
8个菠萝品种的叶片解剖和生态化学计量
2023-08-14 00:07潘艳菊冯莹欧阳有勤陈晓慧叶倚歆蔡志全
热带作物学报 2023年7期
潘艳菊 冯莹 欧阳有勤 陈晓慧 叶倚歆 蔡志全



摘 要:以江门市同质园11 个月生的8 个菠萝品种(金钻菠萝、红皮菠萝、红香菠萝、手撕菠萝、金菠萝、巴厘菠萝、西瓜菠萝、芒果菠萝)为研究对象,探讨植株的生长、叶片气孔、解剖和元素(C、N、P、K)生态化学计量特征,拟揭示不同菠萝品种功能性状的差异,叶片元素含量和生态化学计量与植株生长的关系,以及生长的营养限制。研究结果表明:在菠萝成熟叶片的上、中、下不同部位中,多数品种中部的气孔密度最大;叶片厚度从上部到下部基本呈增大的趋势。且不同叶片部位或同一叶片部位菠萝品种间的气孔密度、保卫细胞长度、叶片总厚度和各组织解剖结构比例大小存在显著差异。不同菠萝品种间叶片C、N、P、K 含量差异显著。菠萝对K 的需求较高,因为其叶片K 含量远远高于其N 和P 含量。菠萝叶片N 和P 含量远低于中国草地植物叶片N 和P 含量,但其C 含量、C∶N 和C∶P 值高于全国草地植物和农田生态系统中植被的叶片平均水平。这表明菠萝吸收单位营养物质所能同化C 的能力较高;可能是通过特殊的叶片形态或组织结构减少对营养元素的依赖,从而采取“减少需求策略”。8 个菠萝品种间叶片C 含量、C∶P 和N∶P 与植株的叶片总生物量呈显著负相关,符合生长速率假说。叶片N 和P 元素化学计量(N∶P)的初步研究结果表明,金菠萝和芒果菠萝受P 限制,金钻菠萝、红皮菠萝、手撕菠萝和西瓜菠萝受N 限制,而红香菠萝和巴厘菠萝受N 和P 的共同限制。该研究结果可为菠萝优质品种的选择和农业管理提供理论参考。
关键词:菠萝;品种;气孔;解剖;矿质元素;生态化学计量
中图分类号:S668.3 文献标识码:A
叶片是植物对环境最敏感且可塑性较大的器官,其解剖结构特征能够体现叶片对特定环境的适应[1]。如温度较高地区,植物叶片通常具有小而密的气孔,较薄的叶片厚度、栅栏组织和海绵组织厚度[2]。同时,叶片解剖结构对植物的光合作用和水分功能也有重要影响[3]。光合作用主要在栅栏组织中进行,叶片对光的吸收取决于其解剖结构。叶片组织结构通过反射和折射影响光路径,光从叶片的表皮进入,且上表皮吸收光线的表面积大,能够将更多的光线聚集[4]。叶片的栅栏组织排列紧密,其柱状结构为光线通过提供了通道;随着叶片深度的增加,光强度变弱,海绵组织不规则形状增强了光的反射和折射,增加了穿过叶片的光程长度,从而增加了光吸收[5]。此外,叶片海绵组织细胞间隙发达,可以储存水分;栅栏组织和海绵组织影响着植物与大气环境之间的碳水交换[6]。而气孔负责了水分和气体在叶片和大气之间的流通,对于维持叶片光合作用和水分储存具有重要作用[7]。
碳(C)、氮(N)、磷(P)是影响植物生长的重要营养元素,存在强烈的耦合关系;其生态化学计量主要研究植物器官内元素含量及其比值特征、环境因子和植物个体生长发育之间的关系,涵盖了从分子到生态系统等不同尺度能量及化学元素的平衡[8]。生长速率假说和N∶P 的营养限制理论是生态化学计量学发展的2 个重要理论[9]。
