
不同抗虫性茶树品种的叶色及挥发物差异性研究
2023-08-30 14:19:24孙悦刘梦月高晨曦吴全金曹士先余顺甜陈志丹金珊孙威江
茶叶科学 2023年4期
孙悦,刘梦月,高晨曦,吴全金,曹士先,余顺甜,陈志丹,金珊,孙威江*
不同抗虫性茶树品种的叶色及挥发物差异性研究
孙悦1,3,刘梦月2,3,高晨曦1,3,吴全金4,曹士先5,余顺甜5,陈志丹2,3,金珊1*,孙威江1,3*
1. 福建农林大学园艺学院,福建 福州 350002;2. 福建农林大学安溪茶学院,福建 泉州 362400;3. 海峡两岸特色作物安全生产省部共建协同创新中心,福建 福州 350002;4. 福建开放大学文经学院,福建 福州 350003;5. 武夷星茶业有限公司,福建 武夷山 354301
为探究不同抗虫性茶树品种的叶色及挥发物差异性,对11个茶树品种进行为期2年的田间虫口密度调查,通过田间枝条室内孵化试验和室内生命参数测定试验,佐证了田间种群密度作为抗性分级标准的可靠性;并测定色差及叶片单位叶面积蜡质含量,用气相色谱-质谱联用仪(GC-MS)对未被病虫为害的一芽二叶新梢的挥发物进行检测,以期筛选不同抗虫性茶树品种间的差异物。结果显示,小贯松村叶蝉()趋向于叶片色彩较暗、黄色较淡的茶树品种;茶棍蓟马()趋向色彩鲜亮、饱和度高、黄色偏高的茶树品种,且茶棍蓟马更重视叶背的色泽。在挥发物方面,小贯松村叶蝉种群密度与芳樟醇的相对含量呈显著正相关,与壬醛、十二烷的相对含量呈显著负相关。茶棍蓟马的种群密度与十二烷和苯乙醇的相对含量呈显著正相关。十二烷对小贯松村叶蝉和茶棍蓟马作用不同,其含量和茶树叶片蜡质含量与种群密度间的关系呈现相同趋势,这说明同一种挥发物对不同昆虫会产生不同的趋向反应。
茶树;小贯松村叶蝉;茶棍蓟马;叶色;挥发物;抗性
小贯松村叶蝉()属半翅目(Hemiptera)叶蝉科(Cicadellidae),以成、若虫刺吸为害茶树嫩梢芽叶。茶棍蓟马()属缨翅目(Thysanoptera)蓟马科(Thripidae),以锉吸式口器吸食茶树新梢汁液,虫体微小、生活隐蔽,为害山茶科的多种经济作物,尤以茶树最为严重[1]。近年来,茶园中小贯松村叶蝉和茶棍蓟马为害发生频繁,大发生年份常导致高达50%以上的茶叶产量受损,年防治费用占茶树病虫害总防治费用的60%以上[2]。
植物与昆虫长期协同进化过程中,害虫在接触寄主前,会利用触角的化学感受器对植物挥发物进行探测和识别,利用视觉对植物光感、色感进行判断,根据所获信息做出相应的行为选择[3]。而植物自身也逐渐形成了以次生性物质为主的化学防御体系,其中最为重要的是释放挥发性有机化合物(Volatile organic compounds,VOCs)[4]。植物通过释放挥发物,不仅能减少害虫的数量[5],还可“警告”邻近植物有害虫或病原微生物来袭[6]。这种寄主本身具有的对害虫产生吸引或避害的作用,属于品种抗虫或感虫特性的范畴[7]。从该角度切入,选育高抗品种用于有害生物综合治理(Integrated pest management,IPM)将更生态、更经济。目前较多关于茶树挥发性物质对小贯松村叶蝉的引诱或趋避、毒杀作用的报道[8-10],但尚未见针对茶棍蓟马或同时对两种取食方式接近的害虫有趋向作用的化合物筛选的研究报道。
本研究基于2年的田间虫口密度调查,对11个茶树品种抗小贯松村叶蝉、茶棍蓟马的程度进行抗性分级,以未被病虫为害的茶树新梢为材料,研究不同抗虫性级别茶树品种的叶色差异及鲜叶挥发物组成,以期探究茶树叶片及挥发物对小贯松村叶蝉和茶棍蓟马具有趋向作用的成分,为有效选育茶树抗虫品种提供科学依据,并为生态调控、种群监测及开发植物源引诱或趋避剂提供参考。
1 材料与方法
1.1 试验环境与材料
2021年、2022年在福建省武夷山市武夷星茶树种质资源圃开展试验,资源圃地处东经118º0′3.82″,北纬27º42′48.99″,海拔217.4 m,属亚热带季风性湿润气候。供试茶树品种为白鸡冠(BJG)、黄玫瑰(HMG)、黄观音(HGY)、瑞香(RX)、大红袍(DHP)、白牡丹(BMD)、奇兰(QL)、黄棪(HD)、梅占(MZ)、水仙(SX)、紫娟(ZJ),各品种茶树处于同一管理措施、同一生态环境的地块中,坡度一致,每个品种依次排列5行,约250 m2。调查害虫为小贯松村叶蝉和茶棍蓟马。
1.2 试验方法
1.2.1 茶样采集
2021年4月,采集未被病虫为害的一芽二叶新梢,液氮固样,真空冷冻干燥机干燥30 h,干燥后将样品置于多样品组织研磨仪粉碎,时长1 min,然后装入塑封袋置于–20 ℃冰箱备用。
1.2.2 田间种群密度调查
采用5点取样法[11]对每个品种调查记录茶树嫩梢上的小贯松村叶蝉和茶棍蓟马的虫口发生量(每点50个一芽二叶),2021年6月6日—10月26日,累计调查21次;2022年5月25日—7月27日,累计调查7次。
1.2.3 田间枝条虫口孵化试验
参照金珊[12]的试验方法,2021年7月进行田间枝条虫口孵化试验,20次重复。人工气候室温度为(25±2) ℃,相对湿度为70%~75%,光照时间L∶D=14∶10。
1.2.4 室内生命参数测定
供应枝条:2022年6月初对紫娟、黄棪、黄观音、白鸡冠茶树试验地分别量定10 m茶行进行修剪,并喷施生物农药,5%天然除虫菊水剂800倍液(5% Pyrethrins SL)[13],参照施龙清[14]的方法搭建防虫棚,1周后,再进行1次生物农药喷施,确保茶枝无虫口。7月初,药物挥发散尽,待茶树新梢成熟,采集茶树大小基本一致的嫩梢作为试验枝条,供应虫体营养生长。
