
短时程光照对小鼠SCN和LHB谷氨酸受体表达的影响
2023-08-30 09:26李佳逸金巧玲王烈成
安徽医科大学学报 2023年8期
李佳逸,金巧玲,王烈成,程 娟
光对生物节律和情绪的调控是通过自感光神经节细胞(intrinsically photosensitive retinal ganglion cell, ipRGC)对哺乳动物的节律中枢视交叉上核(suprachiasmatic nucleus, SCN)和外侧缰核(lateral habenula, LHB)的直接投射[1]。Fernandez et al[1]验证了短时程光周期可诱发小鼠产生抑郁样行为。ipRGC在SCN和LHB脑区的神经末梢释放谷氨酸和垂体腺苷酸环化酶激活肽(pituitary adenylate cyclase activating polypeptide, PACAP)[2],PACAP主要通过与其特异性受体PAC1结合产生作用[3]。抑制SCN内谷氨酸信号,会使得光诱导的节律性相移停止[4],N-甲基-D-天冬氨酸受体亚型2(N-methyl-D-aspartic acid receptor subunit 2,NR2B)是介导光授时信号传递的重要组成部分[5]。LHB内兴奋性突触传递主要编码厌恶刺激和情感状态,这一活动主要由α-氨基-3-羟基-5-甲基-4-异恶唑丙酸受体(α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor,AMPAR)介导[6]。阻断LHB中的NMDAR可以挽救啮齿动物抑郁症模型中的快感缺乏[7]。此外,LHB中PACAP-PAC1信号通路与焦虑和应激障碍的产生有关[8]。该研究将利用免疫印迹(Western blot)的方法,探究短时程光照下,SCN及LHB脑区中谷氨酸信号通路、PACAP-PAC1信号通路及下游关键性蛋白的变化。
1 材料与方法
1.1 实验动物本实验使用的动物均为SPF级、C57BL/6J雄性小鼠,6~8周龄,体质量20~25 g,购自河南斯克贝斯生物科技股份有限公司。小鼠被置于T24光暗循环 (12 h/12 h)环境下,适应1~2周后,进行分组,将实验组小鼠置于T7 (3.5 h/3.5 h) 的光暗循环环境下饲养,对照组小鼠继续置于T24 (12 h/12 h) 的光环境下饲养,实验期间自由提供水和食物,4~6周后用于实验。所有动物实验操作均符合安徽医科大学动物伦理委员会的要求。
1.2 主要抗体Mouse anti-GAPDH购自美国Santa公司;HRP Goat anti-Mouse、IgG HRP Goat anti-Rabbit IgG购自江苏Affinity公司;Rabbit anti-NR2A、Rabbit anti-NR2B、Rabbit anti-P-PKA购自江苏Affbiotech公司;Mouse anti-PAC1购自上海Absin公司;Mouse anti-GluR1购自美国Novus公司;Rabbit anti-GluR2购自成都正能生物。
1.3 主要仪器垂直电泳槽购自美国Bio-Rad公司;低温高速离心机购自德国Eppendorf公司;电子分析天平购自德国Sartorius公司;研磨仪购自武汉塞维尔公司;凝胶成像显影仪购自上海培清公司; 纯水仪购自合肥宏科公司;滚轴摇床购自海门海宁其林贝尔仪器公司; 金属浴购自杭州米欧公司。
1.4 方法