生长速率假说认为,随生长速率增加,植物体N∶P、C∶N 和C∶P 呈降低趋势,而P 含量呈增加趋势。这是因为N、P 是蛋白质和核酸的重要组成成分,较高的生长速率需要大量的蛋白质和核酸来维持[9-10]。N∶P 的营养限制理论认为,N∶P作为限制性营养元素的指标,可以反映植物受土壤氮或磷限制情况[11-12]。因此,植物C、N 和P化学计量比在一定程度上可以表示植物的C 积累量和N、P 元素养分限制,同时也反映潜在的植物生理生化和对环境变化潜在的适应性[11]。地理变异和种间变异是造成植物叶片化学计量差异的主要原因[13]。目前,野生植物物种水平上特定植物器官植物性状的大型数据库已经建立[14]。由于农田生态系统的物种单一和土壤微生物在群落水平上的化学计量可塑性较低,农田生态系统植被的生态化学计量相对于森林生态系统而言具有较强的保守性[15-16]。但对作物的生态化学计量研究不多。如对不同类型甘薯的研究发现,叶中C∶N、C∶P 分别以紫甘薯和淀粉型甘薯最大。表明这2种甘薯有较高的碳同化能力;且叶的N∶P 值(8.57~9.97)表明甘薯生长可能受到土壤N 元素的限制[17]。有整合分析表明,谷物、豆类和油料三类作物间的P 和N∶P 具有显著差异;而P 是作为N∶P 变化的主要因素,N 与N∶P 仅在豆类作物内显著相关[18]。但目前尚无景天酸代谢途径(CAM)作物的生态化学计量研究。
热带水果菠萝[Ananas comrosus (L.) Mer.]是典型的CAM 作物。以同质园内的8 个菠萝品种为实验材料,研究了植株大小、叶片解剖和主要元素(C、N、P 和K)含量及其化学计量特征。
同质园实验通过将来自不同遗传材料的植物个体放在相同的环境中生长,可以最大程度地降低环境条件造成的影响,进而说明不同植物材料的性狀表现受基因型的影响程度。本研究拟初步回答以下3 个科学问题:(1)不同菠萝品种叶片解剖结构、元素含量及其生态化学计量有无显著的差异?(2)不同菠萝品种叶片元素含量和生态化学计量是否能预测植株的生长(生物量)?(3)不同菠萝品种间的土壤养分元素限制是否有显著差异?结果以期为不同菠萝品种功能性状的研究及其农田管理措施提供相关的科学理论依据。
1 材料与方法
1.1 材料
试验材料8 个菠萝品种来源于中国热带农业科学院南亚热带作物研究所,分别为:金钻菠萝(JZ)、红皮菠萝(HP)、红香菠萝(HX)、手撕菠萝(SS)、金菠萝(J)、巴厘菠萝(BL)、西瓜菠萝(XG)、芒果菠萝(MG),每个品种的吸芽(约30 cm)于2020 年10 月按株行距35 cm35cm 栽植于江门市司前镇大田实验基地,地理位置112°51E,22°29N,海拔500 m。该地属亚热带季风性气候,全年四季分明,气候温和,热量充足,雨量充沛,年均气温21.8 ℃,年均降水量1773.8 mm。菠萝各品种生长期间管理水平一致,每个菠萝品种的施肥量为:N 497 kg/hm2,P2O5396 kg/hm2,K2O 6772 kg/hm2;其中基施占60%,追施占40%。追施在种后6 个月后开沟施入20%,其余20%采用撒施方式。
1.2 方法
1.2.1 样品采集 于2021 年9 月,每个品种随机选择长势一致且健康的菠萝植株4~5 棵,选取健壮的植株上中部完整成熟叶片5~6 片。
1.2.2 叶片和植株生长特征 成熟单叶面积的测定采用方格法,将叶片的轮廓描在标准计算纸(最小方格的规格为1 mm1 mm)上,统计叶轮廓占的小方格数(达到或超过半格的算1 格,不足半格的舍去),得到叶片面积。每品种4 次重复。
叶片称取鲜重后放入烘箱105 ℃杀青1 h,在80 ℃下烘干至恒重后称取干重,以计算各叶片样本的含水量:叶片含水量=(叶片鲜重-叶片干重)/叶片鲜重×100%。
比叶面积(SLA)=叶片面积/叶片干重。
整株叶片总生物量:将植株完整叶片放入烘干箱中,在105 ℃下杀青30 min,80 ℃條件下烘干至恒重,通过整株总叶片数估计叶片面积,估算出整株叶片总生物量。
株高及冠幅(植株南北和东西2 个方向宽度的平均值)用直尺进行测量。
