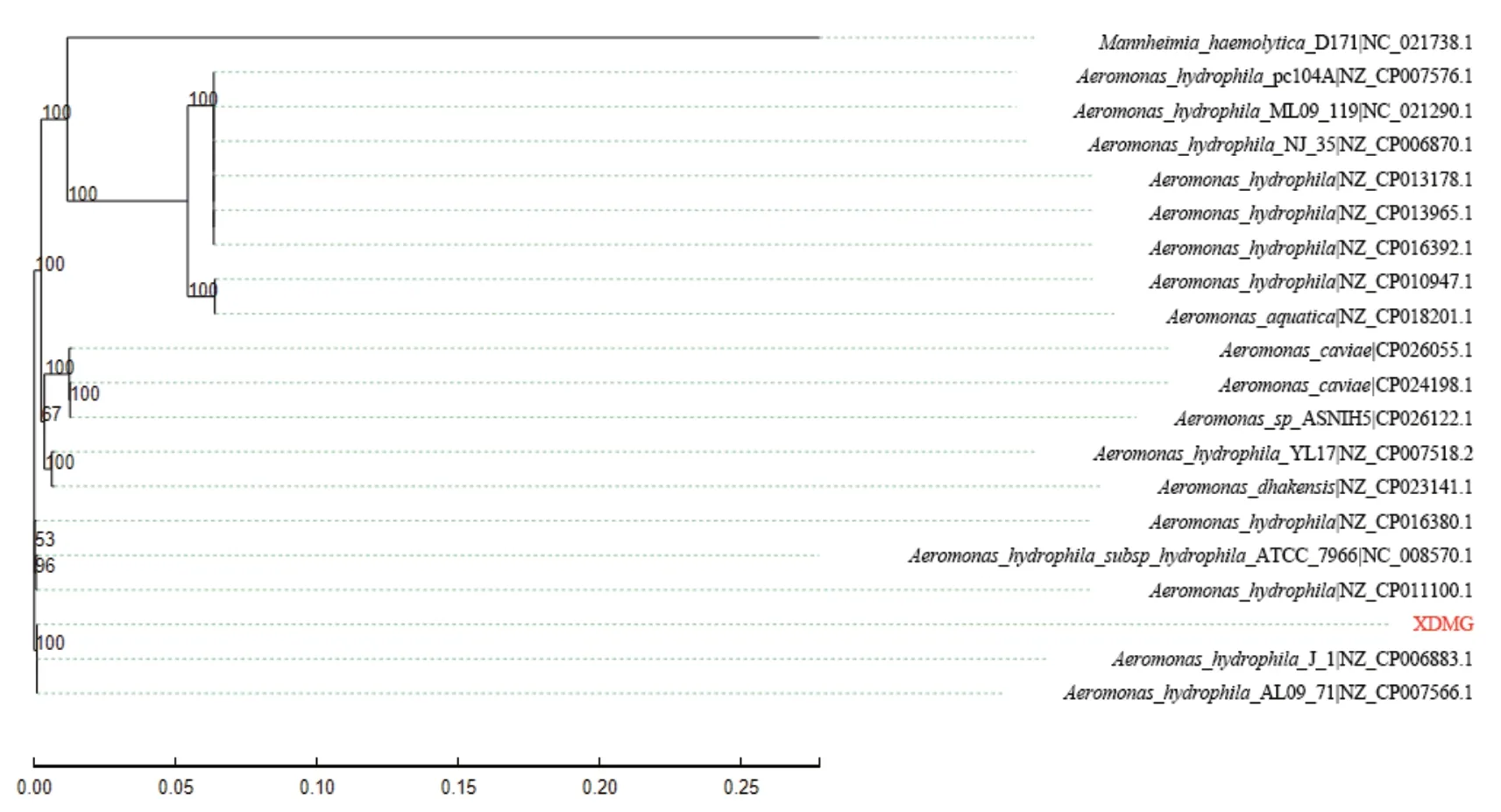
图2 基于31个看家基因序列构建的系统发育树Fig. 2 Phylogenetic tree constructed based on 31 housekeeping gene sequences
串联重复序列是基因调控网络的组成部分,在进化、遗传和变异中具有不同的功能作用,例如核酸碱基的错配修复和损伤修复,并且它在基因表达、染色体构建和转录调控等方面也发挥至关重要的作用。在嗜水气单胞菌XDMG中共有101条串联重复序列,重复序列的总长度占全基因组的0.5%。
2.3 基因功能注释
嗜水气单胞菌XDMG基因序列中预测到了4061个编码基因具有COG功能分类,有COG注释的基因占所有基因的82.29%。嗜水气单胞菌XDMG中编码序列COG功能注释(图3),基因注释结果共有21类,其中S(功能未知)的数量有1219个基因,数量最多占30.02%;A(RNA加工和修饰)与B(染色质结构和动力学)的数量最少,每种分别有1个基因;E(氨基酸的运输和代谢)有331个基因,占8.15%;K(转录)有274个基因,占6.75%;C(能源生产和转化)有256个基因,占6.30%;P(无机离子转运与代谢)有249个基因,占6.13%;T(信号转导机制)有231个基因,占5.69%;G(碳水化合物运输和代谢)有212个基因,占5.22%;M(细胞壁/膜/膜生物合成)有204个基因,占5.02%;J(翻译、核糖体结构和生物发生)有188个基因,占4.63%;L(复制、重组和修复)有186个基因,占4.58%;O(翻译后修饰、蛋白质周转、分子伴侣)有155个基因,占3.82%;H(辅酶转运和代谢)有122个基因,占3.00%;U(细胞内运输、分泌和囊泡运输)有102个基因,占2.51%;在COG功能注释中,有1219个编码基因的基因功能未知,说明在该基因组中还有许多基因功能需要去挖掘。
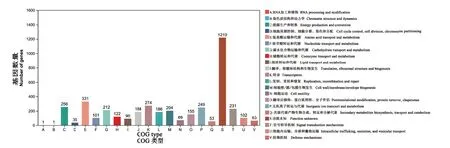
图3 嗜水气单胞菌XDMG在COG数据库的功能注释Fig. 3 Functional annotation of A. hydrophila XDMG in COG Database
嗜水气单胞菌XDMG编码蛋白的序列中有3228个编码序列在GO数据库中得到注释(图4)。占总编码序列的65.41%,基因差异性表达主要分布在3大类41个亚功能中:生物过程,包括19个亚功能;细胞成分,包括11个亚功能;分子功能,包括11个亚功能。有2574个编码序列被注释到生物过程类中,其中大部分参与代谢过程,细胞过程,单一生物过程。有1663个编码序列被注释到细胞组成类中,其中占比最多的是细胞、细胞部分、膜。有2662个编码序列被注释到分子功能类中,其中被注释到催化活性、结合、转运活性亚功能中编码序列最多。

图4 嗜水气单胞菌XDMG的GO注释分布Fig. 4 GO annotation distribution for A. hydrophila XDMG