饲养虫源:2022年6月,在武夷星茶树种质资源圃的福云6号茶树上捕捉小贯松村叶蝉成虫,用体式显微镜辨别性别,雌雄配对,饲养于养虫笼内的茶枝(福云6号,40~50 cm长、健康无虫害)上,花泥保水培养,放置于温度(25±2) ℃,相对湿度70%~75%,光照时间L∶D=14∶10的人工气候室,饲养3代以上,将同一天孵化的1龄若虫作为供试昆虫。
饲养皿、养虫瓶制作:选择高2 cm,直径9 cm的培养皿,皿盖上均匀扎10个孔,皿内放入大小合适的方块花泥,用该装置作为小贯松村叶蝉饲养皿。在透明塑料杯(底部直径7.2 cm,杯口直径11.9 cm,高16.2 cm)杯璧扎10个小孔,保持杯内通风,倒扣于培养皿中,培养皿(直径14 cm,高1.5 cm)内放置直径为14 cm的滤纸,保水处置,内部装上块状花泥,便于插入枝条,用该装置作为小贯松村叶蝉养虫瓶。
生命参数记录:(1)试验前1 d,将供应虫源的茶树(福云6号)枝条刷净,保证枝条上无虫,取同一天孵出的1日龄小贯松村叶蝉80只;(2)每个饲养皿中放入1只1日龄小贯松村叶蝉,单皿单养,80个重复;(3)每日早晚各加水1次,当天更换1次新梢,更换时记录虫体若虫历期,以发现白色虫蜕确定龄期;(4)虫体从若虫羽化为成虫当天,进行同天羽化雌雄配对,转入养虫瓶内(瓶内加入大小基本一致具木质部的一芽四叶嫩梢,保湿放置),离体新梢采摘于遮虫网内新发的枝条,每天更换养虫瓶内新梢。(5)逐日查看每对雌雄成虫,记录成虫的死亡时间。逐日观察每个品种每次重复被更换的离体新梢,记录若虫孵化时间和头数。若当天羽化的成虫雌雄数量不对等,则使用备用虫源配对补齐。配对后,继续记录试验虫源的寿命。若试验虫源雌虫的配偶先死亡,则随机从备用虫源中为其另选配偶进行配对,但补充的雄虫寿命不统计在内;若雌虫死亡,则雄虫单独培养直至死亡。
1.2.5 色差测定
色差计为ADCI全自动色差计(北京辰泰克仪器技术有限公司),白板校正后,测定芽下第三叶正面与背面,获得L*、a*、b*的测量值,10次生物学重复,通过L*、a*、b*计算出颜色差异值(ΔE)、色调彩度(Cab)、色彩饱和度(Sab),其中各项色差指标中的s下角标代表叶片的表面;b下角标代表叶片的背面。L*是衡量色彩明暗程度的指标,其值越大,表示表面越亮;a*是衡量红绿程度的指标,其为正值时表示偏红色,为负值时表示偏绿色;b*是衡量黄蓝程度的指标,其为正值时表示偏黄色,为负值时表示偏蓝色,绝对值越大表示黄色或蓝色越深。
1.2.6 叶片蜡质含量测定
参照刘勇等[15]的方法进行调整,随机剪取10根40~50 cm带有芽头的茶树枝条插入花泥,黑暗处理3 h,取10个健康完整的芽下第二叶,测量叶片长、宽后清水冲洗,置于烧杯中,加入50 mL三氯甲烷,提取5 s,转移至旋转仪(52 ℃)中浓缩至10~20 mL,倒入已称重过的培养皿(称重质量记为W1),放入通风厨风干后称重记为W2,计算出茶树单位叶面积的蜡质含量。
叶面积(S)计算公式为=0.7×叶长×叶宽;单位叶面积蜡质含量(W蜡)计算公式为蜡=(2-1)/,单位为mg·cm-2
1.2.7 香气挥发物检测
参照吴少玲等[16]的方法进行测定,使用石碧滢等[17]的方法对挥发物定性,峰面积归一化法计算各挥发性成分的相对含量。
1.3 数据分析
采用Excel 2020对数据平均值和标准误进行计算,使用IBM SPSS Statistics 26单因素ANOVA检验进行方差分析;使用SIMCA 14.0进行正交偏最小二乘判别分析(Orthogonal partial least squares-discrimination analysis,OPLS-DA);采用TBtools和GraphPad Prsim 7绘图。
2 结果与分析
2.1 种群发生密度
小贯松村叶蝉与茶棍蓟马对同一生境条件的不同茶树品种为害存在明显偏好差异,各茶树新梢上的虫口种群密度存在显著差异。2年的种群密度平均值结果显示(表1),小贯松村叶蝉总虫口量为每50梢42.60~161.80头,茶棍蓟马总虫口量为每50梢49.30~355.90头。2021年和2022年的两种刺吸式害虫的虫口高发期均集中于6月—7月,处于夏茶期(图1),该时间段,11个茶树品种均遭受2种植食性害虫不同程度的危害。
2.2 抗性分级
将2年的种群密度结果聚类分析,结果如图2所示。11个茶树品种对小贯松村叶蝉的抗性等级聚为3类:紫娟和梅占为感虫(S)等级,水仙、黄棪、奇兰、白牡丹、大红袍和瑞香为中感(M)等级,黄观音、黄玫瑰和白鸡冠为抗虫(R)等级。对茶棍蓟马的抗性等级聚为4类:黄玫瑰为感虫(S)等级,白鸡冠和瑞香为中感(MS)等级,奇兰和黄观音为中抗(MR)等级,白牡丹、黄棪、大红袍、紫娟、梅占和水仙为抗虫(R)等级。
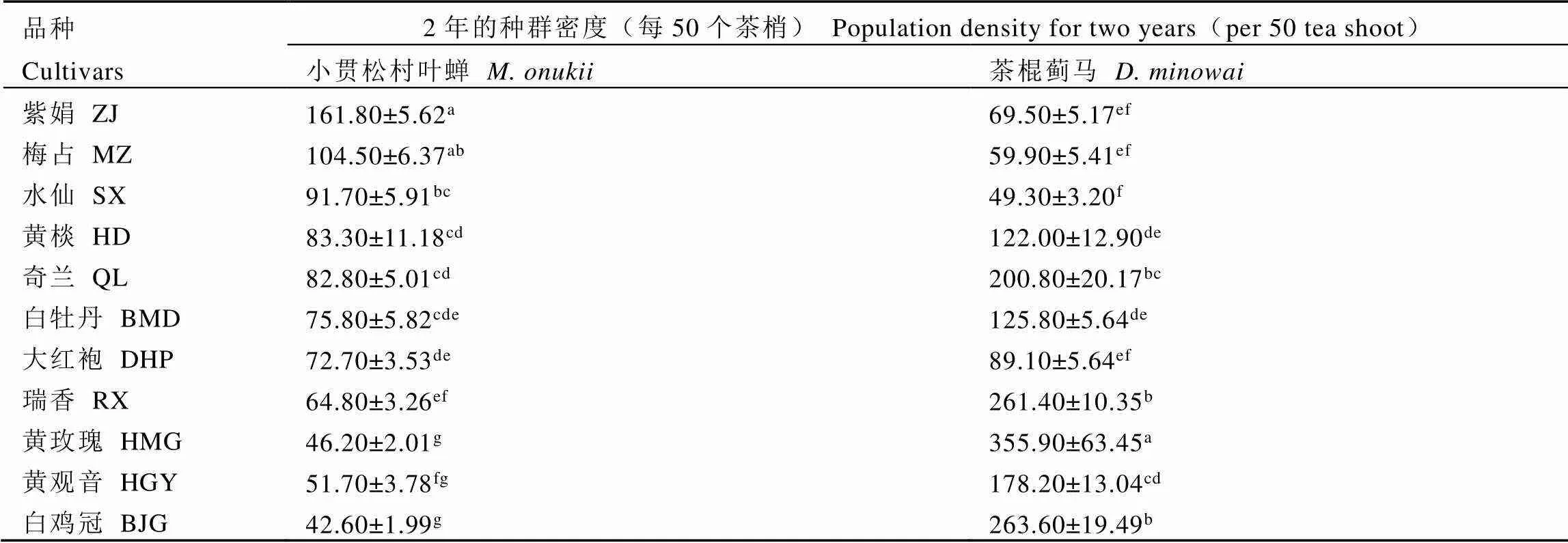
表1 11个茶树品种上小贯松村叶蝉和茶棍蓟马的种群密度
注:表中所列数据为平均值±标准误;同列不同字母表示差异显著(<0.05),下同
Note: Data in the table are mean ± SE. The different letters in the same column represent significant differences (<0.05). The same blew
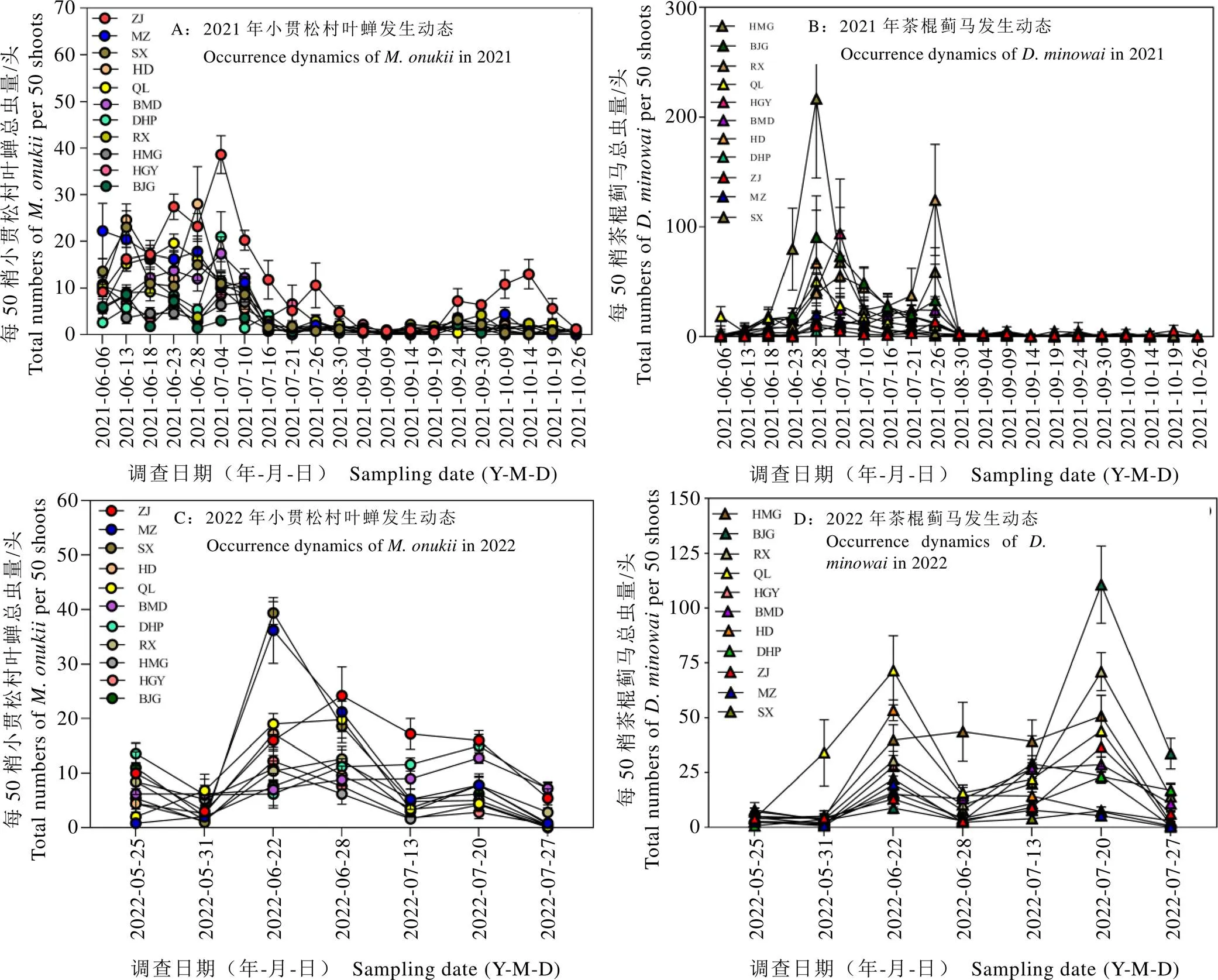
图1 不同年份11个茶树品种的虫害发生动态

注:不同字母代表差异显著(P<0.05)
2.3 室内验证结果
2.3.1田间枝条室内孵化试验结果