1.2.3 叶片气孔和解剖特征 新鲜叶片不同部位(上、中、下部)的选择:以叶片基部为起点,5~15 cm 片段为下部,25~35 cm 片段为中部,45~50 cm 片段为上部。
气孔密度和大小:分别用指甲油涂抹在叶片背面上、中、下部约2 cm2 区域内,等待3~5 min指甲油干后,用镊子撕下印记干膜,制成装片。
置光学显微镜(Nikon)40 倍镜下随机拍摄3 个视野。将微尺放于同一倍镜下拍摄,并利用ImageJ 软件对图片进行测量分析,每品种4 次重复,得出气孔密度和保卫细胞长度。
叶片解剖:在叶片上、中、下部沿垂直于叶脉的方向剪取1 cm1 cm 的正方形叶块,进行徒手切片制成装片。将装片置于光学显微镜10 倍镜下随机拍摄3 个视野。将微尺放于同一倍镜下拍摄,利用Image J 软件对切片图各结构厚度进行测量和分析,得出叶片的上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度和总厚度。并分别计算其占总厚度的比例。
1.2.4 叶片元素测定 全碳含量:植物全碳光度法;全氮含量:凯氏定氮法;全磷含量:钼锑抗比色法;全钾含量:火焰光度法。每个元素重复3~4 次。
1.3 数据处理
变异系数(CV)是描述变量变异程度的指标,为标准差与平均值之比,计算公式为:CV=标准差/平均值×100%。
采用SPSS 25.0 软件进行平均值间比较的单因素方差(One-way ANOVA)、双因素方差(TwowayANOVA)及多重比较分析。利用Pearson 相关系数分析相关关系,采用Graphpad Prism 8.0软件制图。
2 结果与分析
2.1 叶片和植株生长特征
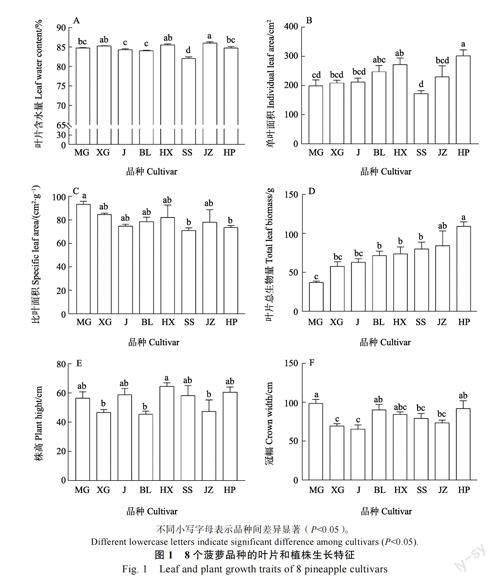
8 个菠萝品种间的成熟叶片含水量(F=21.89,P<0.001)、单叶面积(F=3.96,P=0.005)、冠幅(F=3.85,P=0.006)和叶片总生物量(F=5.59,P=0.001)均具有显著差异,但株高(F=2.42,P>0.05)和比叶面积(F=1.58,P>0.05)无显著差异(图1A~图1F)。其中,叶片含水量以金钻菠萝最大,单叶面积和叶片总生物量以红皮菠萝最大,冠幅以芒果菠萝最大。叶片含水量和单叶面积均以手撕菠萝最小,叶片总生物量以芒果菠萝最小,冠幅以金菠萝最小。8 个菠萝品种间,株高和冠幅呈显著正相关(r=0.39,P=0.029),生物量与SLA 呈显著负相关(r=‒0.58,P<0.001)。
叶片总生物量与株高(r=0.28,P>0.05)和冠幅(r=0.13,P>0.05)的相关性不显著。
2.2 叶片气孔和解剖特征
叶片不同部位(上、中、下部)对8 个菠萝品种的气孔密度有显著影响(图2A)。其中,叶片上部的气孔密度相对较高。对于同一部位的叶片,不同菠萝品种的气孔密度具有显著差异(上部F=8.62,中部F=17.73,下部F=8.96;P<0.05)。
在上部叶片中,芒果菠萝气孔密度最大,巴厘菠萝气孔密度最小。而在中、下部叶片中,菠萝品种气孔密度的变化规律不同。叶片不同部位(上、中、下部)对8 个菠萝品种的气孔保卫细胞长度有显著影响(图2B)。其中,叶片下部的保卫细胞长度相对较大。对于同一部位的叶片,不同菠萝品种的气孔保卫细胞长度无显著差异(P>0.05)。在上部叶片气孔中,金钻菠萝保卫细胞长度最大,西瓜菠萝保卫细胞长度最小。