嗜水气单胞菌XDMG编码蛋白的序列中有2663个编码序列在KEGG数据库中得到注释,分为6大类(图5),即1797个基因被注释到代谢;391个基因被注释到环境信息处理;300个基因被注释到细胞过程;231个基因被注释到遗传信息处理;166个基因被注释到人类疾病;65个基因被注释到生物系统,如图所示。通过对KEGG信号通路层次分析发现,代谢中的碳水化合物代谢包含基因最多,共391个基因;并发现有166个基因被注释到人类疾病,分别为:癌症概述(22个基因)、细菌传染病(31个基因)、心血管疾病(9个基因)、内分泌和代谢疾病(10个基因)、寄生虫传染病(4个基因)、神经退行性疾病(14个基因)、特定类型癌症(3个基因)、耐药性抗肿瘤(12个基因)、抗微生物药物(59个基因)、免疫疾病(2个基因)。
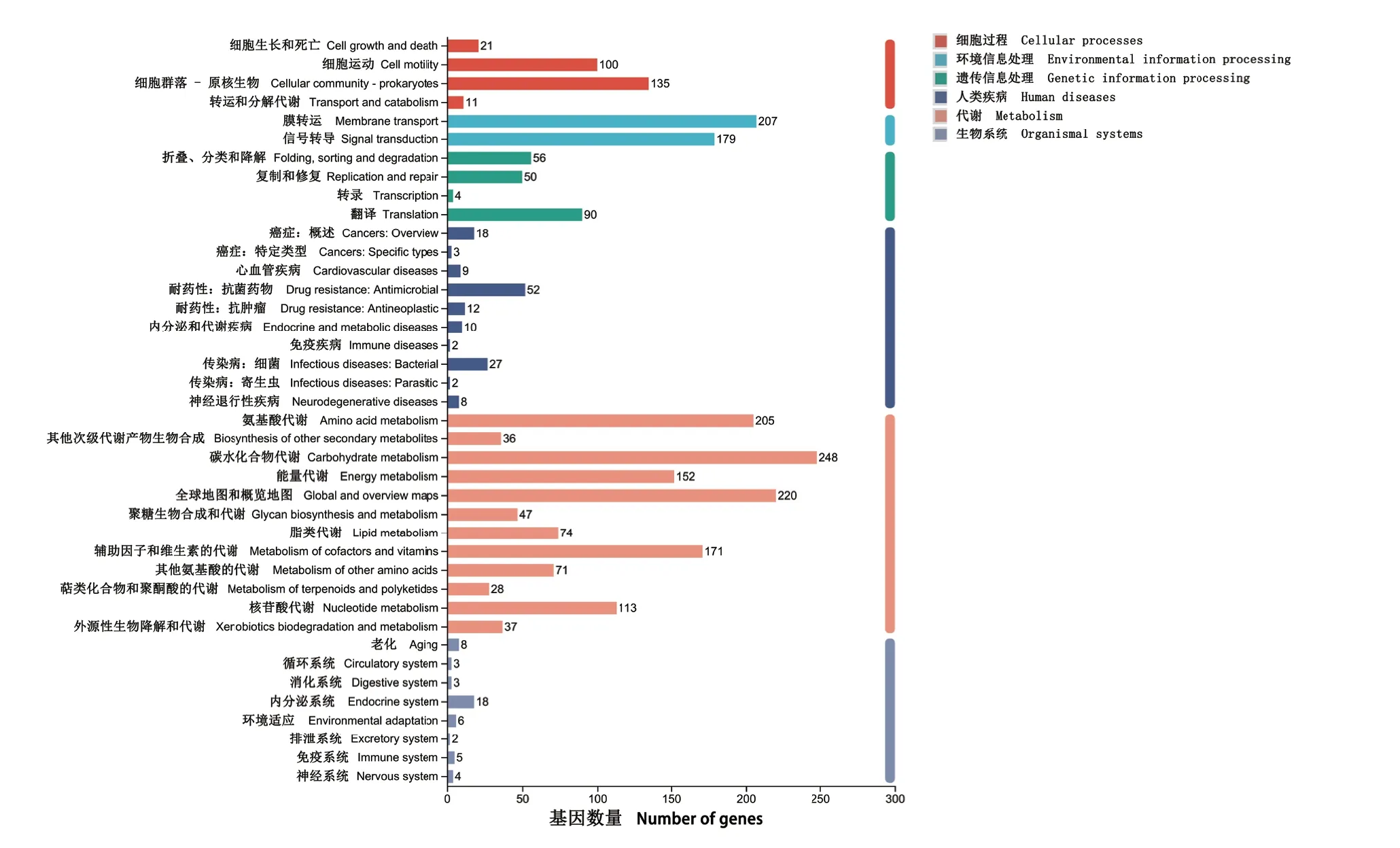
图5 嗜水气单胞菌XDMG的KEGG标注分布Fig. 5 KEGG labeling distribution for A. hydrophila XDMG
2.4 基因岛
通过与数据库对比,在嗜水气单胞菌XDMG中共发现14个基因岛(表3),长度在10-15 kb之间的基因岛数量最多,占35.71%,最大的基因岛长度为25.03 kb。全部基因岛共涉及255个基因,具有明确生物学意义的基因有113个基因,其中涉及基因最多的基因岛,共涉及34个基因,具有明确生物学意义的基因有2个基因,涉及明确生物学意义的基因最多的基因岛,共涉及25个基因,其中有16个具有明确生物学意义,具体情况如表。同时这些基因涉及的多种功能,与蛋白质编码与转运、蛋白质修复、酶合成、DNA修复、能量系统等均有密切的联系。
2.5 毒力基因及耐药基因分析
在嗜水气单胞菌XDMG中共鉴定出527个毒力基因(表4),其中有256个毒力基因有明确的分类,分别为防御性毒力因子(34个基因)、攻击性毒力因子(158个基因)、非特异性毒力因子(54个基因)、毒力相关基因的调控(10个基因)。
在嗜水气单胞菌XDMG中存在279个耐药基因,分为35类(表5),其中外排泵赋予抗生素耐药性的基因最多,包含179个基因,酚类耐药性基因、大环内酯类耐药性基因、利奈唑胺耐药基因、林可酰胺耐药基因、呋喃妥因耐药基因、参与抗生素自身耐药的基因最少,都只有1个相关基因。
在5株嗜水气单胞菌中都存在II型分泌系统(表6)。而III型分泌系统仅存在于嗜水气单胞菌J-1中,在其他4株嗜水气单胞菌中都不存在。在嗜水气单胞菌XDMG中发现多个T6SS的效应因子,包括一个脯氨酸-丙氨酸-丙氨酸-精氨酸重复序列(PAAR,gene4949),两个溶血素共调控蛋白(Hcp,gene3839,gene4717),3种缬氨酸-甘氨酸重复序列G(VgrG,gene3035,gene3409,gene4971)。在嗜水气单胞菌AL09-71中只存在VgrG,缺失PAAR与Hcp。
在嗜水气单胞菌XDMG中存在3个极性鞭毛基因簇(gene0132-gene0147、gene0124-gene0126、gene2719-gene2744),5个IV型菌毛基因(gene0576-gene0580、gene2243-gene2246、gene2293-gene2497、gene2478、gene3398)与一个I型菌毛基因(gene1946-gene1951),这些基因也存在于其他4株菌中,但是在这5株菌中均不存在与侧鞭毛相关的基因。