1.4.1溶液配制 ① 10×三羟甲基氨基甲烷(hydroxymethyl aminomethane, Tris)-甘氨酸缓冲液:精确称量144 g甘氨酸,30.3 g Tris,定容至1 L,混匀后,室温保存。② 1×电泳溶液:取100 ml 10×Tris-甘氨酸缓冲液定容至1 L,称取1 g十二烷基硫酸钠(sodium dodecyl sulfate, SDS),混匀后4 ℃储存。③ 1×转膜溶液:取100 ml 10×Tris-甘氨酸缓冲液,加入200 ml甲醇,定容至1 L,混匀后4 ℃ 储存。④ 10×Tris缓冲液:称取24.2 g Tris加入双蒸水800 ml,用HCl调pH至7.6后加入80 g NaCl,混匀后,定容至1 L,4 ℃储存。⑤ 1×吐温三羟甲基氨基甲烷缓冲液(tris buffered saline with Tween, TBST)溶液:称取100 ml 10×三羟甲基氨基甲烷缓冲液(tris buffered saline, TBS)定容至1 L,加入1 ml吐温20,混匀后4 ℃储存。
1.4.2组织蛋白的提取 授时因子时间(zeitgeber time, ZT)为人为控制的光暗时间,以ZT 0为开灯时间,在24 h内,组织样品的取样时间分别是ZT 1、ZT 5、ZT 9、ZT 13、ZT 17、ZT 21,夜间配戴红外头灯取样。取样在光学显微镜下利用眼科手术器械操作,选取年龄体质量相近的小鼠,全程低温通氧。样品获取后分别置于2 ml离心管中,加入200 μl RIPA裂解液(加1%蛋白酶抑制剂)于离心管中,在4 ℃研磨仪内充分研磨。用无菌的镊子取出离心管中的钢珠,放入提前预冷的4 ℃ 低温超速离心机进行离心, 12 000 r/min,4 ℃,15 min。离心后吸取上清液,加入5×蛋白上样缓冲液,100 ℃、10 min煮样,分装后-80 ℃ 保存。
1.4.3配制十二烷基硫酸钠(sodium dodecyl sulfate, SDS)凝胶 分离胶:5.9 ml ddH2O,5.0 ml 30%聚丙烯酰胺,3.8 ml 1.5 mol/L Tris-HCl(pH 8.8),0.15 ml 10%SDS,0.15 ml 10%过硫酸铵,0.006 ml四甲基乙二铵(N,N,N',N'-tetramethylethylenediamine, TEMED)。按照10%的配方加入试剂,最后加入200 μl异丙醇,室温放至凝固。浓缩胶:4.1 ml ddH2O,1.0 30% 聚丙烯酰胺,0.75 ml 1.0 mol/L Tris-HCl(pH 6.8),0.06 ml 10%SDS,0.06 ml 10%过硫酸铵,0.006 ml TEMED。将异丙醇倒出,按照5%的配方加入试剂,充分混匀后,立刻灌入玻璃板,室温放至凝固。
1.4.4SDS-聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis, PAGE)设置凝胶电泳参数 恒压80 V 20 min,切换电压为恒压100 V,直至所需目的蛋白充分分离后停止。
1.4.5转膜与显色 电泳结束后,将分离胶放入转膜液中,设置参数为恒流260 mA、120 min。转膜结束后,封闭2 h。弃去封闭液,加入1 × TBST洗涤3次,1次/1 min。将谷氨酸受体和下游分子受体的一抗按以下的稀释浓度配制:NMDAR-2A(NMDA receptor 2A, NR2A) (1 ∶500); NMDAR-2B(NMDA receptor 2B, NR2B)(1 ∶500); AMPAR1(AMPA receptor 1, GluR1) (1 ∶1 000);AMPAR2(AMPA receptor 2, GluR2)(1 ∶1 000); anti-PAC1 (1 ∶500); 磷酸化的蛋白激酶A(phosphorylated protein kinase A, P-PKA)(1 ∶1 000); 磷酸化的细胞外调蛋白激酶(phosphorylated extracellular regulated protein kinases, P-ERK)(1 ∶1 000); Ca2+/钙调素依赖性蛋白激酶Ⅱ(calcium-CaM-dependent protein kinase II, CaMK II)(1 ∶1 000); 甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH) (1 ∶1 000), 4 ℃过夜。取出膜,加入1×TBST洗涤3次,1次/10 min。二抗孵育1 h,HRP Goat anti-Mouse IgG/HRP Goat anti-Rabbit IgG (1 ∶10 000)。弃去二抗, 1× TBST充分洗涤,3次/10 min。将膜置于凝胶成像系统中显影。
