
玉米矮秆基因与矮秆育种研究
2023-09-01 10:41:28王天依王荣焕王夏青张如养徐瑞斌焦炎炎孙轩王继东宋伟赵久然
生物技术通报 2023年8期
王天依 王荣焕 王夏青 张如养 徐瑞斌 焦炎炎 孙轩 王继东 宋伟 赵久然
(北京市农林科学院玉米研究所 玉米DNA指纹及分子育种北京市重点实验室,北京 100097)
玉米(Zea mays L.)为一年生禾本科作物,起源于现在的墨西哥等中南美洲地区,与水稻、小麦并称为世界“三大主粮”,在我国约有500年的栽培史。玉米籽粒中含有丰富的淀粉、蛋白质、油分等营养物质,是重要的粮食作物。此外,玉米含有大量的碳水化合物,有效能值高,且富含维生素、不饱和脂肪酸和氨基酸,有着“饲料之王”的美誉。除食用和饲用外,玉米可被加工成黏合剂、工业酒精和燃料乙醇等工业品,亦是重要的化工原料[1]。
我国对玉米的需求呈持续增长态势,在有限的耕地面积下提高玉米产量尤为重要。在过去的90年间,美国玉米的单株产量并未明显增加,亩产量却提升7倍有余,其主要归功于耐密植品种的选育及推广[2]。然而高密度种植可能会加剧植株间光、水和养分等资源的竞争,造成植株徒长,并降低植株对生物和非生物胁迫的抵御能力,增加倒伏风险[3-4]。株高是制约耐密育种的关键因素。适度降低株高,既能增加单位面积的种植株数、提高单位面积总产量,又能有效避免倒伏的发生,对增强植株的耐密性和抗倒伏性极为重要[5]。株高性状的半矮化遗传改良已经成为玉米育种的重要方向之一[6]。本文详述了调控玉米矮秆相关基因及其分子机制,并对玉米矮秆品种选育进行概述。对现代生物技术在矮秆品种遗传改良中的发展前景加以展望,以期为玉米矮秆遗传育种提供参考。
1 玉米矮秆基因的克隆
1.1 首个玉米矮秆基因
玉米株高通常指从地面到植株最顶端的距离,在一些研究中,玉米株高被定义为植株地上部到旗叶基部的距离,以排除雄穗变异对株高的影响[7]。玉米株高由节间长度和节间数目共同决定,一般来说节间数目越少,节间长度越短,株高越矮。因此在一定生育期内,控制节间数目或节间伸长可以有效降低株高[8]。
1935年,Emerson发现了第一个玉米矮秆基因Brachytic2(Br2),Br2的发现为玉米矮秆、半矮秆育种奠定了基础[9]。br2突变体茎秆细胞长度较野生型相比减少40%-50%,使得其节间缩短,尤其是果穗以下节间缩短更为显著,株高降低约50%。与茎秆长度相反,茎秆的周长明显增加,br2突变体的茎秆表皮下方较野生型相比多形成了10余层薄壁细胞,极大增强了br2的茎秆强度。从农艺学角度,br2的表型是极具高产潜力的:其上疏下密的株型有助于通风透光,发达的根系使植株对水肥有着极强的耐性,而它最关键的优势则是对茎秆强度的协同效应,是公认的颇具商业价值的株高改良基因[10]。目前已知的矮秆基因有60余个,其中大部分已被克隆(表1)。
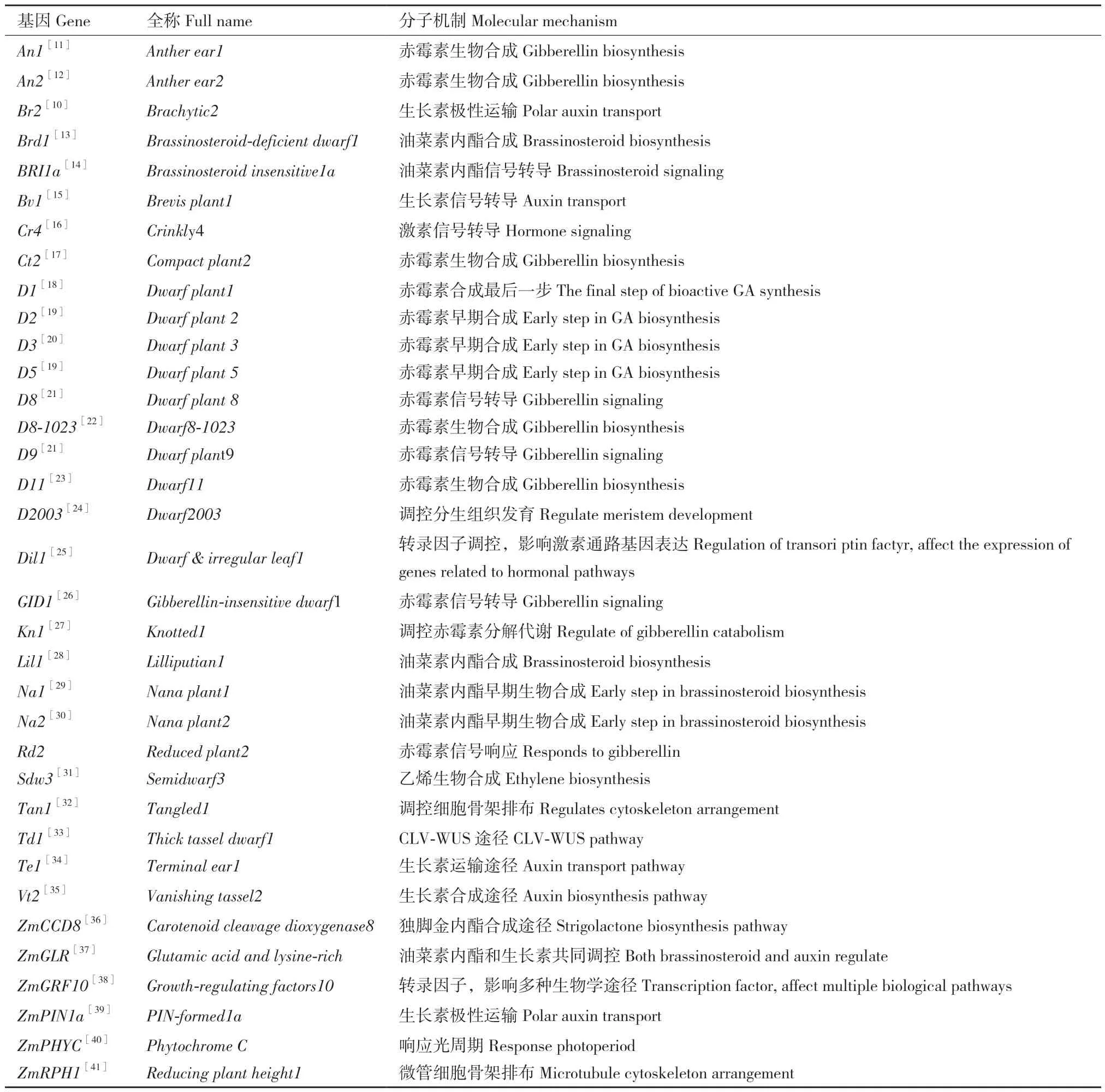
表1 已克隆的玉米矮秆基因Table1 Cloned genes related to dwarf maize
1.2 玉米株高数量性状位点的研究
一般而言,单基因调控的矮秆突变体对其他农艺性状和产量的影响较为剧烈,数量性状位点对株高和产量的影响较为温和,在遗传改良中更具有应用价值。随着定位技术的不断发展,研究者也开展了许多与株高相关的遗传研究。