茶枝上茶棍蓟马每日的若虫孵化量远高于小贯松村叶蝉,可直观比较茶棍蓟马在不同茶树品种的产卵偏好。将若虫孵化量与种群密度关联分析发现(图3),茶棍蓟马的种群密度与次日的若虫孵化量呈显著正相关(<0.05),与其他天的若虫孵化量均呈正相关趋势。这说明以田间种群密度为抗虫性标准划分茶树对茶棍蓟马的抗性等级,具有一定的可靠性,而茶树对小贯松村叶蝉的抗性分级需进一步验证。
2.3.2 小贯松村叶蝉生命历期
利用2021年的分级结果[18],筛选代表不同抗性级别的茶树品种各1个,分别为紫娟、黄棪、黄观音、白鸡冠。通过室内饲养小贯松村叶蝉,探究不同品种对昆虫生长发育的影响。小贯松村叶蝉若虫有5个龄期,羽化为成虫后才可辨别雌雄。小贯松村叶蝉在各品种上的发育历期如表2所示,1~3龄的若虫历期约1~2 d,4~5龄的平均历期为2~3 d。饲养在黄棪上的若虫发育期显著短于另外3个品种,雌、雄成虫的生命历期均显著长于紫娟和白鸡冠的,说明取食黄棪的小贯松村叶蝉生长发育更快,成虫存活期更长。但取食紫娟和黄棪的小贯松村叶蝉的总体生命周期无显著差异,通过生命周期只能将黄棪、紫娟与黄观音、白鸡冠区分开。
2.3.3 小贯松村叶蝉存活率
观测不同茶树品种上小贯松村叶蝉若虫在各阶段的存活率,进一步比较不同茶树品种对小贯松村叶蝉生存繁衍的适应度。图4所示,4个茶树品种上的若虫存活率在50%~81%,3~5龄、若虫的存活率排序为紫娟>黄棪>黄观音>白鸡冠。若虫在黄观音与白鸡冠上的存活率十分接近,这与生命周期呈现一致规律;在紫娟和黄棪上的存活率差异明显,且高于黄观音和黄棪,黄棪上的小贯松村叶蝉虽然生长发育快,成虫存活期长,但存活率低,这说明紫娟更适宜小贯松村叶蝉的种群发展。按田间种群密度进行抗性划分,这4个品种的抗性表现为白鸡冠>黄观音>黄棪>紫娟;室内室外结果基本一致,佐证了以田间种群密度作为茶树抗虫性的划分标准具有可靠性。
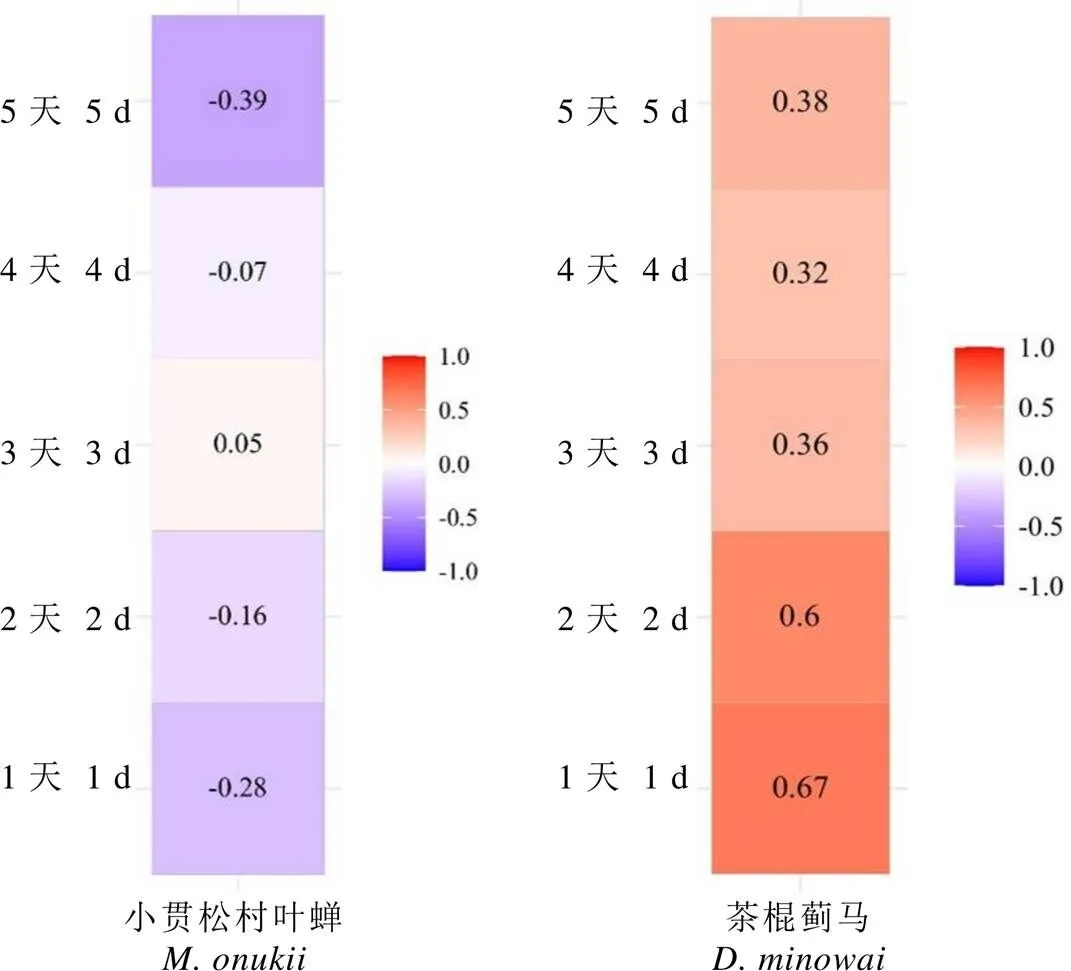
图3 孵化量与种群密度的相关性

表2 4个茶树品种上小贯松村叶蝉各阶段发育历期
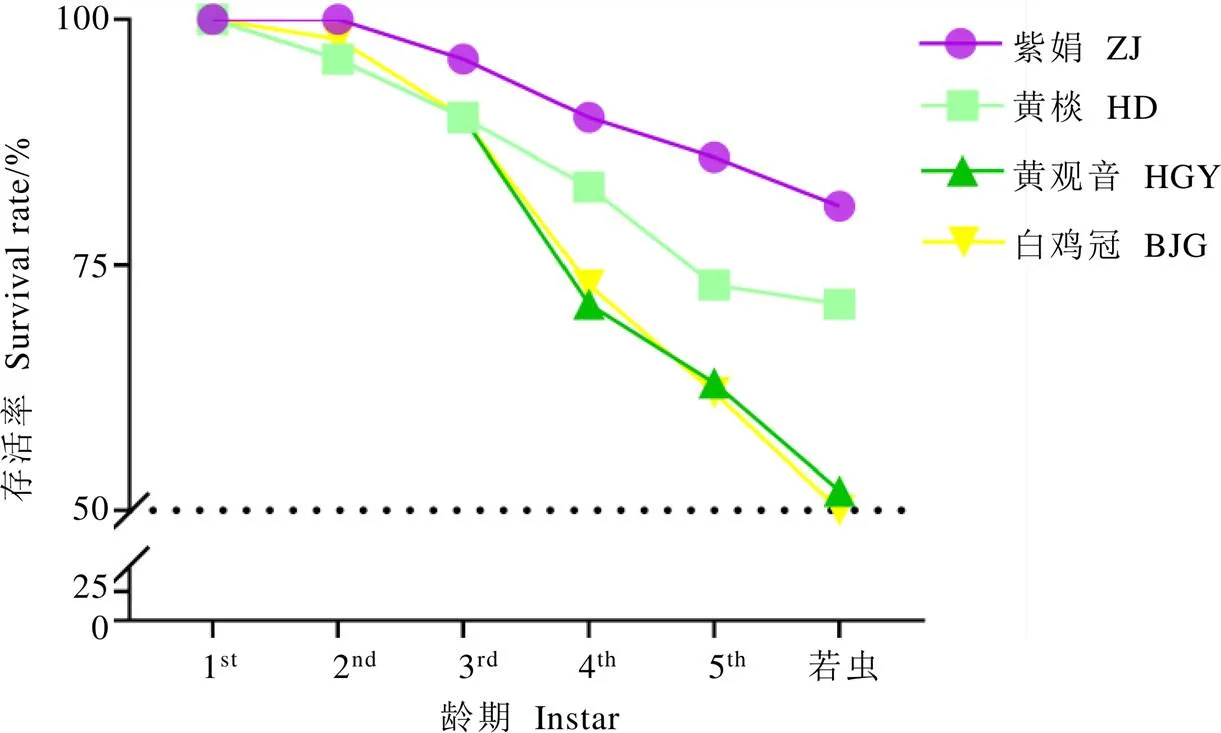
图4 4个茶树品种上的小贯松村叶蝉若虫存活率
2.4 不同茶树品种的色差分析
表3和表4显示,仅白牡丹和梅占的叶背色差不明显(ΔE<1),其他品种间均可被区分。种群密度与茶树叶片的色差具有强相关性(图5),色差各项指标与小贯松村叶蝉种群密度显著负相关(<0.05),与茶棍蓟马种群密度显著正相关(<0.05),表明茶树叶色对小贯松村叶蝉与茶棍蓟马的寄主定位起重要作用。其中a*值(红绿程度)对2种主要刺吸式害虫的寄主定位选择影响均无明显作用,主要是叶片的L*(色彩明暗度)和b*(黄蓝程度)、Cab(色调彩度)和Sab(色彩饱和度)发挥作用,但Sabs、Cab*s和b*s与茶棍蓟马种群密度的相关性不显著,这说明茶棍蓟马在寄主定位选择时更在意叶背的色彩呈现。
2.5 不同抗虫级别茶树品种的挥发物差异分析
通过固相微萃取法(SPME-GC-MS)对11个不同抗性级别的茶树品种一芽二叶新梢挥发物进行鉴定,共鉴定出49种挥发物(表5),共有挥发物12种(图6),表明不同抗虫级别茶树品种间存在明显差异。如图7所示,总挥发物分为8类,醇类14种(33.55%~68.57%),醛类8种(4.69%~25.58%)、酯类9种(2.89%~25.64%)、烯烃类8种(0.58%~2.90%)、烷烃类5种(0.22%~3.80%)、酮类2种(0~1.31%)、酚类1种(0.10%~0.56%)、其他2种(0~1.23%),所有品种间均为醇类挥发物相对含量最高,其次为醛类、酯类。
为探究挥发物中哪类物质参与影响刺吸式害虫的寄主定位选择,将各类挥发物相对含量与种群密度进行关联分析,结果显示(表6),小贯松村叶蝉种群密度与醇类挥发物相对含量呈显著正相关(<0.05,=0.636),与醛类呈负相关但不显著(=–0.446),与烷烃类挥发物相对含量呈显著负相关(<0.05,=–0.668);而烷烃类挥发物与茶棍蓟马种群密度呈极显著正相关(<0.01,=0.737)。说明茶树鲜叶挥发物中参与影响两种刺吸式害虫对茶树寄主定位的主要是醇类和烷烃类,醛类次之。
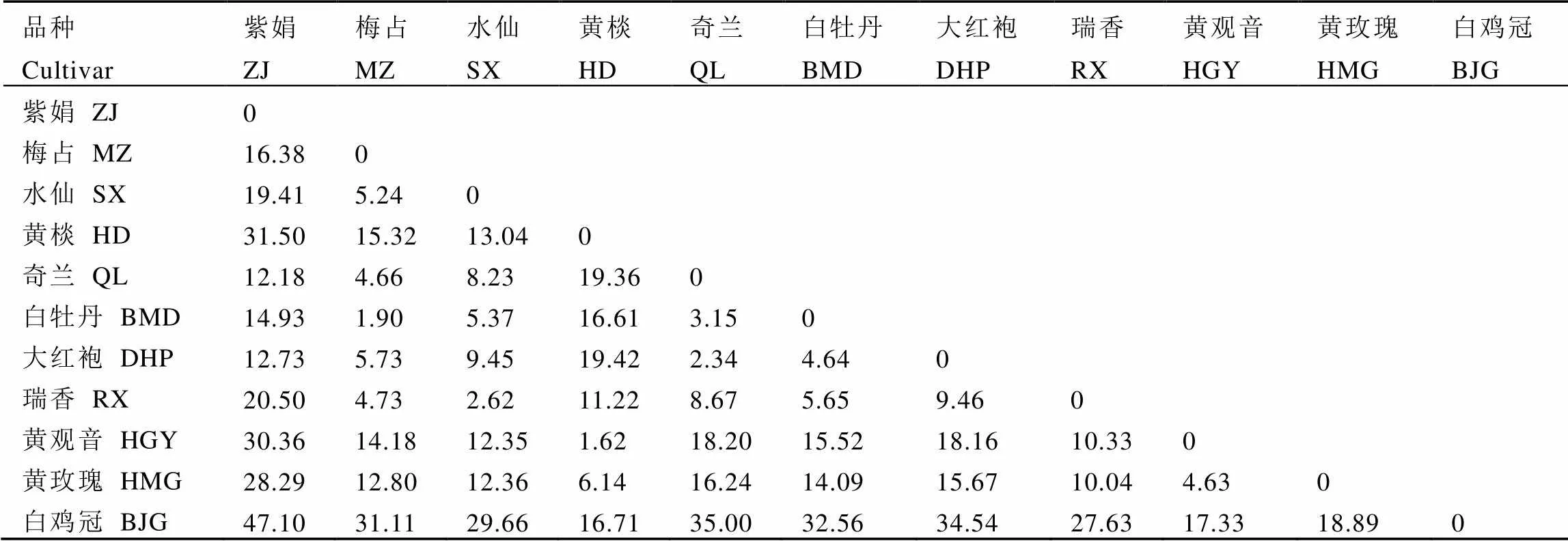
表3 茶树芽下第3叶片叶表色差参数ΔE表
注:ΔE<1表示色差不明显;1<ΔE<2表示色差只有经验丰富的观察者才能注意到;2<ΔE<3.5表示没有经验的观察者会注意到色差;3.5<ΔE<5表示色差明显;5<ΔE对观察者来说能清晰辨别色差;同下
Note: ΔE<1, color difference is not obvious. 1<ΔE<2, color difference can only be noticed by experienced observers. 2<ΔE<3.5, inexperienced observers can notice the chromatic aberration. 3.5<ΔE<5 indicates obvious color difference. 5<ΔE, the color difference can be clearly distinguished. The same below
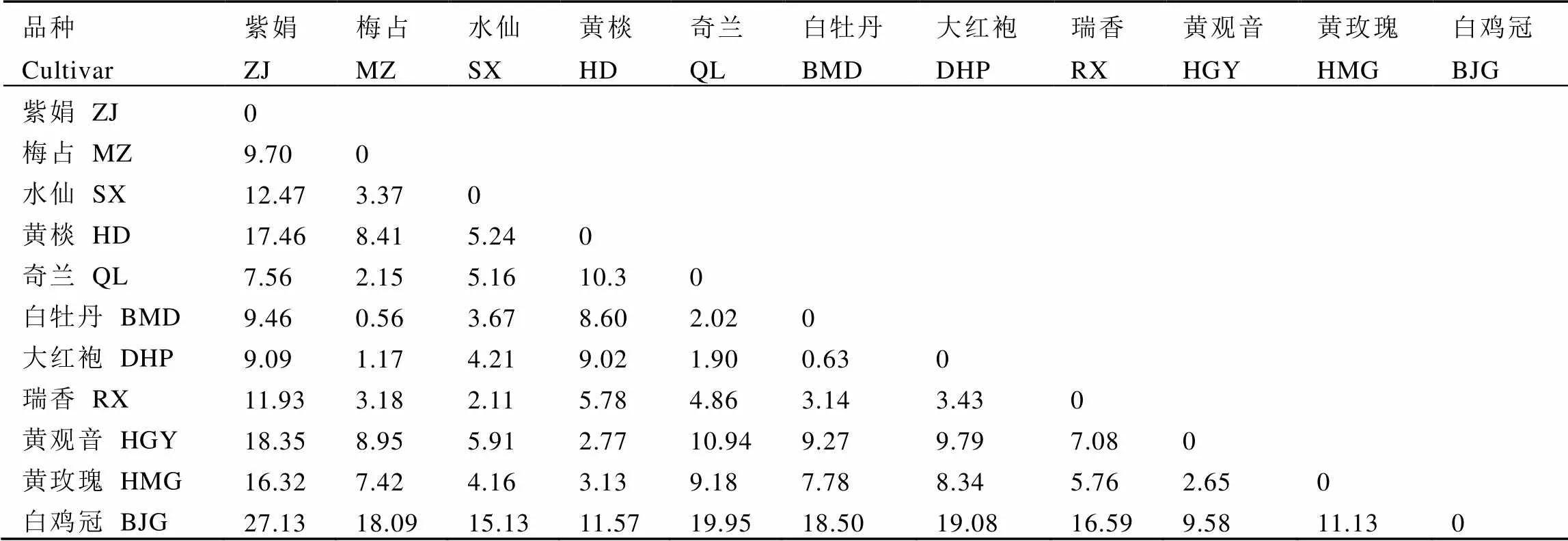
表4 茶树芽下第3叶片叶背色差参数ΔE表
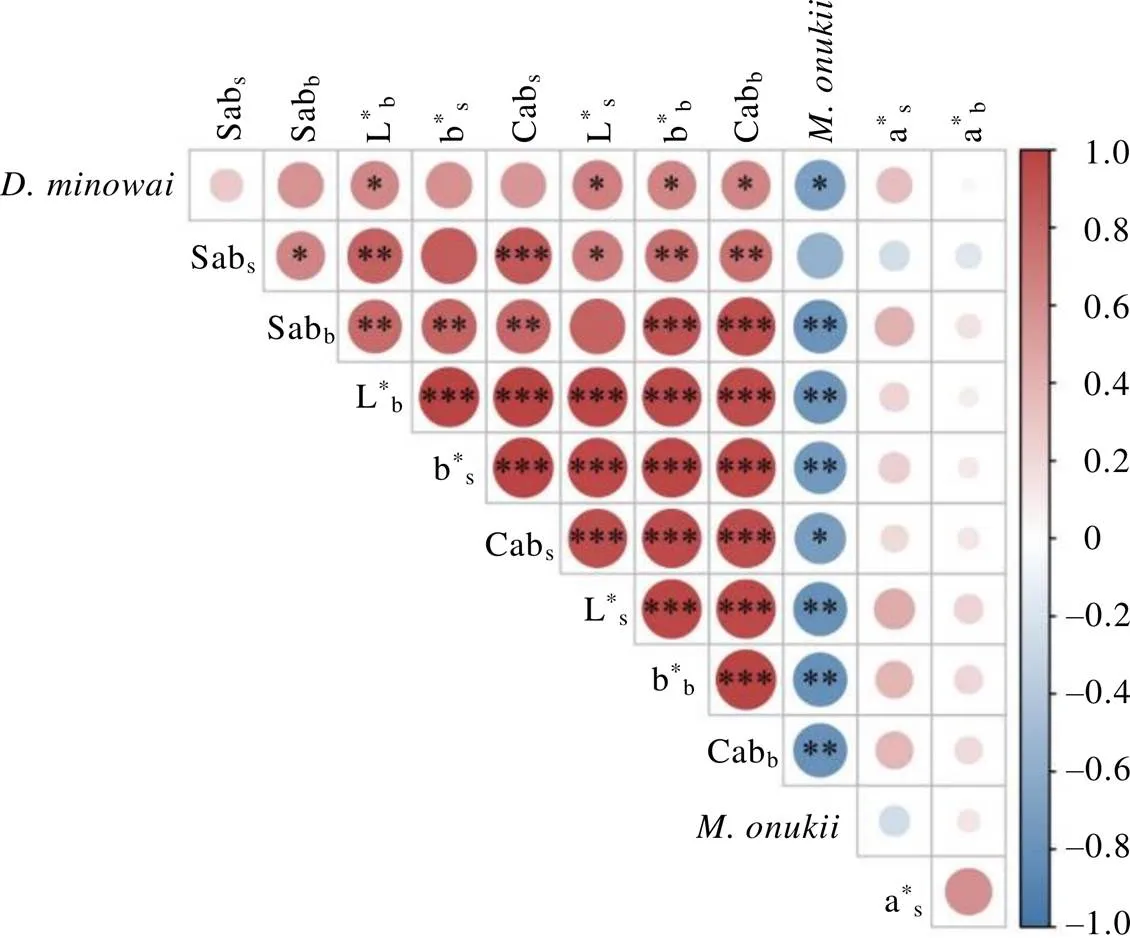
图5 虫口密度与叶片色差的相关性