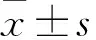
2 结果
2.1 短时程光周期影响SCN谷氨酸受体和PAC1受体的表达与T24光周期相比,T7光周期下SCN的PAC1受体变化差异无统计学意义(图1B);与T24光周期相比,在T7光周期下,GluR1的总体表达呈现低于对照组的趋势,但差异无统计学意义(图1D);在ZT 1和ZT 5,与T24光周期相比,T7光周期的GluR2蛋白表达上升(图1F:t=3.565,P=0.016 1;t=4.064,P=0.009 7),且GluR2的表达均在ZT 13达到峰值;与T24光周期相比,T7光周期下NR2B的表达整体上升(图1J,P=0.011 5),且T7光周期下和T24光周期下小鼠SCN中NR2B的表达均在ZT 13达到峰值。与T24光周期相比,T7周期下GluR2的总体表达上调(图1F,P=0.006 7),结果显示SCN中GluR2和NR2B在T7光周期下表达上升,表明短时程光照可能是通过影响谷氨酸受体的表达从而影响生物节律的稳定。
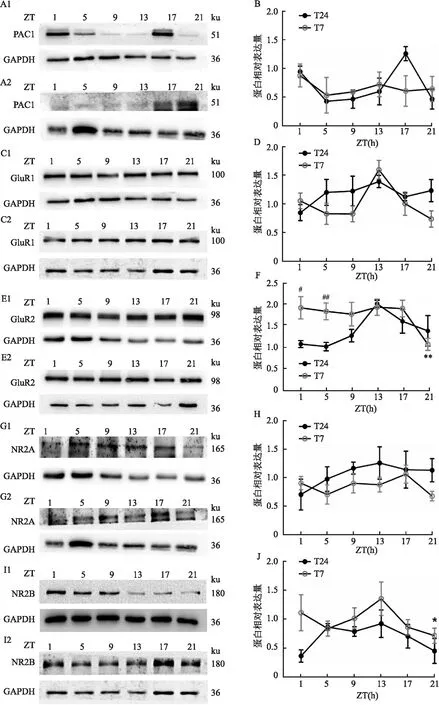
图1 T24及T7光周期对SCN的谷氨酸受体和PAC1受体的影响A、C、E、G、I:T7或T24周期下PAC1、NR2A、NR2B、GluR1、GluR2受体蛋白表达的代表图;1:T24;2:T7;B:SCN中PAC1受体的表达;D、F:SCN中GluR1、GluR2的表达;H、J:SCN中NR2A、NR2B的表达;与T24光周期蛋白整体表达比较:*P<0.05,**P<0.01;与T24光周期比较:#P<0.05,##P<0.01
2.2 短时程光周期影响LHB谷氨酸受体和PCA1受体的表达与T24光周期相比:T7光周期下LHB的PAC1受体变化差异无统计学意义(图2B),但整体呈现表达减少的趋势;T7光周期下不同时间点和24 h内GluR1总体的表达差异无显著性(图2D);T7光周期下GluR2的总体表达下降(图2F,P=0.002 4);而T7周期下不同时间点的NR2B差异无显著性(图2H)。结果说明,短时程光周期下LHB脑区中谷氨酸受体表达显著减少,提示小鼠的抑郁样行为可能与GluR2的表达显著降低有关。
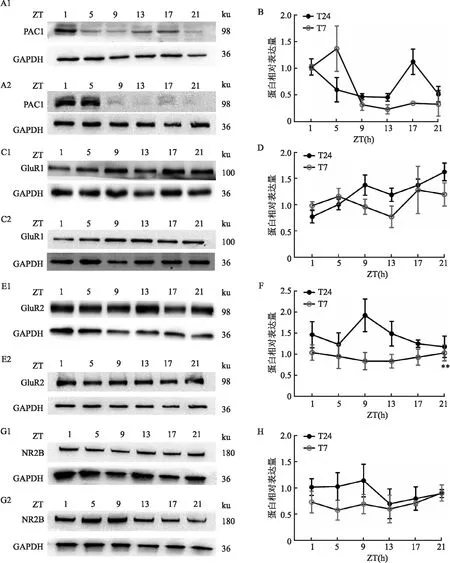
图2 T24及T7光周期对LHB的谷氨酸受体和PAC1受体的影响A、C、E、G:T7或T24周期下PAC1、GluR1、GluR2、NR2B受体蛋白表达的代表图;1:T24;2:T7;B:LHB中PAC1受体的表达;D、F:LHB中GluR1、GluR2的表达;H:LHB中NR2B的表达;与T24光周期蛋白整体表达比较:**P<0.01
2.3 短时程光周期下SCN中下游信号分子的表达与T24光周期相比,T7光周期下SCN中各个时间点CaMK II的表达差异无统计学意义(图3B);T7光周期下SCN中P-PKA的表达高于T24光周期,但差异无统计学意义(图3D)。
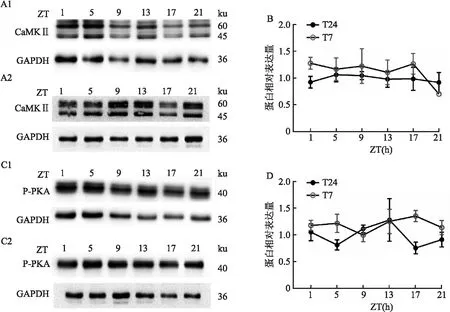
图3 T24及T7光周期对SCN中下游信号分子表达的影响A、C:T24或T7光周期下SCN下游信号分子蛋白表达的代表图;1:T24;2:T7;B:SCN中CaMK II的表达;D:SCN中P-PKA的表达