Peiffer等[7]在13个环境下,对包括NAM群体在内的7300多份玉米自交系的株高、穗位高进行关联分析发现,株高的遗传力较高,达到90%,株高由强烈的遗传因素决定,但仍然微效多基因控制。Ku等[42]将4个重组自交系(RIL)群体在两个不同的密度下种植,通过分析在高密度条件下鉴定的株高QTL,发现部分QTL成簇存在,推测株高可能受一些具有加性效应的共有变异决定。另外,Fernandez等[43]研究表明株高与开花期、叶长、叶宽、茎节构造、小穗长度等性状相关,其中株高与开花期紧密相关,即晚花植物的株高较高。
目前,随着高通量测序技术、统计分析方法和遥感技术的快速发展,株高QTL的检出效率也随之增加。Yin等[44]将全基因组关联分析与高分辨率连锁作图法相结合,通过NAM群体对玉米株高、穗位高相关QTL进行定位,共鉴定出105个SNPs和22个QTLs与株高和穗位高显著相关。Sa等[45]利用Illumina Maize-SNP50芯片对玉米Mo17/KW7的RIL群体进行基因分型,检测到3个与株高相关的QTLs,表型变异范围为在16.01%-19.85%之间。Zhang等[46]发明了一种新的统计分析方法——数量性状基因测序(QTG-seq)来对数量性状进行定位。即通过QTL性状分离、极端表型混池测序技术以及关联分析新方法smoothLOD相结合,快速鉴定株高的候选基因[46]。Wang等[47]采用无人机遥感技术,对玉米自然群体不同发育时期的株高性状进行全基因组关联分析,鉴定到涉及不同发育阶段株高的QTL,为玉米株高的高通量研究提供借鉴。
Gramene数据库资料显示(https://archive.gramene.org/qtl),截至今日,已确定的调控玉米株高的quantitative trait locus(QTLs)共计219个,它们分布在玉米的10条染色体上,但在自然界中,或许仍有很多QTLs位点尚未得到鉴定,可能的原因有以下4方面:(1)株高相关QTLs位点的检测效率会受到群体结构的影响;(2)QTLs表型受遗传背景的影响;(3)对于具有拮抗多效性的基因,在自然群体中难以检测到,可以利用突变材料进行定位;(4)QTLs定位受环境或受基因型与环境互作的影响[48]。
2 矮秆玉米的分子调控机制
株高主要受植物激素调节,研究发现,生长素、油菜素内酯、赤霉素、乙烯、独脚金内酯等激素的合成、转运以及信号转导相关基因突变,均可影响玉米株高。另外,有部分基因则通过改变微管骨架排布或影响成花转变来调控玉米株高。
2.1 激素途径
2.1.1 生长素对玉米株高的调控 生长素(auxin,IAA)是发现最早的一种植物激素,它影响细胞分裂分化、器官生长、维管束分化、顶端优势等多个过程,对植物生长发育具有广泛的调节作用[49]。IAA主要合成于茎尖分生组织、发育中的果实等细胞快速分裂的组织,并通过多种途径向作用部位运输。
生长素的生物合成、极性运输和信号转导有关基因发生突变,均会对株高造成影响。玉米Vanishing tassel2(Vt2)基因通过色氨酸依赖途径中的吲哚-3-丙酮酸途径参与IAA的生物合成,突变后IAA含量下降,雄穗、雌穗发育严重缺陷,叶片数减少,株高降低约55%[35]。Terminal ear1(ZmTE1)是株高的关键调节因子,通过介导IAA的极性运输来维持节间细胞伸长和居间分生组织形成。ZmTE1突变后,IAA的极性运输受到抑制,从而影响细胞分裂和细胞伸长,使玉米节间缩短、株高降低约40%[34]。而br2突变体则是由于ABCB1/PGP1基因异常表达所致。该基因在茎尖表达,将IAA从玉米节运输至节间部位;突变后生长素极性运输受阻,节间伸长被抑制,从而使株高降低[50]。研究表明,玉米中的Brevis plant1(bv1)基因与水稻矮秆基因D50同源,编码具有WD40结构域的肌醇多磷酸5-磷酸酶,对生长素信号转导起关键作用[51]。该基因突变会造成肌醇磷酸酶结构域的保守残基发生非同义替换,使IAA介导的节间薄壁细胞排列异常,从而导致节间缩短,株高降低约70%[15]。