在嗜水气单胞菌XDMG中发现细胞兴奋性肠毒素基因(Alt,gene4667)、细胞毒素基因(Ast,gene4528)、气溶素(Aer,gene2040)、溶血素(HlyA,gene3120)、耐热性的溶血素(gene0601)和溶血素III(gene4993),毒素基因均可在其他4株嗜水气单胞菌中找到相关基因。在嗜水气单胞菌XDMG中有大量编码胞外酶的基因,并且这些基因具有明显保守性。此外,嗜水气单胞菌XDMG菌株中检测到了RTX毒素的相关基因RtxA(gene2749)与RtxC(gene2750),在J-1、AL09_71、AH10和ATCC_7966中同样存在RTX毒素激活基因RtxA和RtxC。
在嗜水气单胞菌XDMG基因中参与合成Amonabactin的基因位于一个簇(gene1421-gene1427)内,从而使得嗜水气单胞菌XDMG在感染宿主时能够获取足够的铁元素用于生长所需。在嗜水气单胞菌XDMG和其他4株嗜水气单胞菌的基因中都存在铁铬铁摄取(gene3527-gene3530)、铁摄取调节剂(gene3139)。
在嗜水气单胞菌XDMG的染色体中存在16个β-内酰胺抗性基因,包括AmpC(gene0493)、AmpS(gene4846)和CphA(gene1713),并且存在编码多黏菌素B(ArnA,gene3680)、耐有机氢过氧化物(gene2546-gene2549)、多药耐药蛋白(DsbA,gene2156)、肽类抗生素耐药蛋白(gene3921)和磷霉素抗性蛋白(gene2177、gene2683、gene3268)的基因。在XDMG中有179个外排泵赋予耐药性基因,其中包括Smr(gene2646)、Mfs(gene1077、gene1529、gene3362、gene4896)、Dmt(gene1039)、Rnd(gene0290、gene4352)与ABC超家族(gene4866)。
嗜水气单胞菌中群体感应(quorum sensing,QS)系统分为3类,分别为自体诱导物信号分子-1(AI-1)、自体诱导物信号分子-2(AI-2)与自体诱导物信号分子-3(AI-3)。在嗜水气单胞菌XDMG基因组中发现编码AhyR蛋白(gene1912)与AhyI蛋白(gene1913)是AI-1 QS系统的调控基因。在XDMG基因组存在编码MetK(gene0484)、SAM甲基转移酶(gene1399)、LuxS(gene1756)、MetH(gene2048)、MtnN(gene3349)及MetE(gene3532)的基因,LuxS基因是AI-2 QS系统中的重要基因。同时在其基因组中也存在AI-3 QS系统的调控基因QseB(gene0610)与QseC(gene0611),通过对嗜水气单胞菌XDMG基因组的分析,在嗜水气单胞菌XDMG基因组中发现AI-1、AI-2和AI-3 三种QS系统。
为了更加清楚5株嗜水气单胞菌在致病因素方面的差异,以嗜水气单胞菌XDMG作为参考菌株,与嗜水气单胞菌J-1、嗜水气单胞菌AL09_71、嗜水气单胞菌AH10和嗜水气单胞菌ATCC_7966进行核苷酸水平与氨基酸水平的序列对比,主要比对Hly、Aer、Ast、Alt、AhyI和AhyR六个致病相关基因。结果表明嗜水气单胞菌XDMG与4株嗜水气单胞菌在核苷酸水平上均高度保守(表7)。