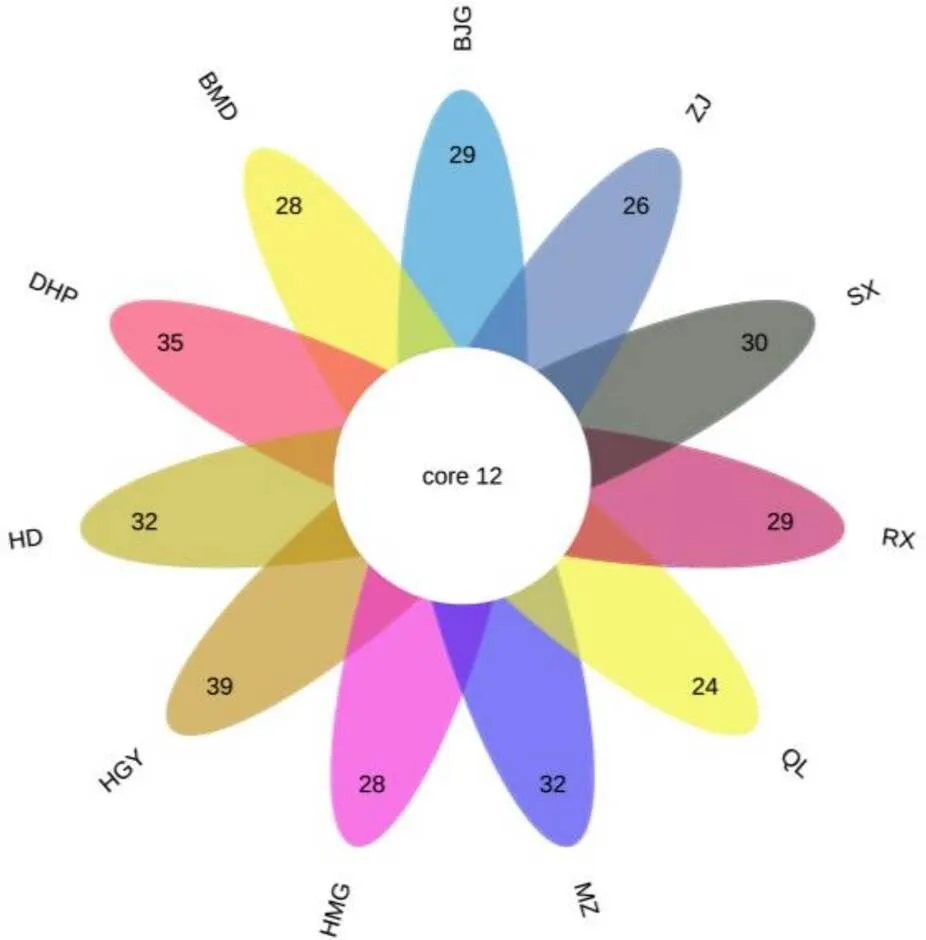
注:数字代表每组的挥发物的种类;core 12表示共有成分12种
表5 11个茶树品种挥发性物质相对含量
Table 5 Relative contents of volatile compounds in 11 tea cultivars
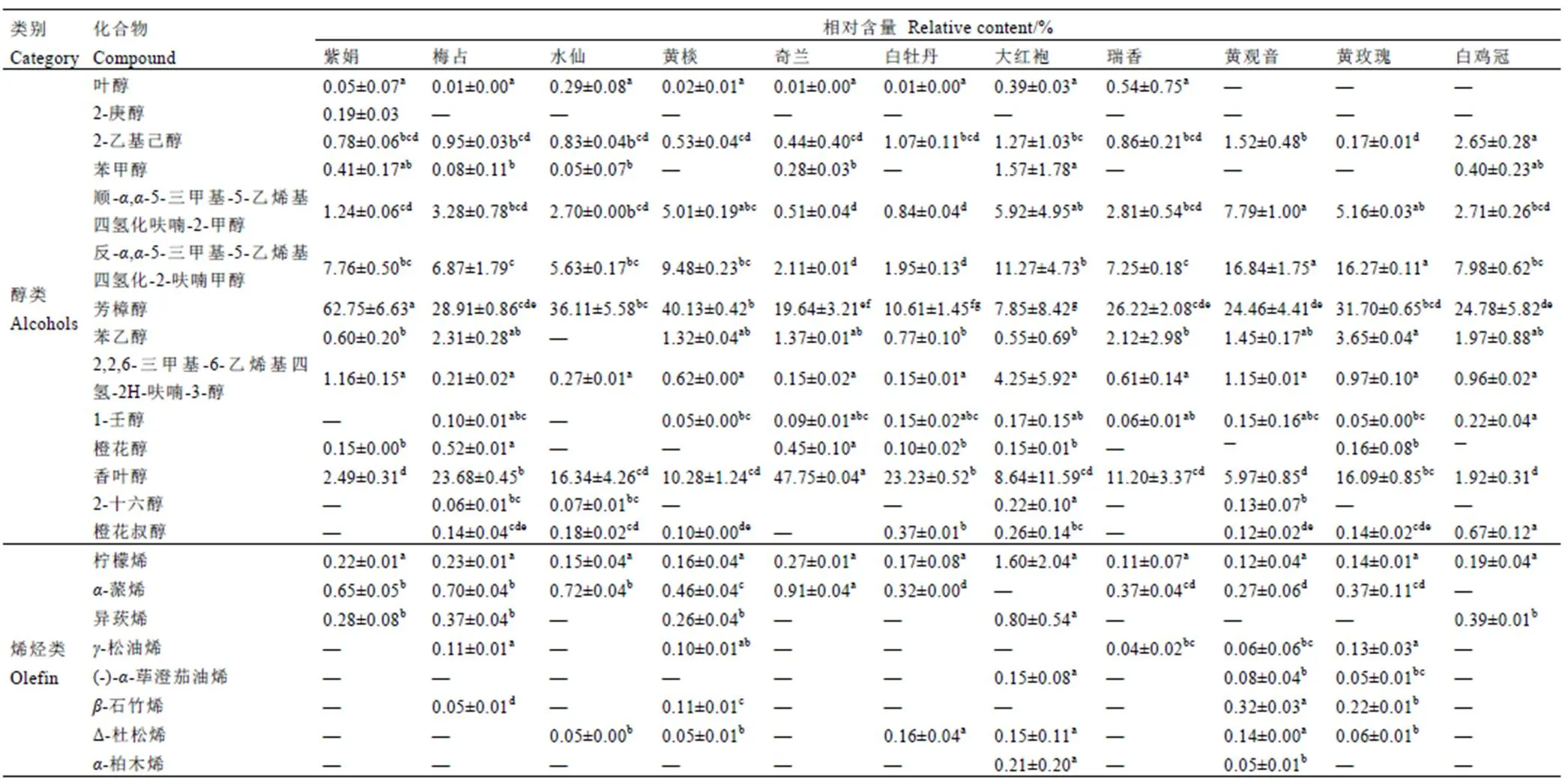
注:同行不同小写字母代表<0.05水平差异显著
Note: Different lowercase letters in the same line represent significant differences at the< 0.05 level
续表5
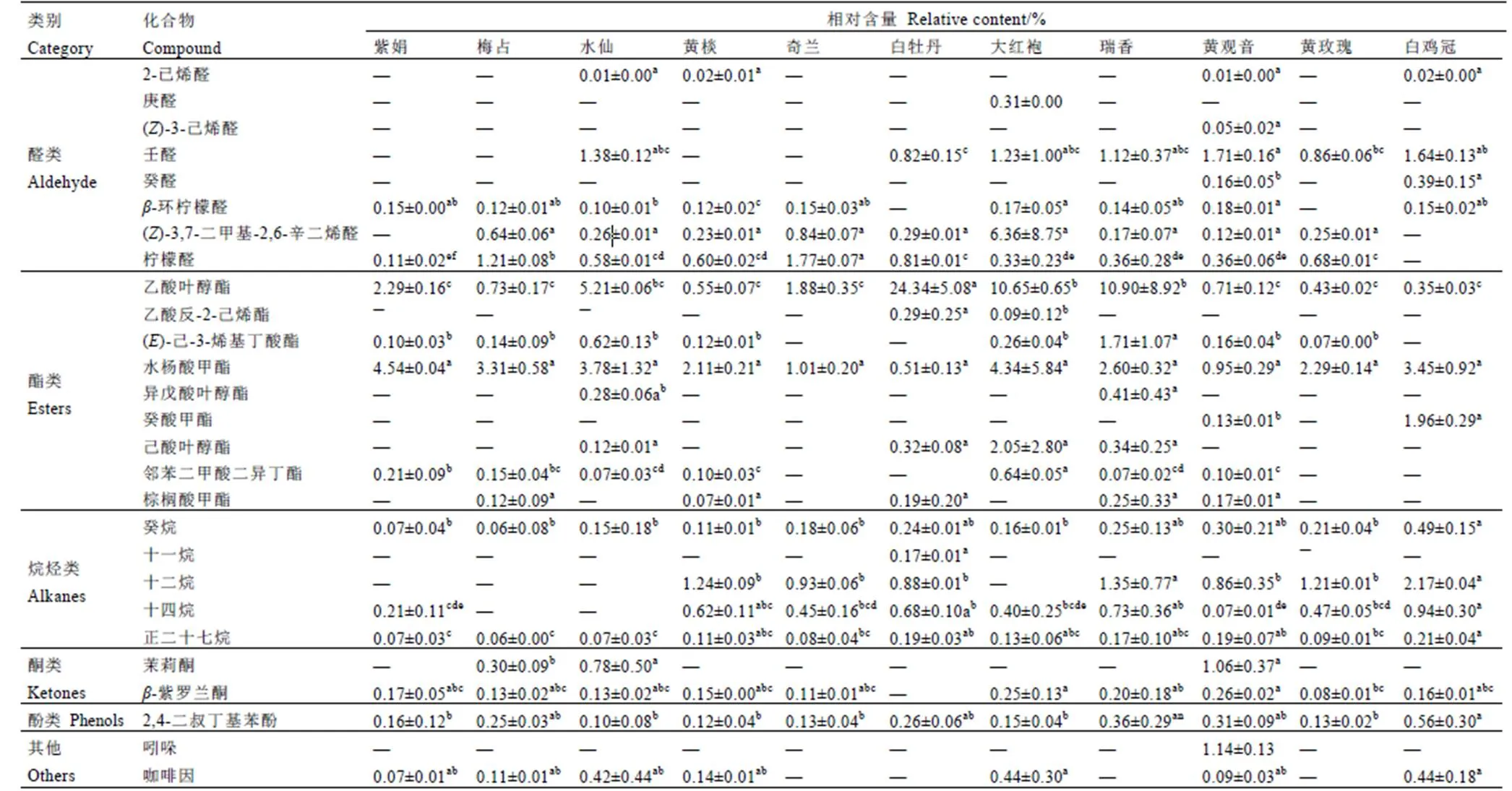
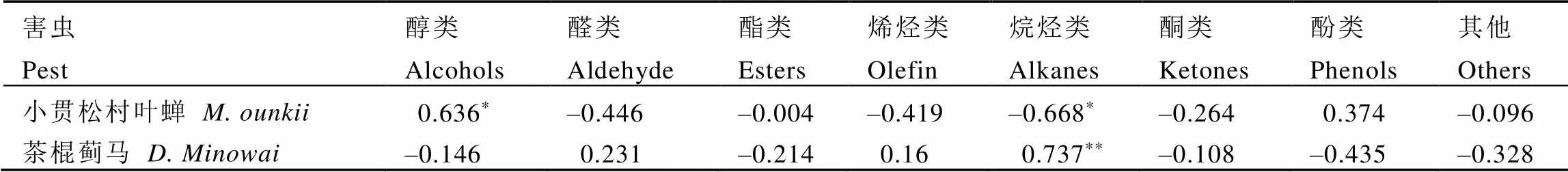
表6 挥发性分类物质与虫口种群密度的相关性
注:*表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关
Note: * means significant correlation at 0.05 level (bilateral). ** means significant correlation at 0.01 level (bilateral)
2.6 不同抗虫性茶树品种挥发物的OPLS-DA分析
基于不同抗虫性级别的茶树品种鲜叶挥发物,构建有监督模式的OPLS-DA模型,小贯松村叶蝉模型的拟合参数为2=0.992,2=0.993,2=0.972;茶棍蓟马模型的拟合参数为2=0.998,2=0.994,2=0.981。设置随机排列200次的置换检验(图8B、8E),2拟合回归直线均与Y轴的截距小于0,斜率为正,表明所建模型均不存在过度拟合现象,模型较为可靠(小贯松村叶蝉:2=0.422,2=–1.29;茶棍蓟马:2=0.418,2=–1.22),能有效对不同抗虫性级别茶树品种的挥发物进行判别分析。
由得分矩阵图可知,OPLS-DA模型中所有点均在95%置信区间内,各组间区分明显。按茶树品种对小贯松村叶蝉的感虫(1)、中感(2)、抗虫(3)等级分组(图8A),感虫品种聚在第1象限,中感品种聚在第2、3象限,抗虫品种聚在第4象限,表明对小贯松村叶蝉不同抗性级别的茶树品种的特征挥发物存在差异。按茶树品种对茶棍蓟马的感虫(1)、中感(2)、中抗(3)、抗虫(4)等级分组(图8D),4组各占一个象限,不同分组的特征挥发物相对含量间存在明显差异,能有效区分不同茶树品种对茶棍蓟马的抗性程度。
图8C和图8F显示,OPLS-DA模型中VIP(Variable importance in projection)>1的特征挥发物分别有7个(小贯松村叶蝉)与9个(茶棍蓟马),这些特征挥发物可能是引起不同茶树品种抗虫性差异的关键性物质。
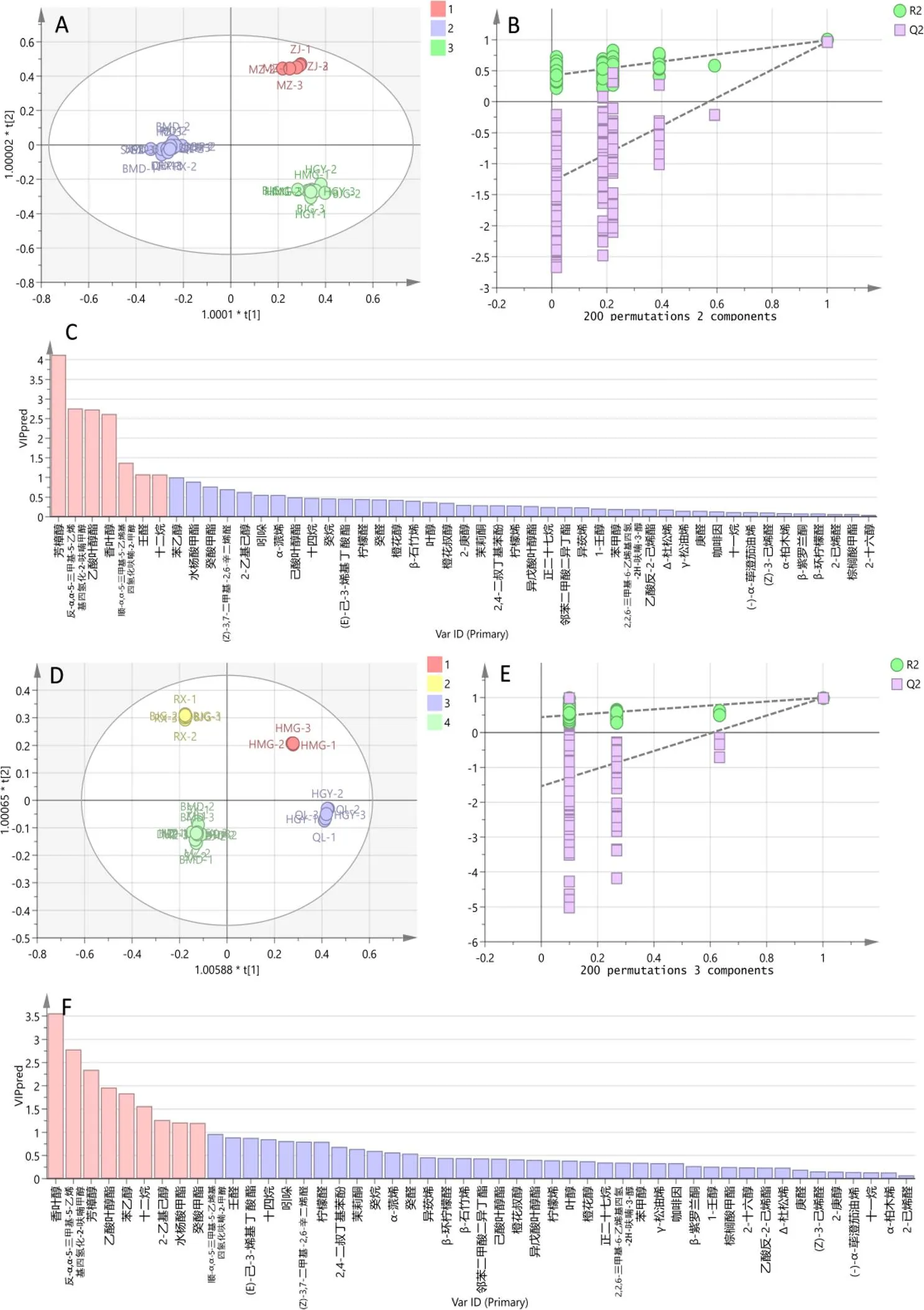
注:A、B、C分别为不同抗小贯松村叶蝉级别茶树品种挥发物的OPLS-DA得分图、置换检验图及变量重要因子;D、E、F分别为不同抗茶棍蓟马级别茶树品种挥发物的OPLS-DA得分图、置换检验图及变量重要因子
Fig. 8 OPLS-DA analysis of volatile organic compounds in different insect-resistant tea cultivars
2.7 特征挥发物与抗虫性的相关性分析
VIP值显示,芳樟醇、反-,-5-三甲基-5-乙烯基四氢化-2-呋喃甲醇、乙酸叶醇酯、香叶醇、顺-,-5-三甲基-5-乙烯基四氢化呋喃-2-甲醇、壬醛、十二烷是界定11个茶树品种感抗小贯松村叶蝉级别的特征挥发物;香叶醇、反-,-5-三甲基-5-乙烯基四氢化-2-呋喃甲醇、芳樟醇、乙酸叶醇酯、苯乙醇、十二烷、2-乙基己醇、水杨酸甲酯、癸酸甲酯是区分11个茶树品种感抗茶棍蓟马级别的特征挥发物;有4种挥发物共同参与了茶树对两种刺吸式害虫的抗性界定。
将种群密度与特征挥发物进行pearson相关性分析(图9),结果显示,小贯松村叶蝉种群密度与芳樟醇相对含量呈显著正相关(<0.05,=0.67);与壬醛、十二烷相对含量呈显著负相关(<0.05,1=–0.66,2=–0.68),说明芳樟醇可能对小贯松村叶蝉具有引诱活性,壬醛、十二烷对小贯松村叶蝉具有趋避或毒害作用。茶棍蓟马种群密度与十二烷和苯乙醇呈极显著正相关(<0.01,1=0.79,2=0.78),表明十二烷和苯乙醇对茶棍蓟马可能存在引诱作用。十二烷对小贯松村叶蝉和茶棍蓟马的作用不同,且小贯松村叶蝉和茶棍蓟马的种群密度呈显著负相关,推测十二烷可能参与影响小贯松村叶蝉和茶棍蓟马之间的生态位竞争。
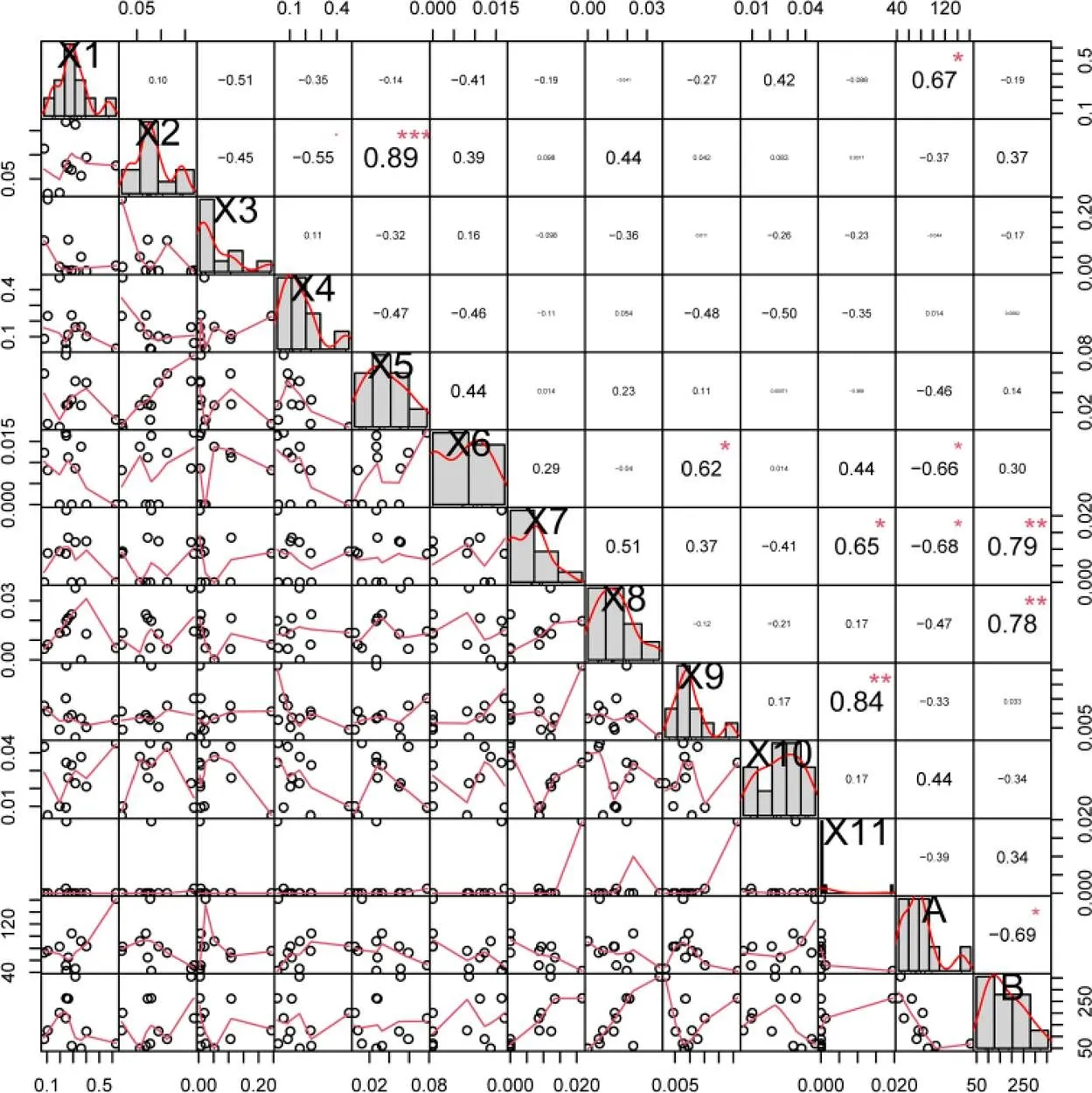
注:X1为芳樟醇,X2为反-α,α-5-三甲基-5-乙烯基四氢化-2-呋喃甲醇,X3为乙酸叶醇酯,X4为香叶醇,X5为顺-α,α-5-三甲基-5-乙烯基四氢化呋喃-2-甲醇,X6为壬醛,X7为十二烷,X8为苯乙醇,X9为2-乙基己醇,X10为水杨酸甲酯,X11为癸酸甲酯,A表示小贯松村叶蝉2年虫口密度均值,B表示茶棍蓟马2年虫口密度均值
2.8 不同抗虫性茶树品种单位叶面积蜡质含量差异分析