2.1.2 赤霉素对玉米株高的调控 19世纪末,日本的水稻中流行一种会使植株异常徒长的恶苗病,科学家从患有恶苗病的水稻中分离出了这种引起稻苗徒长的物质并命名为赤霉素(gibberellin, GA)[52]。目前已有130余种赤霉素被报道,分别参与了种子萌发、株高调控、花器官发育、结实等多个生物学过程[53],对调控植物生长发育以及应对生物和非生物胁迫方面发挥重要作用[54]。
赤霉素的生物合成可分为3个阶段:在质体中进行从异戊烯焦磷酸到贝壳杉烯阶段;在内质网上进行从贝壳杉烯到GA12醛阶段;在细胞质中进行将无活性的GA12醛转化成有活性GA,该过程需经过GA20氧化酶、GA3氧化酶等催化[53]。Teng等[55]利用综3与其染色体片段代换系SL15杂交的群体,在第3染色体上鉴定到一个与株高有关的主效QTL qPH3.1。并进一步精细定位将候选基因确定为编码GA3 β-羟化酶的ZmGA3ox2,其编码区大片段缺失会使GA合成受阻、植株变矮。与导致严重矮化的矮秆突变体不同,qPH3.1将株高降低约10%,并且不影响产量相关性状和开花期,因此ZmGA3ox2可用于玉米分子标记辅助选择育种[55-56]。
赤霉素的信号转导主要受DELLA蛋白和GID1(GA-insensitive dwarf)受体蛋白调控[54]。玉米中的两个编码DELLA蛋白的基因座Dwarf8(D8)和Dwarf9(D9)影响GA的信号转导。二者突变后DELLA结构域的N端保守区均发生变异,使得GAGID1-DELLA复合体形成受阻,GA信号转导被抑制,从而造成玉米植株矮化[21]。
2.1.3 油菜素内酯对玉米株高的调控 油菜素内酯(brassinosteroid, BR)参与茎秆细胞伸长、根系生长、维管束分化、叶片脱落等多个植物生长发育阶段。BR的生物合成途径与赤霉素较为相似,二者均以异戊烯焦磷酸为基本单位进行合成。从菜油甾醇开始,经过C-6氧化途径,最终形成BR[57]。Nana plant2(Na2)编码d24-甾醇还原酶,参与BR早期的生物合成。Na2功能缺失使BR合成途径中24-亚甲基胆固醇还原为菜油甾醇的过程受阻,BR含量下降,导致植株分蘖减少、叶片直立、严重矮化[30]。而Lilliputian1(Lil1)和Brassinosteroid-deficient dwarf1(Brd1)则参与BR生物合成的最后一步,二者互为等位基因,均编码细胞色素P450蛋白家族的BR C-6氧化酶,突变后植株体内油菜素内酯合成受到抑制,株高降低[13,28]。
BRI1是BR在质膜上感知信号的主要受体,与BAK1相结合形成异源二聚体,使BR信号传递到细胞核,进而激活驱动细胞生长的BR响应基因的转录[57]。BR信号转导有关基因突变会使植株器官生长发育严重受损,进而形成矮秆表型[58]。玉米Brassinosteroid insensitive1a(ZmBRI1a)基因编码一个富含亮氨酸的受体激酶,该激酶位于质膜上,含有一个与BR结合的胞外结构域,该基因功能缺失后会使BR的信号转导途径受抑制,减少细胞分裂和伸长,从而使节间缩短导致植株矮化[14]。