叶片不同部位(上、中、下部)对8 个菠萝品种叶片的总厚度有显著影响。其中,叶片下部的叶片总厚度相对较大(图3A)。对于8 个品种菠萝同一部位的叶片,其叶片总厚度均具有显著差异(上部F=5.79,中部F=1.77,下部F=9.23;P<0.05)。在上部和下部叶片中,总厚度均以红皮菠萝最大;在中部叶片中,总厚度以红香菠萝最大。在上部和中部叶片中,总厚度均以巴厘菠萝最小, 在下部叶片中, 总厚度以金钻菠萝最小。8 个菠萝品种不同部位叶片和同一叶片部位上的上表皮厚度、栅栏组织厚度、海绵组织厚度和下表皮厚度比例均无显著差异(P>0.05)(图3B~图3E)。上表皮厚度比例和栅栏组织厚度比例分别在叶片上部和下部最大,在叶片下部和中部最小。海绵组织厚度比例和下表皮厚度比例均在叶片中部最大,在叶片下部最小。
2.3 叶片元素含量和生态化学计量
不同菠萝品种间叶片的C ( F=3.15 ,P=0.017)、N(F=8.74,P<0.001)、P(F=35.02,P<0.001)、K(F=4.99,P=0.001)、C∶N(F=6.80,P<0.001)、C∶P(F=47.28,P<0.001)和N∶P(F=20.32,P<0.001)具有显著差异(图4)。8个菠萝品种叶片C、N、P、K、C∶N、C∶P 和N∶P 范围分别为46.52%~51.51%、0.58%~0.90%、0.03%~0.10% 、2.12%~3.22% 、50.24~86.85 、512.76~1653.35 和6.40~19.07。N,P 和K 三种宏量元素中,K 含量最高,远远高于N 和P 含量。
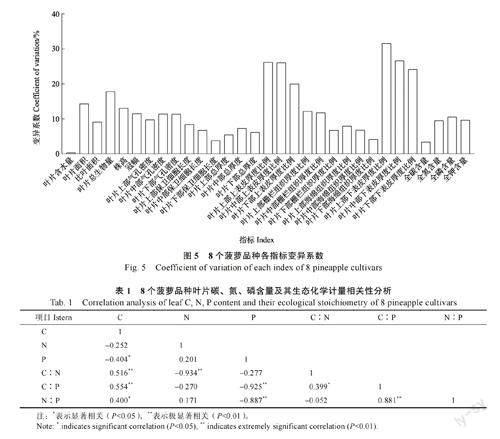
叶片元素变异由大到小依次为P>N>K>C(图5)。
8 个菠萝品种叶片P 含量与N∶P 呈负相关(P<0.05),N 含量与N∶P 相关性不显著(表1)。
2.4 叶片元素含量及生态化学计量与叶片总生物量的关系
8 个菠萝品种叶片C、C∶P、N∶P 与叶片总生物量呈显著负相关(P<0.05),叶片P 与叶片总生物量呈显著正相关(P<0.05),而叶片N、K、C∶N 与叶片总生物量无显著相关关系(图6)。
3 讨论
3.1 叶片和植株生长特征
植物叶片总生物量、株高和冠幅均可在一定程度上反映植物生长的大小。由于菠萝的叶片生物量约为整株生物量的90%以上[19-20],叶片总生物量可表征菠萝的整株生物量。葉片面积、株高及冠幅越大,菠萝的生长状况越好[21]。在本研究中,8 个菠萝品种间,叶片总生物量与株高和冠幅的相关性不显著,因而株高和冠幅不能精确预测菠萝不同品种整株的生物量。此外,8 个菠萝品种间,比叶面积与叶片总生物量呈显著负相关,这与一般的“叶片经济谱”的规律相反。其原因可能是菠萝作为CAM 光合途径的植物,由于其特殊的叶片解剖结构导致叶片构建的投入不能指示其生长(生物量),较小的比叶面积可能与菠萝的抗旱性有关[22]。
3.2 气孔和解剖特征
气孔密度和气孔大小与植物的抗旱性、光合作用有一定关系。通常情况下,当植物面临干旱胁迫时,小气孔可以快速关闭减少蒸腾耗水,水分利用效率高,气孔密度大有利于光合作用且加大散热,增强植物应对碳同化与水分散失的平衡,从而更快地适应环境的变化[2, 23]。通过对芭蕉芋的研究发现,其气孔密度越大,叶片散热和水分散失的能力就越强,从而提高植物水分利用率和抗旱性,同时提高光合速率[24]。但也有研究结果显示,随着干旱程度的加剧,气孔密度不是一直升高,而是先升高后下降。糜子气孔密度小,降低蒸腾,提高植物的含水量,叶片散失水分途径减少,提高了光合效率,最终表现出较强的抗旱性[25]。在本研究中,不同菠萝品种叶片部位解剖组织存在差异,大部分品种叶片中部的气孔密度最大,而少量品种的气孔密度分别在上部和下部叶片最大。所以,在采样的时候需考虑对不同叶片部位的选择。植物叶片气孔特性对周围环境变化的响应是一个非常复杂的生理过程,仅以气孔密度和气孔大小作为影响植株蒸腾作用的指标来衡量其保水性和抗旱性还不够全面,必须结合其他指标综合考虑。因而,不同品种菠萝的气孔特性是否表征其抗旱性有待进一步研究。