为探究叶片蜡质含量是否参与影响茶树抵御两种刺吸式害虫的为害,对不同茶树品种进行单位叶面积蜡质含量的测定,结果如图10所示。11个茶树品种的单位叶面积蜡质含量在0.19~1.09 mg·cm-2,其中,白鸡冠单位叶面积蜡质含量最高,大红袍最低。不同抗虫级别的茶树品种间存在部分显著差异,单位叶面积蜡质含量与小贯松村叶蝉的种群密度呈负相关,与茶棍蓟马种群密度呈正相关(=0.376),但均不显著。烷烃类挥发物主要来自叶片蜡质层,十二烷含量与单位叶面积蜡质含量呈现的结果一致。
3 讨论
种群发生数量是害虫抗性鉴定关键的判别指标,田间种群密度是最直观的呈现方式[19]。本研究对11个茶树品种进行为期2年的连续性调查,将各品种对小贯松村叶蝉的抗性聚为3级(感虫、中感、抗虫),对茶棍蓟马抗性聚为4级(感虫、中感、中抗、抗虫)。各品种的抗性差异显著,是茶树物理抗性[20-21]、化学抗性[22-23]、诱导抗性[24-25]共同作用的结果。本研究以连续2年的田间种群密度为抗性分级标准,利用田间枝条室内孵化试验和室内生命参数测定验证田间种群密度作为抗虫性分级标准的可靠性。结果显示,枝条孵化的茶棍蓟马数量连续5 d均与茶棍蓟马种群密度呈正相关,小贯松村叶蝉的室内生命参数与种群密度划分结果呈现一致性,表明两种刺吸式害虫以田间种群密度为抗性分级标准具有可靠性,能有效反映抗虫性程度。
3.1 茶树叶片色差对两种刺吸式害虫的抗性影响
昆虫寄主定位决定其食性和生态位,昆虫在远距离寄主定位前,视觉占主导作用[26]。刺吸式昆虫均具单眼和复眼,可对寄主植物的颜色、形状、大小等进行识别[27]。本研究发现,茶棍蓟马更倾向叶色黄、色调彩度高及色彩饱和度强的茶树品种,与林金丽等[28]和Bian等[29]研究结果一致。本研究结果显示,小贯松村叶蝉更倾向选择叶色偏深、色调彩度低及色彩饱和度弱的茶树品种。小贯松村叶蝉和茶棍蓟马被多种颜色所吸引,但更趋向选择黄绿或草绿品种[30]。寄主植物主要为绿色与黄绿色之间的差异时,昆虫更倾向于黄绿色,黄绿色比其他颜色更为显眼,目标群体更清晰,更易引诱害虫,更易感虫,B型烟粉虱[31]也有相同趋向性。在本研究结果中,茶棍蓟马更倾向黄绿色,小贯松村叶蝉更倾向绿色,其原因可能是茶棍蓟马更具种群优势所造成的。如Bian等[29]筛选验证茶棍蓟马和茶黄蓟马更受吸引的颜色。结果发现,在同一生境下,茶黄蓟马倾向草绿色,茶棍蓟马更倾向于黄绿色。而王琛等[32]仅单独对茶黄蓟马趋色性进行试验,发现室内的茶黄蓟马嗜好颜色为黄绿色,显著高于对其他颜色的偏好。乔利等[33]单独测试小贯小绿叶蝉成虫的趋光行为,发现小贯小绿叶蝉成虫对黄光(590~595 nm)趋向性相对较强,其次为紫光(420~425 nm)。叶火香等[34]在多种植食性昆虫的存在下进行大田试验,发现小贯松村叶蝉强烈趋向于芽绿和素馨黄色板,芽绿色板诱捕的虫口数显著高于素馨黄色板,这说明种群优势参与影响害虫的视觉寄主定位。
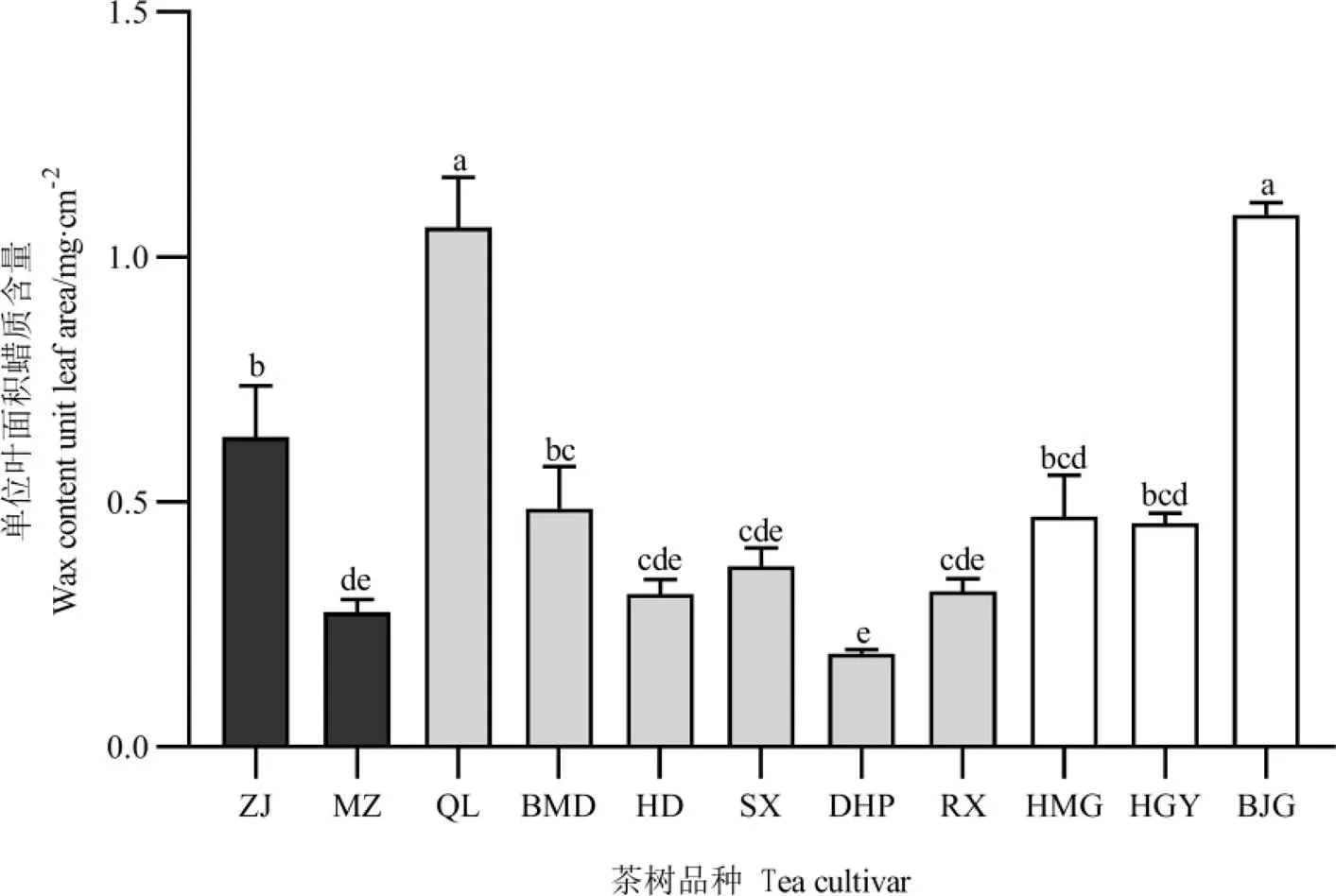
图10 11个茶树品种的单位叶面积蜡质含量
3.2 茶树特征挥发物对两种刺吸式害虫寄主选择的影响
茶树叶片挥发物在很大程度上由芳樟醇、香叶醇等单萜烯醇类化合物和苯甲醇、苯乙醇等芳香族化合物以及青叶醇等不饱和脂肪族化合物决定。脂肪酸过氧化降解生成青叶醇、己烯醛等不饱和脂肪族醇、醛类化合物[35],这些挥发性有机化合物通过被动扩散、囊泡或专门的质膜转运体(PT)从植物中释放到空气中,成为了草食动物捕食性天敌的引诱剂[36]。本研究发现,芳樟醇对小贯松村叶蝉具有引诱活性,壬醛、十二烷对小贯松村叶蝉具有趋避或毒害作用。韩宝瑜[37]和赵冬香等[38]试验均证实芳樟醇对小贯松村叶蝉具有引诱活性。金珊等[39]和任倩倩等[40]的结果也表明芳樟醇对小贯松村叶蝉具有强烈引诱作用。张辉等[41]研究发现,被茶跗线螨不同程度为害的福云6号挥发物中均检测到了壬醛。壬醛具有强烈的油脂气息和柑橘气味,而十二烷来自于叶片蜡质层[42],这可能是茶树能够抵御小贯松村叶蝉为害的部分原因。
本研究还发现,苯乙醇可能对茶棍蓟马存在引诱作用。苯乙醇具有清甜的玫瑰花香,Teulon等[43]利用该特点研究发现苯乙醇能显著提高对烟蓟马的引诱效果。十二烷对茶棍蓟马的作用与小贯松村叶蝉不同,刘莹等[44]发现十二烷能显著驱避圆柏大痣小蜂雄成虫,而陈旭鹏[45]利用白榆树干挥发物中的十二烷和柠檬烯等物质制成的引诱剂对榆木蠹蛾的引诱效果明显,这说明同种挥发物对不同的昆虫会产生不同的趋向作用。本研究通过直接采集茶树鲜叶收集挥发物,而非活体采集,可能存在部分挥发物在茶树活体状态下不释放,本研究将最终筛选的差异特征挥发物与前人的研究比对,发现壬醛、十二烷、芳樟醇、苯乙醇均有在茶树活体鲜叶挥发物中释放[39,46]。
本研究通过对不同抗小贯松村叶蝉和茶棍蓟马级别的茶树种质资源进行叶片色差测定与挥发物鉴定,初步发现小贯松村叶蝉趋向于叶片色彩较暗、黄色较淡的茶树品种;茶棍蓟马趋向色彩鲜亮、饱和度高、黄色偏高的茶树品种。芳樟醇、壬醛、十二烷是小贯松村叶蝉寄主定位的关键特征挥发物,而苯乙醇和十二烷是影响茶棍蓟马寄主定位的关键特征挥发物。今后将进一步利用室内试验验证该结果,找寻小贯松村叶蝉和茶棍蓟马对寄主植物色差选择的原因,并筛选关键组分或混合组分的最佳浓度,为小贯松村叶蝉和茶棍蓟马的生态调控、种群监测及开发高效的植物源引诱剂提供可靠的理论依据。
[1] 任倩倩. 抗、感茶树品种对茶小绿叶蝉取食诱导的防御反应[D]. 福州: 福建农林大学, 2020. Ren Q Q. The defense strategies of resistant and susceptible tea cultivars in response tofeeding [D]. Fuzhou: Fujian Agriculture and Forestry University, 2020.
