
miR-145-5p 在正常大鼠与2 型糖尿病大鼠骨髓间充质干细胞成骨分化中的表达比较
2023-08-08 10:20:48戴雅文鄂玲玲马小草刘洪臣解放军总医院研究生院北京00853解放军总医院第一医学中心口腔医学研究所口腔颌面战创伤军队重点实验室北京00853
解放军医学院学报 2023年5期
戴雅文,鄂玲玲,郑 颖,马小草,张 戎,时 权,刘洪臣解放军总医院研究生院,北京 00853;解放军总医院第一医学中心口腔医学研究所、口腔颌面战创伤军队重点实验室,北京 00853
全球超过5 亿人患有糖尿病,其中2 型糖尿病(type 2 diabetes mellitus,T2DM)约占90%[1]。T2DM 可引起一系列并发症,除视网膜病变、心血管疾病、肾病、神经病变等疾病外,还会导致骨丢失和骨质疏松,影响骨愈合,增加骨折的风险。随着糖尿病患者数量增加,糖尿病导致的骨代谢疾病成为全球重要的疾病负担[2-4]。有证据表明,糖尿病微环境可影响糖尿病骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMMSCs)而导致机体骨代谢失衡,但其具体机制尚不清楚[5-6]。
微RNA (microRNA,miRNA) 是一类短的非编码RNA,通过靶向靶基因调控蛋白表达参与细胞增殖、凋亡、迁移等多种生物学过程,与骨代谢和骨稳态密切相关[7]。miR-145-5p 靶向SOX9 抑制正常人脂肪间充质干细胞增殖及成软骨分化[8];靶向Smad4 可降低Sox-9、Col-2A1、聚集蛋白聚糖的表达,抑制骨关节炎患者源骨髓间充质干细胞的增殖和成软骨分化[9]。miR-145-5p 还可靶向SEMA3A 降低Wnt 通路基因Wnt3a、Wnt10a 的表达,抑制2 型糖尿病大鼠脂肪间充质干细胞成骨分化[10]。然而,miR-145-5p 对2 型糖尿病股骨BMMSCs 成骨分化的影响尚不清楚。本研究比较正常大鼠与2 型糖尿病大鼠股骨BMMSCs 的增殖和成骨分化能力,初步探讨正常大鼠与2 型糖尿病大鼠股骨BMMSCs 成骨分化中miR-145-5p、SEMA3A、β-catenin 表达的差异,为进一步研究miRNA-145-5p 在糖尿病环境下影响骨代谢的分子机制提供依据。
材料与方法
1 实验动物 8 周龄SPF 级雄性Wistar 大鼠(共12 只) 购于维通利华有限公司,动物许可证号:SCXK(京)2021-0011。8 周龄雄性GK 大鼠(共12只)购于常州卡文斯实验动物有限公司,动物许可证号:SCXK(苏)2016-0010。动物实验经解放军总医院实验动物伦理委员会审核并予以批准(2021-X17-90)。
2 主要试剂和仪器 罗氏Accu-Check Active 活力型血糖仪(Roche,瑞士),低糖DMEM (Gibco,美国),胎牛血清(Gibco,美国),CCK-8 试剂盒(日本同仁,日本),结晶紫染色液(碧云天,中国),抗坏血酸(Sigma,美国),β-甘油磷酸钠(Sigma,美国),地塞米松(Sigma,美国),茜素红S 染色液(Sigma,美国),OriCell®间充质干细胞(大鼠)表面标记检测试剂盒(Cyagen, 中国),碱性磷酸酶(alkaline phosphatase,ALP) 染色液(偶氮偶联法) (Solarbio,中国),ALP 检测试剂盒(碧云天,中国),Western blot 及IP 细胞裂解液(无抑制剂) (碧云天,中国),BCA 蛋白定量试剂盒(Solarbio,中国),RNA 提取液(Servicebio,中国),ECL 化学发光试剂盒(Servicebio,中国),荧光定量PCR 仪(Bio-Rad,美国)。
3 2 型糖尿病大鼠模型建立 高脂高糖饲料喂养GK 大鼠(12 只) 构建2 型糖尿病大鼠模型,作为实验组;普通饲料喂养Wistar 大鼠(共12 只)作为对照组。8 ~ 13 周每周于固定时间测定体质量、大鼠尾静脉采血进行随机血糖检测。2 次随机血糖浓度≥16.7 mmol/L 视为2 型糖尿病模型造模成功[11]。
4 股骨BMMSCs 原代培养及传代 取正常Wistar大鼠和造模成功的GK 大鼠各12 只,腹腔注射过量戊巴比妥钠处死,分离出双侧股骨,用咬骨钳去除股骨两端,暴露骨髓腔,用5 mL 注射器吸取普通培养液(含10% 胎牛血清和1% 青链霉素的DMEM 培养液)冲洗骨髓腔,收集冲洗液于100 mm大皿中,细胞孵育箱内培养(37 ℃,5% CO2)。当细胞密度达80%融合时进行传代培养,使用第3 代Wistar 大鼠BMMSCs (WT-BMMSCs)和GK 大鼠BMMSCs (GK-BMMSCs)进行后续的实验[12]。