2.1.4 其他激素对玉米株高的调节 近年来,除生长素、赤霉素和油菜素内酯途径调控株高,还发现乙烯、独脚金内酯(strigolactone, SL)等激素也对株高起到调节作用。独脚金内酯是2008年发现的,被称作第七类植物激素,由植物根系分泌的类胡萝卜素经过酶促反应后衍生而来,可以抑制侧芽生长[59]。Guan等[36]发现玉米中编码类胡萝卜素裂解双加氧酶的基因Carotenoid cleavage dioxygenase8(ZmCCD8)敲除后影响SL的生物合成,使茎秆变细、节间缩短、根发育延迟、株高降低约10%。Li等[31]研究表明,玉米半矮秆突变体Semidwarf3(Sdw3)株高降低的原因是乙烯生物合成途径中限速酶ZmACS7的C端结构发生了改变,提高了ZmACS7蛋白的稳定性,增加了Sdw3突变体中乙烯的含量,使得节间细胞的纵向伸长被抑制,最终形成矮化表型。
2.2 非激素途径
目前已克隆的大部分矮秆基因多受激素调控,而还有部分突变体通过改变微管骨架排布或影响成花转变来调控玉米株高。细胞骨架包含微管骨架和微丝骨架两部分,对植物生长发育起到调节作用。微管可以引导植物细胞向指定的方向极性生长,该过程受一系列微管结合蛋白(microtubuleassociated proteins, MAPs)调控。玉米Reducing plant height1(ZmRPH1)基因编码一个MAP蛋白,过表达ZmRPH1使下胚轴和节间长度均有所缩短,植株呈现矮秆表型。过表达ZmRPH1材料根表皮细胞的横向微管比例不足10%,与野生型(93%)相比显著减少,且节间表皮细胞和薄壁细胞长度也显著减少,表明ZmRPH1基因通过调节微管延伸方向来控制玉米株高[41]。过表达ZmRPH1基因在降低玉米株高的同时没有显著减少玉米产量,可以将其应用到玉米矮秆品种改良中。
株高与开花期紧密相关,一般而言,开花期短则株高矮,反之亦然,因此,影响开花期的基因常常影响株高。据报道,光敏色素在植物的光周期和开花期控制中发挥重要作用。当植物处在植被茂密的环境中,会感受到光竞争,从而引发避荫反应综合征(SAS),表现为株高和穗位高增加、输送到籽粒的营养物质减少、吐丝-散粉间隔期延长、雄穗发育异常等,最终造成作物减产。因此,解析玉米中的SAS调控机制可以为选育耐密植的玉米品种提供指导。Phytochrome C1(ZmPHYC1)、ZmPHYC2是玉米的两个光敏色素C基因,研究表明,ZmPHYC1与ZmPHYC2双敲除突变体在长日照条件下表现出中度的早花表型,而过表达ZmPHYC2会使植株株高适度降低[40]。ZmPHYC1和ZmPHYC2可以作为玉米基因工程的潜在靶位点,为创制适合高密度种植的株型结构作出贡献。
3 玉米矮秆品种的选育
目前,玉米矮秆育种中研究利用最多的基因仍是Br2。20世纪50年代,墨西哥育种家率先利用br2种质资源培育出本雀·利比亚AN-360超矮秆玉米杂交种,在超高密度(8000株/亩)种植条件下能达到1000 kg亩产量[60]。然而这种超高密度的种植方式,在带来高产的同时也大大增加了农药、化肥等用量,不利于土地资源的可持续发展[61]。我国在玉米矮化育种之路上也取得了一定成果。研究人员利用br2的矮生特性,经过连续12代的“高矮杂交、选择矮株”,以改善br2叶片重叠、结实率低等不利性状,育成了矮秆玉米杂交种矮单268,该品种在4500株/亩的种植密度下,平均亩产量为636 kg,高产攻关田亩产量可达903 kg[62]。