[2] Chen L L, Yuan P, Pozsgai G, et al. The impact of cover crops on the predatory mite(Acari, Anystidae) and the leafhopper pest(Hemiptera, Cicadellidae) in a tea plantation [J]. Pest Management Science, 2019, 75(12): 3371-3380.
[3] 唐美君, 肖强. 茶树病虫及天敌图谱[M]. 北京: 中国农业出版社, 2018. Tang M J, Xiao Q. Colored pitiorial handbook of tea plant pests and natural enemies [M]. Beijing: China Agriculture Press, 2018.
[4] Affeldt H A, Thimijan R W, Smith F F, et al. Response of the green-house whitefly (Homoptera: Aleyrodidae) and the vegetable leafminer (Diptera: Afromyzidae) to photospectra [J]. Journal of Economic Entomology, 1983, 76(6): 1405-1409.
[5] Snoeren T A L, Peter W, Jong D E, et al. Ecogenomic approach to the role of herbivore-induced plant volatiles in community ecology [J]. Journal of Ecology, 2007, 95(1): 17-26.
[6] Kessler A, Baldwin I T. Defensive function of herbivore-induced plant volatile emissions in nature [J]. Science, 2001, 291(5511): 2141-2144.
[7] Shulaev V, Silverman P, Raskin I. Airborne signalling by methyl salicylate in plant pathogen resistance [J]. Nature, 1997, 385(6618): 718-721.
[8] 蔡晓明. 三种茶树害虫诱导茶树挥发物的释放规律[D]. 北京: 中国农业科学院, 2009. Cai X M. The emission of tea plant volatiles induced by three herbivore insect pests [D]. Beijing: Chinese Academy of Agricultural Sciences, 2009.
[9] 钮羽群, 王梦馨, 崔林, 等. 迷迭香挥发物不同组合对假眼小绿叶蝉行为的调控[J]. 生态学报, 2015, 35(7): 2380-2387. Niu Y Q, Wang M X, Cui L, et al. Control of the tea green leafhopper via volatile compounds of rosemary: the potential for further development of a Push-Pull strategy [J]. Acta Ecologica Sinica, 2015, 35(7): 2380-2387.
[10] 王梦馨, 李辉仙, 武文竹, 等. 假眼小绿叶蝉对茶梢挥发物的行为反应[J]. 应用昆虫学报, 2016, 53(3): 507-515. Wang M X, Li H X, Wu W Z, et al. Behavioral responses ofGöthe to volatiles from tea shoots [J]. Chinese Journal of Applied Entomology, 2016, 53(3): 507-515.
[11] 王蔚, 吴满容, 张思校, 等. 茶小绿叶蝉在福建省茶树品种上的选择机制初探[J]. 河南农业科学, 2016, 45(4): 80-84. Wang W, Wu M R, Zhang S X, et al. Preliminary study on selection mechanism ofon tea varieties in Fujian Province [J]. Journal of Henan Agricultural Sciences, 2016, 45(4): 80-84.
[12] 金珊. 不同茶树品种抗假眼小绿叶蝉机理研究[D]. 咸阳: 西北农林科技大学, 2012. Jin S. Resistance mechanisms of tea plant cultivars to tea green leafhopper [D]. Xianyang: Northwest A&F University, 2012.