嗜水气单胞菌XDMG与嗜水气单胞菌AL09_71在6个致病相关基因序列上同源性均达到了100%;与国内株嗜水气单胞菌 J-1相比同源性均在99%以上;与嗜水气单胞菌ATCC_7966和嗜水气单胞菌AH10相比同源性均在97%以上,存在缺失、插入和替换现象。在氨基酸水平上,以嗜水气单胞菌XDMG作为参考菌株,5株嗜水气单胞菌在各致病相关基因编码的氨基酸序列上也高度保守,5株嗜水气单胞菌Hly与AhyI编码的氨基酸序列的保守程度均达到100%;嗜水气单胞菌AL09_71与嗜水气单胞菌XDMG在Hly、Aer、Ast、Alt、AhyI和AhyR编码的氨基酸序列上均具有100%的保守性;嗜水气单胞菌 J-1 除基因AhyR外,Hly、Aer、Ast、Alt和AhyI编码的氨基酸序列与嗜水气单胞菌XDMG各基因编码的氨基酸序列均具有99%以上的保守性;嗜水气单胞菌ATCC_7966和嗜水气单胞菌AH10与嗜水气单胞菌XDMG在Hly、Aer、Ast、Alt、AhyI和AhyR编码的氨基酸序列上保守性均高于97%(表8)。
在嗜水气单胞菌XDMG中发现了12个外膜蛋白,分别是3个OmpA(gene2333、gene2864、gene3980)、2个OmpAI(gene2059、gene2862)、OmpAII(gene2863)、OmpC(gene4897)、OmpK(gene3024)、OmpN(gene0081)、OmpU(gene2094)、OmpW(gene4572)和Omp26La(gene3292)。为了更加清楚5种嗜水气单胞菌在外膜蛋白因素方面的差异,以嗜水气单胞菌XDMG作为参考菌株,以上述12个外膜蛋白与嗜水气单胞菌J-1、嗜水气单胞菌AL09_71、嗜水气单胞菌AH10和嗜水气单胞菌ATCC_7966进行核苷酸水平与氨基酸水平的序列对比。结果表明嗜水气单胞菌XDMG与嗜水气单胞菌AL09_71、嗜水气单胞菌ATCC_7966和国内株嗜水气单胞菌J-1和嗜水气单胞菌AH10无论是在核苷酸水平还是氨基酸水平序列均高度保守。嗜水气单胞菌XDMG与嗜水气单胞菌AL09_71在核苷酸水平和氨基酸水平上均达到了100%(表9)。国内株嗜水气单胞菌J-1 除基因OmpN外,其他外膜蛋白基因序列及各外膜蛋白基因编码的氨基酸序列与嗜水气单胞菌XDMG外膜蛋白基因序列和各外膜蛋白基因编码的氨基酸序列保守性均达到100%(表10)。嗜水气单胞菌AH10与嗜水气单胞菌XDMG相比缺少OmpAI(gene2059)和Omp26La(gene3292),嗜水气单胞菌ATCC_7966与嗜水气单胞菌XDMG相比缺少OmpC(gene4897)和OmpAI(gene2059),除此之外,其他外膜蛋白基因序列与XDMG的外膜蛋白基因序列同源性均在94%以上,且嗜水气单胞菌ATCC_7966和嗜水气单胞菌AH10各外膜蛋白基因编码的氨基酸序列与XDMG各外膜蛋白基因编码的氨基酸序列保守程度均高于94%。
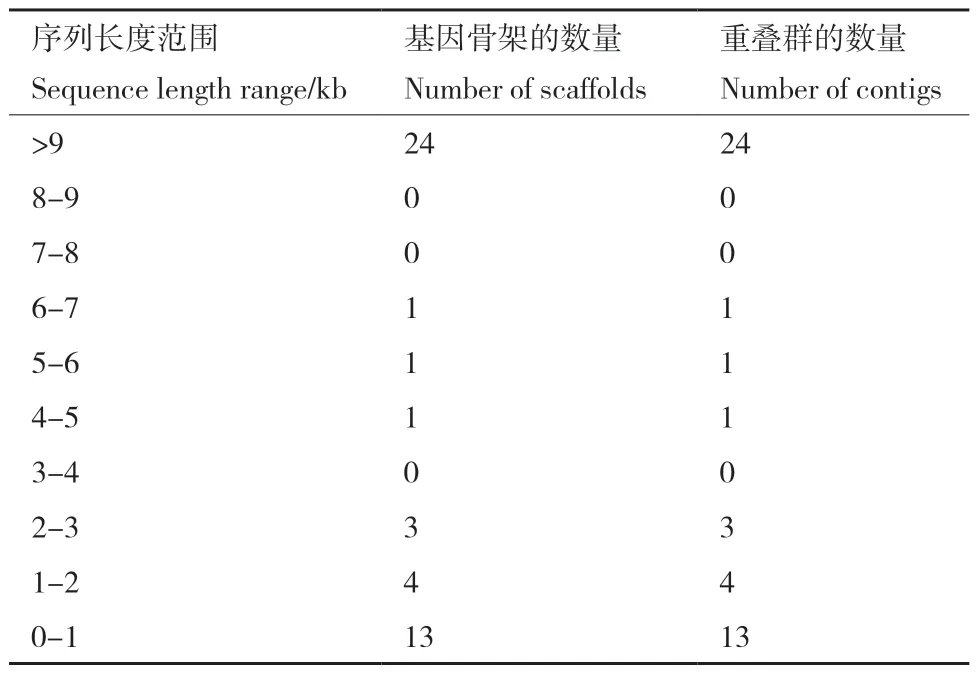
表1 嗜水气单胞菌 XDMG基因组骨架数据Table 1 Aeromonas hydrophila XDMG genome backbone data
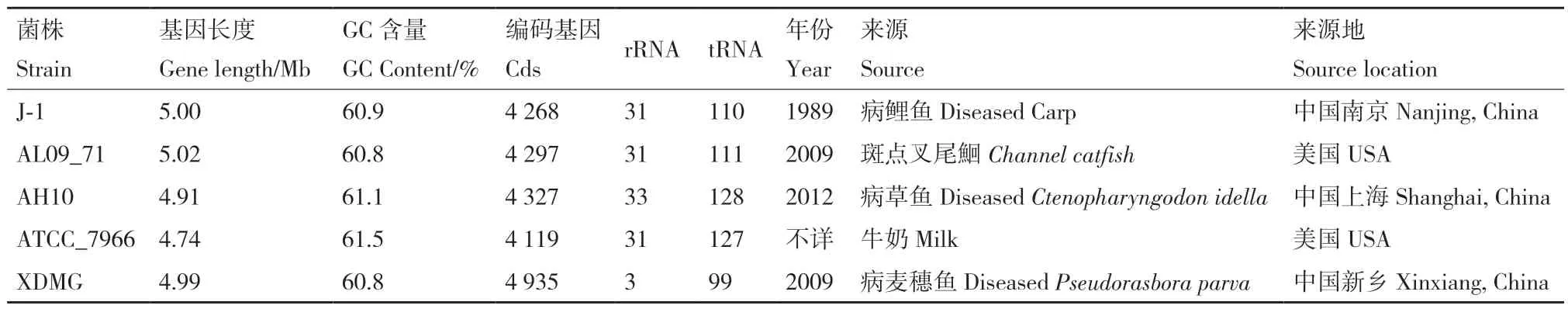