除了常规育种手段,育种家们也在挖掘与创造Br2的有利突变。Wei等[63]定位到了Br2的一个新等位基因qpa1,该基因在最后一个外显子上存在241 bp缺失,突变后可以极大地改善株型结构,并对产量没有负面影响。d129是通过EMS诱变技术获得Br2的等位突变,其第二个外显子上发生了G-A的碱基替换[6]。而br2-sil则是通过太空育种产生的,在Br2的第5个外显子上存在着一个逆转录转座子的插入[64]。这些优良等位变异将助力玉米矮秆品种选育。
相较于单基因矮秆突变体,多基因调控的矮秆表型是由数个基因共同作用所形成,效应更为温和,是株型改良颇具潜力的遗传资源。沈阳农科院利用美国杂交种3147选育出的多基因矮生系5003及其姊妹系5005,是玉米优良核心种质,目前育种中大面积应用的自交系郑58(株高140 cm)、478(株高160 cm)、浚9058(株高165 cm)均为含有5003血缘的矮秆自交系[65]。这些亲本育种材料都曾培育出具有代表性的玉米品种,例如以478为母本育成的玉米杂交种掖单13,其株高在250 cm左右,在现代育种历程中发挥着重要作用。
近20年来随着育种技术的发展,我国育种家通过系谱法、分子标记辅助育种等技术,选育出多个综合性状优良的株高小于250 cm的中矮秆品种。如登海661、登海1717、丰德存玉22、金博士825、鹏玉35等。北京市农林科学院玉米研究所创制培育的新一轮黄改血缘骨干亲本自交系——京2416,已组配育成国审品种40多个,具有良好控制株高的效果。例如将京92与京724组配育成的京科968株高达300 cm以上,而用京2416与京724组配育成的国审品种MC738的株高为248 cm。随着越来越多矮化玉米自交系及杂交种的育成,预示着我国或将进入玉米矮秆育种新阶段。
基于目前对玉米杂交种植株高度标准划分尚未进行明确界定,本团队赵久然多年研究认为,玉米株高可划分为5个等级:(1)超高秆,>340 cm;(2)高秆,280-340 cm[(310±30)cm];(3)中秆,220-280 cm[(250±30)cm],其中250-280 cm可称为中高秆,220-250 cm可称为中矮秆;(4)矮秆,160-220 cm[(190±30)cm];(5)超矮秆,<160 cm。目前,我国近十年所育成的表现优异的矮化玉米新品种,株高距矮秆标准尚有一定距离,仍需加大玉米矮秆育种研究力度,在矮秆有利等位基因鉴定、矮秆分子标记开发、矮秆种质资源创制等方面开展创新型研究[4]。
4 总结与展望
在20世纪60年代,世界人口迅速增加,全球均面临着粮食短缺的危机[66]。国际玉米小麦改良中心的Norman Borlaug利用两个Reduced plant height(Rht)半矮化基因Rht-B1b和Rht-D1b,成功培育出一系列矮秆、高产、耐肥、抗倒伏的小麦新品种,并迅速在墨西哥和印度广泛种植[67]。同时期,国际水稻研究所利用水稻矮秆基因Semidwarf1(SD1)育成半矮秆水稻新品种IR8,提高了水稻抗倒伏性,进而提升了收获指数[68]。这一系列通过矮化育种技术提高作物新品种产量的农技改革运动,使世界粮食总产量得以大幅提升,极大程度改善了粮食短缺问题,被誉为“绿色革命”[69]。推动绿色革命的SD1与Rht1基因在水稻和小麦育种中早已被大规模应用,而玉米中的同源基因D8功能缺失后,在降低株高的同时,分蘖增加、延迟开花、雌穗着生花药,这些农艺性状都是玉米生产的不利因素,因此没有像水稻、小麦一样普遍应用。