[13] 李红莉, 崔宏春, 黄海涛, 等. 联苯菊酯等4种杀虫剂对茶小贯小绿叶蝉的田间防效[J]. 浙江农业科学, 2021, 62(3): 565-566. Li H L, Cui H C, Huang H T, et al. Field control effect of 4 insecticides such as bifenthrin on[J]. Journal of Zhejiang Agricultural Sciences, 2021, 62(3): 565-566.
[14] 施龙清. 福建茶小绿叶蝉种类鉴定及其对光的行为响应[D]. 福州: 福建农林大学, 2016. Shi L Q. Identification of the tea green leafhopper in Fujian, China, and its behavioral responses to varying light conditions [D]. Fuzhou: Fujian Agriculture and Forestry University, 2016.
[15] 刘勇, 陈巨莲, 程登发. 不同小麦品种(系)叶片表面蜡质对两种麦蚜取食的影响[J]. 应用生态学报, 2007(8): 1785-1788. Liu Y, Cheng J L, Cheng D F. Effects of wheat leaf surface waxes on the feeding of two wheat aphid species [J]. Chinese Journal of Applied Ecology, 2007(8): 1785-1788.
[16] 吴少玲, 王治会, 商虎, 等. 不同堆青时间对白茶风味品质的影响[J]. 茶叶科学, 2023, 43(1): 78-90. Wu S L, Wang Z H, Shang H, et al. Effect of pile-up processing duration on flavor quality of white tea [J]. Journal of Tea Science, 2023, 43(1): 78-90.
[17] 石碧滢, 周承哲, 田采云, 等. 不同贮藏时间白牡丹茶风味品质差异分析[J/OL]. 食品科学, 2022: 1-23 [2023-01-12]. http://kns.cnki.net/k cms/detail/11.2206.TS.220221125.1537.012.html. Shi B Y, Zhou C Z, Tian C Y, et al. Analysis of flavor quality differences between white peony tea with different storage time [J/OL]. Food Science, 2022: 1-23 [2022-12-11]. http://kns.cnki.net/kcms/detail/11.2206.TS. 20221125. 1537. 012.html.
[18] 孙悦, 吴俊, 韦朝领, 等. 抗小贯松村叶蝉和茶棍蓟马的茶树种质筛选及其抗性相关因素分析[J]. 茶叶科学, 2022, 42(5): 689-704. Sun Y, Wu J, Wei C L, et al. Screening of tea germplasm resistant toandPriesner and analysis of resistance-related factors [J]. Journal of Tea Science, 2022, 42(5):689-704.
[19] 杨春, 孟泽洪, 李帅, 等. 贵州茶园茶棍蓟马和小贯小绿叶蝉种群动态及主栽茶树品种寄主抗性[J]. 南方农业学报, 2021, 52(3): 671-681. Yang C, Meng Z H, Li S, et al. Population dynamics ofPriesner andMatsuda and host resistance of major tea varieties in Guizhou [J]. Journal of Southern Agriculture, 2021, 52(3): 671-681.
[20] Sun Q Y, Chen W L, Ge C M, et al. Correlation of tea green leafhopper occurrence with leaf structure and biochemical components in different tea cultivars [J]. International Journal of Pest Management, 2023, 69(2): 109-119.
[21] 刘丰静, 冉伟, 李喜旺, 等. 小贯小绿叶蝉在5个茶树品种(系)上的蜜露排泄量与茶树叶片结构比较[J]. 茶叶科学, 2020, 40(5): 625-631. Liu F J, Ran W, Li X W, et al. The comparison of leaf structures andhoneydew excretion among five tea cultivars (strains) [J]. Journal of Tea Science, 2020, 40(5): 625-631.
[22] 金珊, 孙晓玲, 张新忠, 等. 8个茶树品种生化成分分析及抗性成分的初步鉴定[J]. 应用昆虫学报, 2016, 53(3): 516-527.