表2 五株嗜水气单胞菌基因组的一般特征Table 2 General characteristics of the five Aeromonas hydrophila genomes

表3 嗜水气单胞菌 XDMG的基因组岛长度分布Table 3 Distribution of genomic island lengths in A. hydrophila XDMG
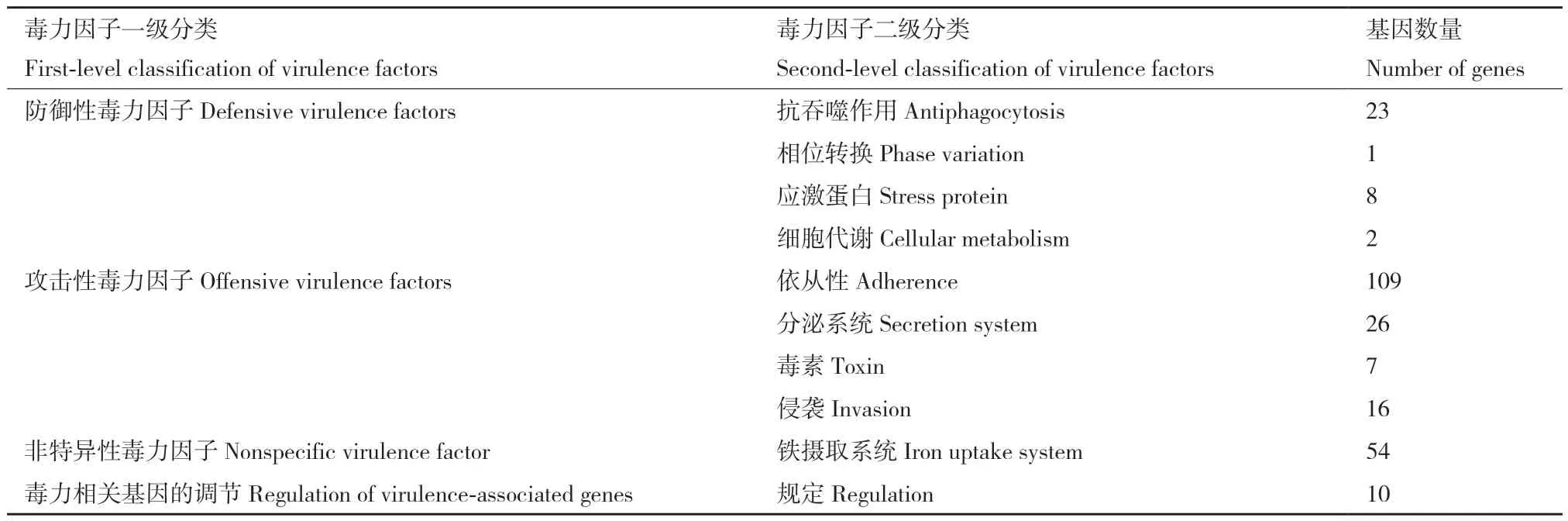
表4 嗜水气单胞菌 XDMG的毒力基因的分类情况Table 4 Classification of virulence genes in A. hydrophila XDMG

表5 嗜水气单胞菌 XDMG的耐药基因的分类情况Table 5 Classification of resistance genes in A. hydrophila XDMG

表6 五株嗜水气单胞菌毒力基因的分布情况Table 6 Distribution of virulence genes in 5 strains of A. hydrophila
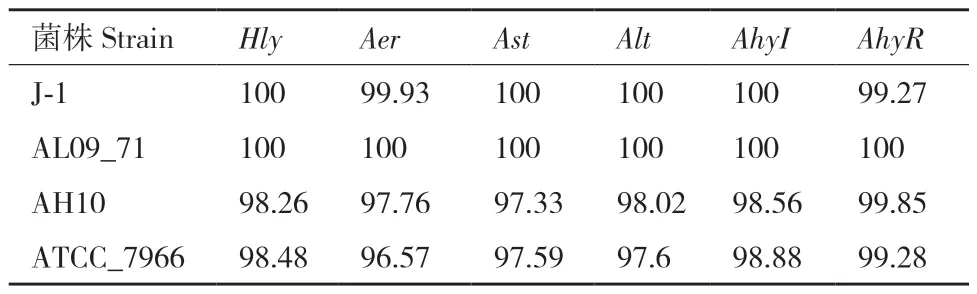
表7 以嗜水气单胞菌XDMG为参考菌株各菌株毒力基因序列的同源性Table 7 Homology of virulence gene sequence of each stra-in using A. hydrophila XDMG as a reference strain/%
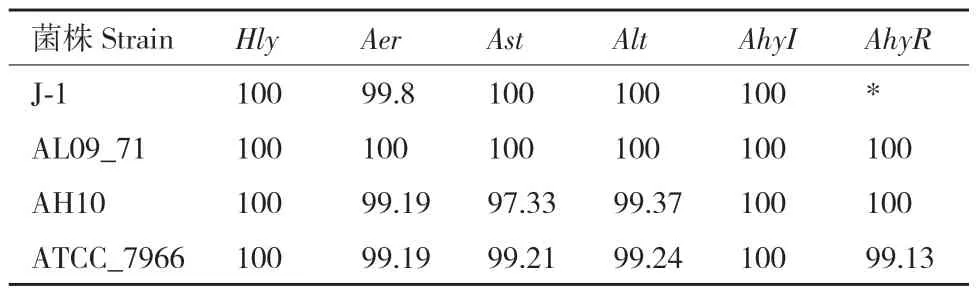
表8 以嗜水气单胞菌XDMG为参考菌株各菌株毒力基因编码氨基酸序列的保守性Table 8 Conservation of amino acid sequences encoded by virulence genes of each strain using A. hydrophila XDMG as a reference strain/%
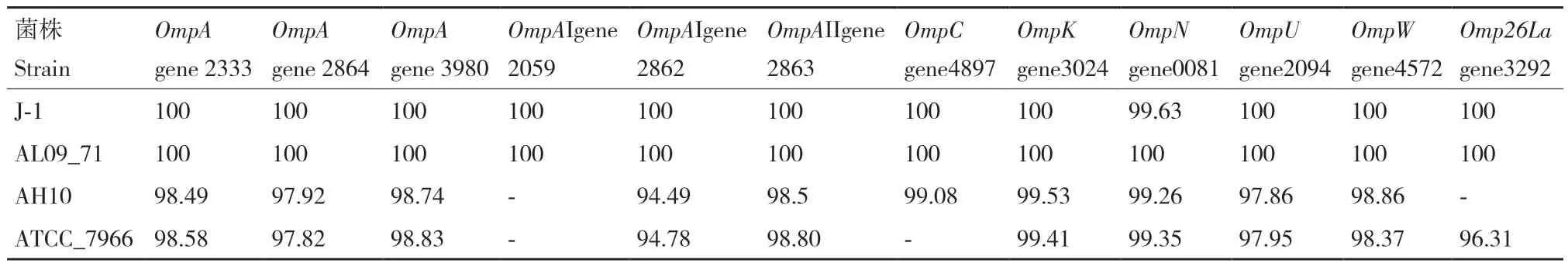
表9 以嗜水气单胞菌XDMG为参考菌株各菌株外膜蛋白基因序列的同源性Table 9 Homology of outer membrane protein gene sequences of each strain using A. hydrophila XDMG as a reference strain/%
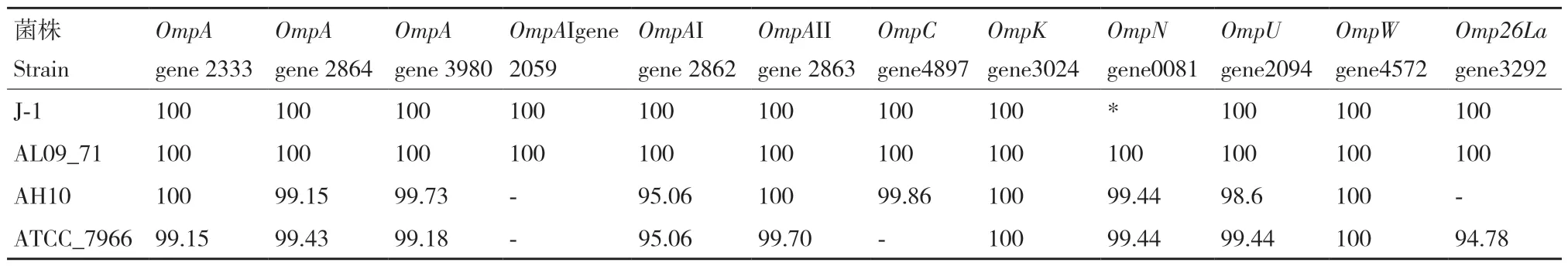
表10 以嗜水气单胞菌XDMG为参考菌株各菌株外膜蛋白基因编码氨基酸序列的保守性Table 10 Conservation of amino acid sequence encoded by outer membrane protein gene of each strain using A. hydrophila XDMG as a reference strain/%