2.4 短时程光周期对LHB中下游信号分子的表达的影响T7光周期下,LHB中CaMK II表达与T24光周期相比,无显著性差异(图4B),而与T24周期相比,T7周期下LHB的P-ERK的表达整体上升(图4D,P=0.046)。这些结果说明在T7光周期下小鼠产生抑郁样行为可能是由于上调LHB中P-ERK的表达导致。
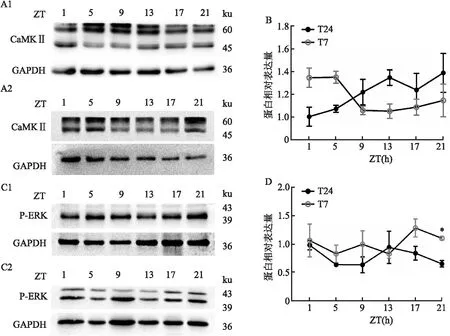
图4 T24及T7光周期对LHB中下游信号分子表达的影响A、C:T7或T24周期下LHB下游信号分子蛋白表达的代表图;1:T24;2:T7;B:LHB中CaMKII的表达;D:LHB中P-PKA受体的表达;与T24光周期蛋白整体表达比较:*P<0.05
3 讨论
研究[3,9]表明,光对于生物节律稳态的调控主要是通过ipRGCs-SCN信号通路。在T24光周期下,PACAP mRNA在ipRGCs中的表达呈现夜间高于白天,且在节律时间(CT)17达到峰值[2]。本研究结果显示,在T24光周期下,SCN中PAC1受体的表达在ZT 17达到峰值,且在夜间呈上升的趋势,但T7光周期下在ZT 17 PAC1受体的表达减少。谷氨酸作为ipRGCs末梢释放的主要神经递质,谷氨酸-NMDA受体信号通路对维持生物节律性振荡的稳定具有重要作用[2-3],SCN中NR2B的表达呈昼夜节律性振荡,且在夜间达到峰值[10]。本研究结果显示,关灯1 h后NR2B的表达达到峰值,在T7光周期下NR2B的24 h内总体表达量高于正常光周期,提示短时程光照可能扰乱了NR2B表达而影响节律的稳定。研究[2]证实,SCN内主要表达AMPA型受体的mRNA在SCN和周围区域大量表达。本研究结果显示,在T7周期下,ZT 1、ZT 5的GluR2的表达上调,但GluR1表达在ZT 13达到峰值,表明短时程光周期可能是通过影响谷氨酸受体的表达从而影响生物节律的稳定。
Fernandez et al[1]验证了T7光周期可使得小鼠产生抑郁样行为,同时揭示了短时程光照诱发的抑郁样情绪与LHB核团神经元活动具有直接关系。LHB是一个较小的双侧上皮核,对厌恶刺激做出反应并进行信息的整合,并促进对未来不良事件的回避来调控情绪[9]。位于LHB的前内侧区域的稀疏分布的神经元群体表达PACAP,它也在其他几个大脑区域表达,并且在应激相关核中表达丰富[8]。本研究结果显示,在T7周期下,PAC1受体表达减少。睡眠稳态的维持和情绪的调控存在直接关联,研究[11]表明LHB参与觉醒睡眠周期的调节,其神经元兴奋具有促进REM睡眠作用,这也可能是LHB调控情绪的又一机制,睡眠稳态维持与LHB中与情绪调控相关蛋白表达之间的关系有待进一步探索。
阻断LHB中NMDAR依赖的神经元爆发性活动,可介导氯胺酮在大鼠和小鼠抑郁症模型中的快速抗抑郁作用[12]。在抑郁症模型下,激活NMDAR的表达,能提高LHB神经元放电频率,且特异性阻断LHB中NMDAR可以挽救享乐缺乏和行为绝望等病理表型[13]。本研究结果显示,在T7光周期下GluR2的整体表达低于T24光周期,推测短周期光可导致小鼠抑郁样行为可能与谷氨酸受体的表达异常有关。研究[14]表明,抑郁症会增加谷氨酸释放到含有AMPAR的LHB突触上的概率,并通过CaMK II 蛋白的过表达增强AMPAR功能。本研究表明T7或T24光周期下,CaMK II的表达与AMPAR的表达趋势一致。
猜你喜欢
遗传(2023年9期)2023-09-25
农业科技通讯(2023年1期)2023-02-12
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
三门峡职业技术学院学报(2017年1期)2017-06-05
中国洗涤用品工业(2017年2期)2017-04-16
中国比较医学杂志(2017年5期)2017-01-17
广西林业科学(2016年3期)2016-03-16
医学研究杂志(2015年12期)2015-06-10
医学研究杂志(2015年5期)2015-06-10