Jin S, Sun X L, Zhang X Z, et al. Chemical analysis of 8 tea cultivars with different levels of resistance toGöthe and a preliminary identification of the chemical basis of this resistance [J]. Chinese Journal of Applied Entomology, 2016, 53(3): 516-527.
[23] 郑雨婷, 王梦馨, 崔林, 等. 基于EPG技术分析茶树品种对茶小绿叶蝉的抗性及其相关的抗性物质[J]. 生态学报, 2017, 37(23): 8015-8028. Zheng Y T, Wang M X, Cui L, et al. Resistance of tea cultivars to the tea green leafhopper analyzed by EPG technique and their resistance-related substances [J]. Acta Ecologica Sinica, 2017, 37(23): 8015-8028.
[24] Liao Y Y, Yu Z M, Liu X Y, et al. Effect of major tea insect attack on formation of quality-related nonvolatile specialized metabolites in tea () leaves [J]. Journal of Agricultural and Food Chemistry, 2019, 67(24): 6716-6724.
[25] Wang M X, Han S J, Wu Y Q, et al. Tea green leafhopper-induced synomone attracts the egg parasitoids, mymarids to suppress the leafhopper [J]. Pest Management Science, 2023. doi: 10.1002/ps.7563.
[26] 边磊, 孙晓玲, 高宇, 等. 昆虫光趋性机理及其应用进展[J]. 应用昆虫学报, 2012, 49(6): 1677-1686. Bian L, Sun X L, Gao Y, et al. Research on the light tropism of insects and the progress in application [J]. Chinese Journal of Applied Entomology, 2012, 49(6): 1677-1686.
[27] Knolhoff L M, Heckel D G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects [J]. Annual Review of Entomology, 2014, 59(1): 263-278.
[28] 林金丽, 韩宝瑜, 周孝贵, 等. 色彩对茶园昆虫的引诱力[J]. 生态学报, 2009, 29(8): 4303-4316. Lin J L, Han B Y, Zhou X G, et al. The attraction of color to insects in tea garden [J]. Acta Ecologica Sinica, 2009, 29(8): 4303-4316.
[29] Bian L, Yang P X, Yao Y J, et al. Effect of trap color, height, and orientation on the capture of yellow and stick tea thrips (Thysanoptera: Thripidae) and nontarget insects in tea gardens [J]. Journal of Economic Entomology, 2016, 109(3): 1241-1248.
[30] Mohammad H A T, Nazrul I M, Zhang Y, et al. Leaf morphological characters can be a factor for intra-varietal preference of whitefly(Hemiptera: Aleyrodidae) among eggplant varieties [J]. Plos One, 2016, 11(4): e0153880. doi: 10.1371/journal.pone.0153880.
[31] 曹凤勤, 刘万学, 万方浩, 等. 寄主挥发物、叶色在B型烟粉虱寄主选择中的作用[J]. 应用昆虫学报, 2008, 45(3): 431-436. Cao F Q, Liu W X, Wan F H, et al. Behavior selection ofB-biotype to different host plants and colors [J]. Chinese Journal of Applied Entomology, 2008, 45(3): 431-436.
[32] 王琛, 朱文静, 符悦冠, 等. 茶黄蓟马嗜好颜色筛选及监测效果测定[J]. 环境昆虫学报, 2015, 37(1): 107-115. Wang C, Zhu W J, Fu Y G, et al. Screening of preferred color and field evalution for monitoringHood (Thysanoptera: Thripidae) [J]. Journal of Environmental Entomology, 2015, 37(1): 107-115.
[33] 乔利, 金银利, 蒋月丽, 等. 光谱对小贯小绿叶蝉趋光行为的影响[J]. 环境昆虫学报, 2022, 44(5): 1197-1204. Qiao L, Jin Y L, Jiang Y L, et al. Effects of spectra on phototactic behavior of theMatsuda [J]. Journal of Environmental Entomology, 2022, 44(5): 1197-1204.
[34] 叶火香, 韩善捷, 韩宝瑜. 假眼小绿叶蝉趋色性及茶梢信息物质强化其趋色性的效应[J]. 安徽农业大学学报, 2015, 42(6): 868-872. Ye H X, Han S J, Han B Y. Phototaxis of tea green leafhopper and strengthening effect of infochemicals from tea shoots on its phototaxis [J]. Journal of Anhui Agricultural University, 2015, 42(6): 868-872.
[35] Zeng L T, Watanabe N, Yang Z Y. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea () to safely and effectively improve tea aroma [J]. Critical Reviews in Food Science and Nutrition, 2019, 59(14): 2321-2334.
[36] 张冬桃, 孙君, 叶乃兴, 等. 茶树萜烯类香气物质合成相关酶研究进展[J]. 茶叶学报, 2015, 56(2): 68-79. Zhang D T, Sun J, Ye N X, et al. Research progress of enzymes associated with terpene synthesis in[J]. Acta Tea Sinica, 2015, 56(2): 68-79.
[37] 韩宝瑜. 茶树-茶蚜-捕食、寄生性天敌间定位、取食的物理、化学通讯机制[D]. 北京: 中国农业科学院, 1999. Han B Y. The physical and chemical communication mechanism of tea plant-tea aphid-predator and parasitoid positioning and feeding [D]. Beijing: Chinese Academy of Agricultural Sciences, 1999.
[38] 赵冬香, 高景林, 陈宗懋, 等. 假眼小绿叶蝉对茶树挥发物的定向行为反应[J]. 华南农业大学学报(自然科学版), 2002, 23(4): 27-29. Zhao D X, Gao J L, Cheng Z M, et al. Orientation response ofto tea shoots volatiles [J]. Journal of South China Agricultural University (Natural Science Edition), 2002, 23(4): 27-29.
[39] 金珊, 韩李伟, 叶乃兴, 等. 茶小绿叶蝉危害乌龙茶茶树品种的挥发物分析[J]. 热带作物学报, 2019, 40(3): 576-582. Jin S, Han L W, Ye N X, et al. Analysis on volatiles of Oolong tea varieties induced bysp.[J]. Chinese Journal of Tropical Crops, 2019, 40(3): 576-582.
[40] 任倩倩, 庄明珠, 蔡晓明, 等. 小贯小绿叶蝉取食诱导抗、感茶树品种挥发物的释放[J]. 茶叶科学, 2020, 40(6): 795-806. Ren Q Q, Zhuang M Z, Cai X M, et al. The release of volatiles in resistant and susceptible tea cultivars underfeeding [J]. Journal of Tea Science, 2020, 40(6): 795-806.
[41] 张辉, 李慧玲, 王定锋, 等. 茶跗线螨为害程度对福云6号鲜叶芳香物质的影响[J]. 福建农业学报, 2015, 30(9): 896-900. Zhang H, Li H L, Wang D F, et al. Aromatics in fresh leaves of Fuyun 6 () understress [J]. Fujian Journal of Agricultural Sciences, 2015, 30(9): 896-900.
[42] 李晓维, 程江辉, 韩海斌, 等. 植物次生代谢物质对蓟马的行为调控作用及其在蓟马防控中的应用[J]. 昆虫学报, 2022, 65(9): 1222-1246. Li X W, Cheng J H, Han H B, et al. Behavioral manipulation of the plant secondary metabolites to thrips and their application in thrips management [J]. Acta Entomologica Sinica, 2022, 65(9): 1222-1246.
[43] Teulon D A J, Davidson M M, Hedderley D I, et al. 4-pyridyl carbonyl and related compounds as thrips lures: effectiveness for onion thrips and new zealand flower thrips in field experiments [J]. Journal of Agricultural and Food Chemistry, 2007, 55(15): 6198-6205.
[44] 刘莹, 吕东, 陈立, 等. 圆柏大痣小蜂对祁连圆柏挥发物的触角电位和行为反应[J]. 生态学报, 2023, 43(9): 3828-3839. Liu Y, Lü D, Chen L, et al. Electroant-ennagram and responses ofto volatiles of[J]. Acta Ecologica Sinica, 2023, 43(9): 3828-3839.
[45] 陈旭鹏. 榆木蠹蛾三种寄主树干挥发物分析及植源性引诱剂的筛选[D]. 晋中: 山西农业大学, 2016. Cheng X P. Identification of volatiles from three host plants attractive forWalker and the development of plant volatile attractant [D]. Jinzhong: Shanxi Agricultural University, 2016.
[46] 孙廷哲, 岂泽华, 梁可欣, 等. 蚜害茶树挥发物组分变化的聚类分析[J]. 植物学报, 2021, 56(4): 422-432. Sun T Z, Qi Z H, Liang K X, et al. Clustering analysis of volatile components from the tea plants infested by tea aphid () [J]. Chinese Bulletin of Botany, 2021, 56(4): 422-432.
Study on the Differences of Leaf Color and Volatiles of Different Insect-resistance Tea Cultivars
SUN Yue1,3, LIU Mengyue2,3, GAO Chenxi1,3, WU Quanjin4, CAO Shixian5, YU Shuntian5, CHEN Zhidan2,3, JIN Shan1*, SUN Weijiang1,3*
1. College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 2. Anxi College of Tea, Fujian Agriculture and Forestry University, Quanzhou 362400, China; 3. Ministerial and Provincial Joint Innovation Centre for Safety Production of Cross-Strait Crops, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 4. College of Law, Chinese Studies & Economics, the Open University of Fujian, Fuzhou 350003, China; 5. Wuyi Star Tea Industrial Company Limited, Wuyishan 354301, China
In order to explore the differences in leaf color and volatiles of different insect-resistant tea cultivars, a two-year field population density survey was conducted on 11 tea cultivars. Through the indoor incubation test of field branches and the indoor life parameter determination test, the reliability of field population density as the resistance grading standard was proved. The color difference and wax content per unit leaf area of leaves were determined, and the volatiles of new shoots of one bud and two leaves that were not harmed by pests and diseases were detected by gas chromatography-mass spectrometry (GC-MS) in order to screen the differences among different insect-resistant tea cultivars. The results show that,preferred tea cultivars with darker, lighter yellow leaves.preferred tea cultivars with brighter, more saturated colors and a higher yellow color, andplaced more emphasis on the color of the leaf back. In terms of volatiles, the population densities ofwere significantly and positively correlated with the relative content of linalool and negatively correlated with the relative contents of nonanal and dodecane. Population densities ofwere significantly and positively correlated with the relative levels of dodecane and phenylethanol. The effects of dodecane onandwere opposite, and the same trend was observed for the relationship between wax and population density in tea leaves, suggesting that the same volatile substance may have different reactions to different insects.
tea plant,,, leaf color, volatile components, resistance
S571.1;S326
A
1000-369X(2023)04-525-19
2023-02-28
2023-05-15
中国白茶研究院开放课题项目(BCY2021K0100)、福建农林大学“双一流”建设科技创新能力提升培育计划项目(KSYLC005)、福鼎市现代国家农业(白茶)产业园白茶种质资源保护提升项目(KH220095A)、福建张天福茶叶发展基金会科技创新基金(FJZTF01)
孙悦,女,硕士研究生,主要从事茶树栽培育种与生物技术方面研究。*通信作者:jinshan0313@163.com;swj8103@126.com
猜你喜欢
今日农业(2021年11期)2021-11-27 10:47:17
今日农业(2021年15期)2021-11-26 03:30:27
乡村地理(2018年2期)2018-09-19 06:44:00
山地农业生物学报(2018年2期)2018-06-22 12:42:06
现代园艺(2017年21期)2018-01-03 06:42:17
四川动物(2017年1期)2017-02-18 07:05:13
湖南农业(2016年3期)2016-06-05 09:37:36
音乐天地(音乐创作版)(2016年11期)2016-02-05 01:10:59
蚕桑茶叶通讯(2015年1期)2015-02-22 15:13:21
新疆农垦科技(2014年9期)2014-02-28 19:20:58