本研究对5株嗜水气单胞菌的O-抗原基因簇进行了比较分析发现,嗜水气单胞菌XDMG与AL09_71的O-抗原血清型为OX 1型,嗜水气单胞菌AH10的O-抗原血清型为OX 5型,嗜水气单胞菌ATCC_7966的O-抗原血清型为OX 6型,嗜水气单胞菌J-1的O-抗原血清型为O 33型(图6)。每种类型的O-抗原基因簇在基因的组成及结构上都存在差距,在嗜水气单胞菌XDMG、AL09_71与ATCC_7966的O-抗原基因簇的组成中有合成GDP-甘露糖所必需的ManA、ManB与ManC基因,在嗜水气单胞菌J-1和AH10的O-抗原基因簇中不存在这3个基因。此外嗜水气单胞菌XDMG与AL09_71的O-抗原基因簇中含有合成Dtdp-糖-3-乙酰氨基-3,6-二脱氧- D-半乳糖所必需的FdtA、FdtB与FdtC基因。此外每种类型的O-抗原基因簇中都含有RmlA、RmlB、RmLC与RmlD基因。以嗜水气单胞菌XDMG作为参考菌株,对RmlA、RmlB、RmLC与RmlD基因的核苷酸序列及氨基酸序列进行对比发现,嗜水气单胞菌XDMG与嗜水气单胞菌AL09_71的4个基因无论是在核酸水平还是在氨基酸水平其同源性达到100%,嗜水气单胞菌XDMG与嗜水气单胞菌J-1、AH10和ATCC_7966的RmlB基因在核苷酸水平与氨基酸水平上同源性达到了80%以上,其他基因无论是在核苷酸水平上还是在氨基酸水平上其同源性均不足65%(表11-12)。
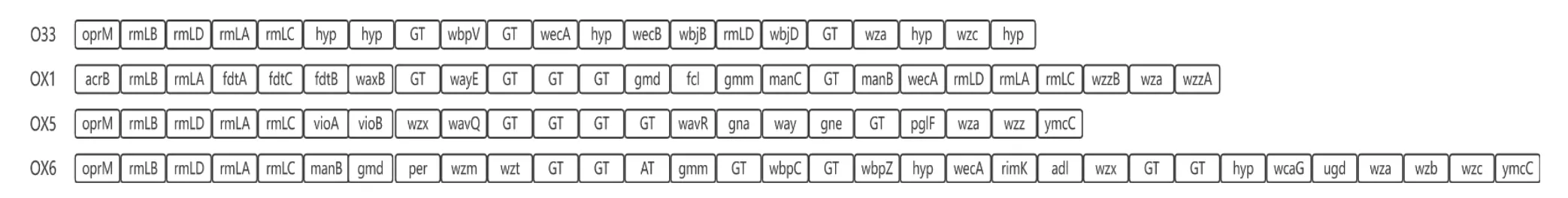
图6 本研究嗜水气单胞菌的O-抗原基因簇Fig. 6 O-antigen gene cluster of A. hydrophila in this study
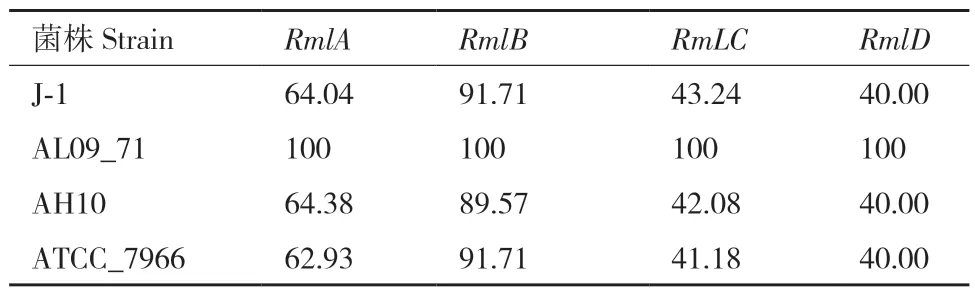
表12 以嗜水气单胞菌XDMG为参考菌株各菌株O-抗原基因簇相同基因编码氨基酸序列的保守性Table 12 Conservation of amino acid sequences encoded by the same gene of O-antigen gene cluster of each strain using A. hydrophila XDMG as the reference strain/%
3 讨论
随着测序技术的快速发展,全基因组测序技术已经广泛应用于动植物病原菌的鉴定与分析,可通过基因组的比较分析了解病原菌的遗传进化关系、致病机制等,为病原菌的鉴别分类、耐药性的检测、防病策略的制定以及疫苗开发提供有价值的参考依据[32-33]。为了了解嗜水气单胞菌XDMG的基因组结构、功能及其进化关系,本研究对其进行全基因测序及基因组的比较分析。
嗜水气单胞菌XDMG测序结果显示,其基因组大小为4.99 Mb,GC含量为60.84%,该基因组包含了4935个编码基因,有4061个编码基因序列在COG数据库中得到功能注释,基因功能注释占比最多是S(基因功能未知),说明在该基因组中还有许多基因功能需要去挖掘。通过GO数据库进行注释表明,嗜水气单胞菌XDMG的蛋白功能主要集中在生物过程的代谢过程与分子功能的催化活性中。KEGG注释表明,嗜水气单胞菌XDMG拥有丰富且完整的代谢途径,可以使生物体自身不断进行物质与能量交换,维持自身活动必需物质和对外界环境及时作出反应,增强病原菌的环境适应性。