一般情况下,发达的栅栏组织可增强叶片光合作用和水分运输速率,减少叶子脱水造成的伤害。紧密的海绵组织可控制减少气体交换,有利于提高植物抗旱性[26]。通过对12 个核桃品种的研究发现,植物通过增加表皮厚度能够减少水分的蒸发,从而增强叶片的保水能力和抗旱性[27]。
张小燕等[7]对红树植物的研究发现,叶片厚度越厚,植物光合速率越大。因为较厚的叶片具有发达的储水组织,叶片内盐分浓度得以稀释,保证叶片在蒸腾情况下维持足够的水分进行光合作用。厚的叶片扩大了栅栏组织和海绵组织的暴露面积,增强了叶片对CO2 的吸收能力,从而避免了CO2 对叶片光合速率的限制。另外,对玉米不同部位叶片的研究表明,下部叶片的光合速率最低[28]。本研究发现,8 个菠萝品种叶片各部位厚度从上部到下部基本呈依次增大的趋势,且各组织解剖结构厚度比例存在差异。因此,菠萝不同部位的叶片可能采取了不同的对策,即增加其厚度以获得单位叶面积内含有更多的光合细胞,保证其进行光合作用。此外,不同菠萝品种间上表皮厚度比例和下表皮厚度比例的变异系数最大,而叶片总厚度的变异系数较小。这说明菠萝上下表皮厚度的比例受遗传因子影响的程度较大,而叶片总厚度具有较强的遗传保守性。
3.3 叶片元素含量及其生态化学计量
本研究中,8 个菠萝品种叶片C 含量(平均值为48.39%)与ELSER 等[29]研究的全球492 种陆地植物叶片平均值(46.40%)相近,但高于全国草地植物叶片平均水平(43.80%)[30]。在热带、亚热带光照、降水充足的环境条件下,菠萝的光合作用和新陈代谢得以充分进行,其营养器官得到快速构建,从而可能导致了叶片中C 的大量积累[31]。菠萝叶片中高的C 含量导致其C∶N∶P的平均值(896∶13∶1)高于陆地生态系统植物叶的C∶N∶P 比值(427∶19∶1)[32]和我国10个农田生态系统中植被的C:N:P(740∶15∶1)[15]。
N 和P 是植物体内最主要的2 个宏量元素,与植物的生长密切相关[9]。与中国草地植物叶片N 和P 含量(平均值分别为2.06%和0.16%)[33]相比,8 个菠萝品种叶片N 和P 含量(平均值分别为0.71%和0.06%)很低。因而,菠萝叶片C∶N(70.30)和C∶P(895.91)值远高于全国草地植物(C∶N=17.9,C∶P=273.9)[13, 30]和我国10 个农田生态系统中植被的相应值(C∶N=51.3,C∶P=740.1)[15]。较高的叶片C∶N 和C∶P 值表明吸收单位营养物质所能同化C的能力较高。因此,菠萝可能是通过特殊的形态或组织结构减少对营养元素的依赖,从而采取“减少需求策略”[34]。
与其他的研究结果一致,菠萝品种叶片K 含量远远高于N 和P 含量[35];表明K 是菠萝叶片的第一元素,对钾的需求量大。这可能是菠萝需要较多的K+来调节气孔的运动,维持细胞的膨胀率和相对含水量,降低渗透势;同时能够通过调节干旱条件下许多生理生化过程与代谢平衡之间的关系,从而提高植物抵御抗旱能力[36-37]。