目前已被克隆的玉米矮秆基因有近40个,却仍未培育出突破性矮秆品种,其可能原因如下:(1)目前玉米中鉴定到的矮秆基因表型多为极端矮化,对产量造成的负面影响较为严重[10];(2)商业化的玉米品种几乎均为杂交种,而株高性状的杂种优势较强,杂交种很容易受双亲基因组间的遗传互作而表现出较高的株高,遮盖亲本自交系中株高基因的效应;(3)玉米矮秆基因可能与调控其他性状的基因紧密连锁,这些矮秆基因突变时会产生很多育种中的不良性状,密植后产能未得到明显提升,无法直接应用于玉米育种中[22]。
尽管玉米矮化育种充满挑战,但是不可否认,矮秆品种具有广阔的应用前景。以拜耳为代表的跨国种业公司已意识到耐密植玉米的潜力,将矮秆玉米研究工作放在了首要位置,计划今年推出系列玉米矮秆品种(https://www.chinaseed114.com/news/25/news_122520.html)。通常来说,玉米株高与产量呈正相关,单基因矮秆突变体对产量影响剧烈,难以得到较为理想的应用。可以将目标瞄准到多个群体共定位的主效QTL位点,抑或通过多个株高QTL聚合的方式,来创制开花期、抗逆性、产量、品质等综合性状优良的玉米矮秆品种。
随着人们对调控关键性状基因的认识与研究逐渐深入,利用生物技术手段创制矮秆种质,或许是助力作物绿色革命的一种新型选择。育种上许多重要性状的遗传变异发生在顺式调控区域,通过采用基因编辑技术对该区域进行不同长度片段的遗传操控,可对目标基因的表达量进行分级调控,进而产生目标表型变异广泛的突变体。Rodríguez-Leal等[70]利用CRISPR/Cas9技术,对番茄CLAVATA3基因的调控区域进行编辑,得到了多种果实大小不同的番茄突变体。我们可以将这种方法应用到玉米矮秆育种中,通过对控制株高基因的启动子区进行编辑,以实现对基因表达量的精细调控。这种突变方式可以对数量性状产生不同程度的影响,从而创造出一系列玉米株高连续变异的材料,以期挖掘出既能适度降低株高又可增加单株产量的育种材料。
自然界中存在一些极为稀有的株高变异类型,在地理分布范围较窄的自然群体中可能会因为等位基因频率较低而难以检测到,可以创建新型群体遗传材料,通过增加稀有等位基因的频率来提高GWAS的检出能力,从而鉴定出更多的矮秆QTLs位点。针对双亲群体鉴定到的株高基因,可通过基因组学的方法,将其在变异范围更广的自然群体或近缘物种间进行扫描,鉴定优异单倍型,进而开发出更有效的分子标记,以应用到玉米株高的分子标记辅助选择中,为种质创新和分子育种提供技术支持。
猜你喜欢
作物学报(2022年3期)2022-01-22 07:11:18
河北果树(2021年4期)2021-12-02 01:14:52
麦类作物学报(2021年5期)2021-09-23 09:19:00
河北果树(2020年1期)2020-02-09 12:31:18
种子(2018年10期)2018-11-14 08:54:00
麦类作物学报(2018年9期)2018-10-09 02:33:56
老友(2017年7期)2017-08-22 02:36:36
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
销售与市场(管理版)(2016年3期)2016-12-29 00:26:25
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