如今已经清楚嗜水气单胞菌可以分泌大量可降解和产毒的胞外蛋白,并且嗜水气单胞菌具有多种分泌机制,包括I型、II型、III型和VI型分泌系统[34-37]。其中II型分泌系统介导宿主黏附和侵袭、宿主细胞破坏和组织坏死等一系列致病过程[38]。VI型分泌系统[39]是一种运输系统,将蛋白质运输到胞外,与其他毒性决定因素密切相关,同时VI型分泌系统自身也是嗜水气单胞菌重要的毒力因子之一。Ⅱ型与VI型分泌系统嗜水气单胞菌的致病性中发挥了重要的作用[40],本研究发现在Ⅱ型分泌系统存在于5株嗜水气单胞菌中,VI型分泌系统存在于4株嗜水气单胞菌,在嗜水气单胞菌AL09-71中只存在VI型分泌系统效应因子的VgrG,缺失PAAR与Hcp。5株嗜水气单胞菌中均不存在III型分泌系统。虽然在嗜水气单胞菌XDMG中不存在III型分泌系统,但是存在与AI-1(AhyI,AhyR)、AI-2(LuxS)和AI-3(QseB、QseC)QS系统相关的基因,并且QseB与QseC基因序列中存在4 bp的重复序列ATGA与Khajanchi等[41]发现的一样,表明嗜水气单胞菌XDMG中可能存在AI-1、AI-2和AI-3三种QS系统。群体感应是细菌一种通信系统,涉及分泌和检测特定信号分子,以调节基因表达和多种生理变化[42-44],并且QS系统在嗜水气单胞菌致病性中起着重要的作用[42]。嗜水气单胞菌中存在多种毒力因子,每种因子均发挥着不同的作用。鞭毛与菌毛是重要的黏附因子,它们在嗜水气单胞菌入侵与定植宿主的过程中起着尤为重要作用[45-46]。根据现有的报道表明,分泌酶和毒素与嗜水气单胞菌致病的主要因素之一[39,47]。铁元素也是嗜水气单胞菌在宿主中生存所必需的重要元素之一,它对嗜水气单胞菌的毒力具有重要的作用[48-49]。Amonabactin是众所周知的酚类铁载体,并广泛存在于气单胞菌中[50],它能使嗜水气单胞菌XDMG在感染宿主时能够获取足够的铁元素用于生长所需。本研究对5株嗜水气单胞菌的毒力因子进行分析发现,嗜水气单胞菌的毒力基因具有保守性和一致性,包括分泌系统、鞭毛、黏附因子、毒素、酶、群体感应和铁获取等基因。Jin等[39]对嗜水气单胞菌HX-3进行全基因组测序,在毒力因子分析结果中同样发现嗜水气单胞菌的毒力基因具有保守性和一致性,并且表明II型与VI型分泌系统在嗜水气单胞菌的致病性中发挥着重要作用,III型分泌系统对嗜水气单胞菌HX-3和其他菌株的毒力贡献较小。同时嗜水气单胞菌的耐药性也是一个不得忽视的重点,嗜水气单胞菌的耐药性阻碍疾病的治疗,对人类健康的具有潜在威胁[51-52]。本研究对5株嗜水气单胞菌的耐药基因进行分析发现,嗜水气单胞菌的耐药基因同样具有一致性。
本研究发现在嗜水气单胞菌XDMG基因组中存在细胞兴奋性肠毒素基因(Alt)、细胞毒素基因(Ast)、细胞毒性肠毒素基因(Act)、溶血素(HlyA)、耐热性的溶血素和溶血素,这些基因与嗜水气单胞菌的致病性有密切的关系[12]。对比5株嗜水气单胞菌Alt、Ast、Hly、Aer、AhyI与AhyR基因的核酸与氨基酸序列发现,无论是基因序列还是氨基酸序列均具有较高的保守性,表明嗜水气单胞菌的主要致病基因在遗传过程中发生变异的概率比较小。在张国亮等[12]通过对嗜水气单胞菌AH10进行全基因组测序及比较分析中也验证了这一点。