通常情况下,植物叶片N 和P 含量高,其光合速率和生长速率较快,对生存所需资源的获取能力强;而叶片C 含量高,其光合速率和生长速率较慢[38-39]。本研究中,菠萝叶片C 含量、C∶P和N∶P 与叶片生物量呈显著的负相关关系,P含量与叶片生物量呈显著的正相关关系。生物量低和生长较慢的菠萝品种增加对叶片的C 分配,叶片C∶P 较高;而生物量高和生长较快的菠萝品种减少对叶片的C 分配,P 元素含量较高,叶片C∶P 和N∶P 减小。菠萝品种间,叶片元素的变异性大小顺序表现为:C<N<P。
同时,各菠萝品种间叶片P 含量与N∶P 呈显著负相关,而N 与N∶P 之间不存在显著相关性,这说明P 含量是影响N∶P 的重要因素[18]。
另外,P 是核酸的重要组成部分,植物在生长过程中需要积累大量的核酸满足自身生长的需求。生长速率假说认为,生长速率高的植物具有较低的N∶P 和C∶P[9, 40]。本研究发现,8 个菠萝品种间叶片C∶P 和N∶P 与叶片生物量呈显著负相关,P 含量与叶片生物量呈显著正相关。这与对毛莨科(Ranunculaceae)冬菟葵(Eranthishyemalis)[40]的研究结论一致,均符合生长速率假说。但在养分限制条件下,N 和P 并不完全被用于植物的生长代谢活动,在植物组织中存在奢侈吸收的情形,使得N、P 与生长速率的关系变得含糊。虽然有研究认为生长速率假说的普遍适用性需要考虑养分限制类型[41]。但本研究证明了在施肥条件下,生长速率假说在某些作物(菠萝)中的实用性[29]。
叶片N 和P 的计量特征被广泛应用于植物受N、P 限制的情况,也可以作为评估环境条件对植物生长养分供应的指标。一般认为,当N∶P<14时,植物更容易被认为受到N 限制;当N∶P>16时,植物更容易被认为受到P 限制;当14<N:P<16时,其中,金钻菠萝、红皮菠萝、手撕菠萝和西瓜菠萝叶片N∶P<14,其生长可能受到了土壤中N 元素的限制。金菠萝和芒果菠萝叶片N∶P>16,其生长可能受到了土壤中P 元素的限制。红香菠萝和巴厘菠萝叶片14<N:P<16.
总之,本研究表明不同菠萝品种叶片解剖结构、元素含量及其生态化学计量有显著的差异。
不同菠萝品种的叶片C 和P 含量以及C∶P 和N∶P 能预测植株的生长(生物量);后者符合生长速率假说。叶片N∶P 的营养限制初步表明,不同菠萝品种对土壤N、P元素的限制程度具有显著分异。
在实际生产中,需要依据具体菠萝品种的特点,优化其施肥方案以提高菠萝的生长和产量。今后还需加强菠萝农田生态系统中植物-土壤-土壤微生物的生态化学计量和长期、多因子交互控制实验的研究,为实现菠萝产业的可持续发展提供依据。
致谢 感谢江门市清沁绿农业科技发展有限公司梁志成先生和中国热带农业科学院南亚热带作物研究所欧雄常为本研究提供的帮助。
参考文献
[1] HETHERINGTON A M, WOODWARD F I. The role of stomata in sensing and driving environmental change[J].Nature, 2003, 424(6951): 901-908.
[2] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报, 2005(S1): 118-127.
[3] CARINGELLA M A, BONGERS F J, SACK L. Leaf hydraulic conductance varies with vein anatomy across Arabidopsis thaliana wild-type and leaf vein mutants[J]. Plant,Cell & Environment, 2015, 38(12): 2735-2746.
[4] BRODERSEN C R, VOGELMANN T C. Do epidermal lens cells facilitate the absorptance of diffuse light?[J]. American Journal of Botany, 2007, 94(7): 1061-1066.
[5] DELUCIA E H, NELSON K, VOGELMANN T C, SMITH W K. Contribution of intercellular reflectance to photosynthesis in shade leaves[J]. Plant, Cell & Environment, 1996,19(2): 159-170.
[6] TERASHIMA I, HANBA Y T, THOLEN D, NIINEMETS Ü. Leaf functional anatomy in relation to photosynthesis[J].Plant Physiology, 2011, 155(1): 108-116.
[7] 張小燕, WEE K S A, KAJUTA T, 曹坤芳. 种源地对两种红树叶片结构和功能的影响: 对温度的适应性遗传[J]. 植物生态学报, 2021, 45(11): 1241-1250.
[8] 田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说[J]. 植物生态学报, 2021, 45(7): 682-713.
[9] STERNER R W, ELSER J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere[M]. New Jersey: Princeton University Press, 2002.
[10] ÅGREN G I. The C ∶ N ∶ P stoichiometry of autotrophs-theory and observations[J]. Ecology Letters, 2004,7(3): 185-191.
[11] KOERSELMAN W, MEULEMAN A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33(6): 1441-1450.
[12] GÜSEWELL S. N∶P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004,164(2): 243-266.
[13] HE J S, WANG L, FLYNN D F, WANG X, MA W, FANG J.Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes[J]. Oecologia, 2008, 155(2): 301-310.
[14] TIAN D, KATTGE J, CHEN Y H, HAN W X, LUO Y K,HE J S, HU H F, TANG Z Y, MA S H, YAN Z B, LIN Q H,SCHMID B, FANG J Y. A global database of paired leaf nitrogen and phosphorus concentrations of terrestrial plants[J].Ecology, 2019, 100(9): e02812.
[15] 赵航, 贾彦龙, 王秋凤. 中国地带性森林和农田生态系统C-N-P 化学计量统计特征[J]. 第四纪研究, 2014, 34(4):803-814.
[16] BERTRAND I, VIAUD V, DAUFRESNE T, PELLERIN S,RECOUS S. Stoichiometry constraints challenge the potential of agroecological practices for the soil C storage. A review[J]. Agronomy for Sustainable Development, 2019,39(6): 1-16.
[17] 郭琪琳, 吴海云, 李欢, 刘庆. 不同类型甘薯茎和叶中碳、氮、磷化学计量学特征研究[J]. 作物杂志, 2020, 36(2):41-47.
[18] SADRAS V O. The N∶P stoichiometry of cereal, grain legume and oilseed crops[J]. Field Crops Research, 2006,95(1): 13-29.
[19] 习嘉民, 谭施北, 郑金龙, 贺军虎, 习金根. 菠萝根系生长发育与地上部相关性研究[J]. 热带农业科学, 2021,41(2): 88-92.
[20] 韩忠钰, 金鑫, 邓燕, 赵艳, 吕烈武, 黄顺坚, 李长江. 不同施氮水平对金菠萝苗期生长及养分吸收的影响[J]. 热带作物学报, 2021, 42(4): 1008-1013.
[21] 邓英毅, 屈啸, 李峰, 覃婵婵, 馮斗, 禤维言, 裴铁雄, 姜建初, 郭标, 刘永南. 香蕉不同品种生长发育、结果性状和产量比较[J]. 热带作物学报, 2018, 39(9): 1683-1688.
[22] 张秀芳, 穆振北, 林美娇, 江淼华, 巩嘉欣, 游巍斌. 琅岐岛4 种优势植物叶功能性状及其影响因子[J]. 应用与环境生物学报, 2020, 26(3): 667-673.
[23] FRANKS P J, FARQUHAR G D. The mechanical diversity of stomata and its significance in gas-exchange control[J].Plant Physiology, 2007, 143(1): 78-87.
[24] 赵铅, 周竣宇, 张文娥, 潘学军. 芭蕉芋叶片结构特征及其与抗旱性的关系[J]. 西北植物学报, 2019, 39(11):1970-1978.
[25] 张盼盼, 慕芳, 宋慧, 屈洋, 王鹏科, 冯佰利. 糜子叶片解剖结构与其抗旱性关联研究[J]. 农业机械学报, 2013,44(5): 119-126.