外膜蛋白(outer membrane protein,Omp)也是嗜水气单胞菌重要的毒力因子之一,它们在黏附和逃避宿主防御机制方面发挥着重要作用[53-54]。此外,外膜蛋白可在宿主受到相应毒力菌株的攻击时提供保护[55]。由于其数量丰富,以及细菌外膜上的暴露区域,宿主很容易将其识别为外来颗粒,并导致产生免疫反应[53]。在众多研究中,外膜蛋白一直被认为是潜在的候选疫苗[56-58],其具有较好免疫原性。例如Yadav等[56]将重组的嗜水气单胞菌外膜蛋白OmpC免疫露斯塔野鲮发现, OmpC免疫鱼的血清中铜蓝蛋白水平、髓过氧化物酶和抗蛋白酶活性在瞬时和瞬时方式中显著增加,并且凝集和溶血活性滴度增加,相对存活率为66.02%。本研究对嗜水气单胞菌的外膜蛋白进行比较分析发现,嗜水气单胞菌J-1外膜蛋白OmpN(gene0081)基因不表达蛋白;嗜水气单胞菌AH-10缺失了外膜蛋白OmpAI(gene 2059)与 Omp26La(gene3292);嗜水气单胞菌ATCC_7966缺失了外膜蛋白OmpAI(gene 2059)与OmpC(gene4897),同时也发现其余外膜蛋白无论是基因序列还是氨基酸序列均具有较高的保守性,表明嗜水气单胞菌XDMG具有更加全面的外膜蛋白,采用嗜水气单胞菌XDMG的外膜蛋白研制基因工程疫苗,其应用范围更加广泛。
O-抗原对宿主-病原体相互作用极其重要,因为它们在黏附、细胞识别和生物膜形成中起着关键作用[59-60]。通过对5株嗜水气单胞菌的O-抗原基因簇比较分析发现,在嗜水气单胞菌XDMG、AL09_71与 ATCC_7966的O-抗原基因簇的组成中有合成GDP-甘露糖所必需的ManA、ManB与ManC基因,在嗜水气单胞菌J-1和AH10的O-抗原基因簇中不存在这3个基因。此外嗜水气单胞菌XDMG与AL09_71的O-抗原基因簇中含有合成Dtdp-糖-3-乙酰氨基- 3,6-二脱氧- D-半乳糖所必需的FdtA、FdtB与FdtC基因。虽然每种类型的O-抗原基因簇中都含有RmlA、RmlB、RmLC与RmlD基因,但是其同源性不高。O-抗原基因簇部分的可变化学结构决定了单个细菌菌株的免疫特异性,O-抗原基因簇在结构上的差异十分重要,这有助于了解嗜水气单胞菌的免疫特异性[61]。
4 结论
通过对嗜水气单胞菌XDMG基因组进行较为全面的基因组注释,有利于了解嗜水气单胞菌基因组的结构、功能及进化关系。对嗜水气单胞菌XDMG与4株嗜水气单胞菌进行基因组的比较分析,有利于了解不同区域的嗜水气单胞菌在毒力因子、外膜蛋白与O-抗原基因簇等方面的差异,为寻找出不同区域的嗜水气单胞菌共同的免疫保护抗原功能基因奠定基础,也为下一步研制嗜水气单胞菌XDMG基因工程疫苗提供理论支持。
猜你喜欢
当代水产(2022年3期)2022-04-26 14:27:36
海洋石油(2021年3期)2021-11-05 07:42:26
西北农林科技大学学报(自然科学版)(2019年8期)2019-07-17 02:43:32
哈尔滨医药(2016年3期)2016-12-01 03:58:34
癌变·畸变·突变(2015年4期)2015-02-27 06:15:17
现代检验医学杂志(2015年1期)2015-02-06 01:59:13
遗传(2015年5期)2015-02-04 03:06:55
海洋科学(2014年12期)2014-12-15 03:35:00
现代检验医学杂志(2014年4期)2014-02-02 02:44:54
食品科学(2013年15期)2013-03-11 18:25:36