[26] ZHU J, CAI D, WANG J, CAO J, WEN Y, HE J, ZHANG S.Physiological and anatomical changes in two rapeseed (Brassica napus L.) genotypes under drought stress conditions[J]. Oil Crop Science, 2021, 6(2): 97-104.
[27] 白重炎, 陈超, 张颖, 高尚峰. 12 个核桃品种叶片解剖结构比较研究[J]. 江苏农业科学, 2010(4): 129-130.
[28] 王燕鹏, 崔震海, 朱延姝, 樊金娟, 张立军. 玉米C4 光合叶不同部位解剖结构和光抑制特性的比较[J]. 植物生理学报, 2012, 48(6): 571-576.
[29] ELSER J J, FAGAN W F, KERKHOFF A J, SWENSON NG, ENQUIST B J. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change[J]. New Phytologist, 2010, 186(3): 593-608.
[30] HE J S, FANG J, WANG Z, GUO D, FLYNN D F, GENG Z.Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006,149(1): 115-122.
[31] 郑帷婕, 包维楷, 辜彬, 何晓, 冷俐. 陆生高等植物碳含量及其特点[J]. 生态学杂志, 2007(3): 307-313.
[32] ZHANG J, LI M, XU L, ZHU J, DAI G, HE N. C∶N∶Pstoichiometry in terrestrial ecosystems in China[J]. Science of The Total Environment, 2021, 795: 148849.
[33] TIAN D, YAN Z B, MA S H, DING Y H, LUO Y K, CHEN Y H, DU E Z, HAN W X, KOVACS E D, SHEN H H, HU HF, KATTGE J, SCHMID B, FANG J Y. Family-level leaf nitrogen and phosphorus stoichiometry of global terrestrial plants[J]. Science China Life Sciences, 2019, 62(8):1047-1057.
[34] SARDANS J, JANSSENS I A, ALONSO R, VERESOGLOU S D, RILLIG M C, SANDERS T G, CARNICER J,FILELLA I, FARRÉ-ARMENGOL G, PEÑUELAS J. Foliar elemental composition of European forest tree species associated with evolutionary traits and present environmental and competitive conditions[J]. Global Ecology and Biogeography, 2015, 24(2): 240-255.
[35] HANAFI M M, SELAMAT M M, HUSNI M H A, ADZEMI M A. Dry matter and nutrient partitioning of selected pineapple cultivars grown on mineral and tropical peat soils[J]. Communications in Soil Science and Plant Analysis, 2009,40(21/22): 3263-3280.
[36] 魏永勝, 梁宗锁. 钾与提高作物抗旱性的关系[J]. 植物生理学通讯, 2001, 37(6): 576-580.
[37] HASSAN M U, AAMER M, CHATTHA M U, ULLAH M A, SULAMAN S, NAWAZ M, WANG Z Q, Ma Y Q, HUANG G Q. The role of potassium in plants under drought stress: mini review[J]. Journal of Basic and Applied Sciences,2017, 13: 268-271.
[38] POORTER L, BONGERS F. Leaf traits are good predictors of plant performance across 53 rain forest species[J]. Ecology,2006, 87(7): 1733-1743.
[39] SHIPLEY B, LECHOWICZ M J, WRIGHT I, REICH P B.Fundamental trade-offs generating the worldwide leaf economics spectrum[J]. Ecology, 2006, 87(3): 535-541.
[40] NIKLAS K J, COBB E D. N, P, and C stoichiometry of Eranthis hyemalis (Ranunculaceae) and the allometry of plant growth[J]. American Journal of Botany, 2005, 92(8):1256-1263.
[41] YAN Z B, KIM N, HAN W X, GUO Y L, HAN T S, DU E Z, FANG J Y. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana[J]. Plant and Soil, 2015, 388(1): 147-155.
猜你喜欢
金桥(2020年10期)2020-11-26
阅读与作文(小学高年级版)(2019年5期)2019-05-27
好孩子画报(2019年12期)2019-01-13
职工法律天地·下半月(2017年2期)2017-02-20
热带农业科学(2016年10期)2016-12-12
科技视界(2016年21期)2016-10-17
安徽农学通报(2015年10期)2015-06-15
医学信息(2015年6期)2015-03-17