5 干细胞表型检测 取第3 代WT-BMMSCs 和GK-BMMSCs,PBS 洗2 次,0.25% 胰蛋白酶消化,1000 r/min 离心5 min,弃去上清液。各管加入流式细胞缓冲液重悬细胞,使细胞浓度为3 ×106/mL。取100 µL 细胞悬液放入1.5 mL EP 管,每管加入2 µL 一抗CD90、CD44、CD34、CD45、CD11b/c 混匀。4℃孵育30 min,用200 µL 流式缓冲液清洗细胞2 次。250 g 离心5 min,弃上清。每管加入100 µL 流式细胞缓冲液,加入二抗,重悬细胞。4℃孵育30 min,用200 µL 流式缓冲液清洗细胞2 次。250 g 离心5 min,弃上清。用300 µL 流式细胞缓冲液重悬细胞后流式细胞仪分析。
6 CCK-8 检测细胞生长 取第3 代WT-BMMSCs和GK-BMMSCs, 以2 × 103/孔的密度接种于96孔板(普通培养液),每组样本量为6。培养的1 ~9 d,每孔加入10 µL CCK-8 工作液,37℃孵箱孵育2 h 后,于450 nm 处检测两组细胞的吸光度值,连续检测9 d,绘制细胞生长曲线。实验重复3 次。
7 细胞集落形成实验 取第3 代WT-BMMSCs和GK-BMMSCs,胰酶消化后重悬并进行细胞计数。以1 × 102/孔的密度接种于六孔板(普通培养液),每组样本量为6。常规培养10 d,弃去原培养液,4% 多聚甲醛固定30 min,结晶紫染色5 min,PBS 清洗3 遍,拍照。计数每孔的集落数,计算集落形成率。实验重复3 次。
8 成骨诱导实验 实验分为WT-BMMSCs、GKBMMSCs、WT-BMMSCs + 成骨诱导液(osteogenic differentiation medium,OM)和GK-BMMSCs + OM四组。以2 × 105/孔的密度接种于六孔板(普通培养液),每组样本量为6。常规培养至细胞达70%融合后分别更换为普通培养液和成骨诱导液(含10%胎牛血清、1% 青链霉素、10 mmol/L β-甘油磷酸钠、50 µg/mL 抗坏血酸和1 × 10-4mmol /L 地塞米松液的DMEM 培养液) 继续培养。每3 d 换液1 次。
9 ALP 染色及半定量分析 成骨诱导7 d 后弃去诱导液,PBS 洗涤3 次,4% 多聚甲醛固定30 min,配置BCIP/NBT 染色工作液,每孔加入1 mL,室温避光孵育15 min,PBS 洗涤2 次后拍照。半定量分析:弃去原培养液,PBS 清洗2 次,每孔加入100 µL 细胞裂解液充分裂解细胞,收集裂解液于EP 管中,4℃下12000 r/min离心30 min,取上清。BCA 法测定蛋白浓度,将样品稀释为50 µg/50 µL。每孔吸取50 µL 加入96 孔板中,同时设置空白对照组和标准品组,每组设5 个复孔。加入50 µL 显色液,37℃孵育10 min,每孔加入100 µL 终止液终止反应,酶标仪于405 nm 处测定吸光度值,根据标准曲线计算ALP 表达量[13]。实验重复3 次。
10 茜素红染色观察钙化结节 成骨诱导21 d后,PBS 清洗1 次,4% 多聚甲醛固定30 min,1%茜素红染色液染色5 min,PBS 清洗后拍照。
11 实 时 定 量PCR 检 测miR-145-5p、SEMA3A、成骨相关基因和成骨通路关键蛋白的mRNA表达 实验分为WT-BMMSCs、GK-BMMSCs、WT-BMMSCs + OM 和 GK-BMMSCs + OM 四组,将细胞以2 × 105/孔的密度接种于六孔板中培养于普通培养液,每组样本量为3,待细胞达70%融合后分别更换为普通培养液和成骨诱导液继续培养7 d。每孔加入1 mL Trizol,提取细胞总RNA,使用超微量分光光度计测定RNA 浓度及纯度。逆转录合成cDNA 共20 µL 反应体系:5 ×Reaction Buffer 4 µL;Gene-specific primer 2 µL;Servicebio®RT Enzyme Mix 1 µL; RNA 10 µL;RNase free water 3 µL,程 序 设 置 为25℃变 性5 min;42℃退火30 min;85℃延伸5 s。然后配置以下反应体系:2 × qPCR Mix 7.5 µL;基因引物(上游 + 下游) 1.5 µL;cDNA 2 µL;Water Nuclease-Free 4 µL,按 照 以 下 条 件 进行PCR 扩增:95℃预变性30 s;95℃变性15 s;60℃延伸30 s,进行40 个循环,荧光定量PCR 检测各组细胞中miR-145-5p、SEMA3A,成骨相关基因COL-1、RUNX2、ALP、OCN 和成骨通路关键蛋白β-catenin 的mRNA 表达水平。引物信息见表1。ΔΔCT 法处理结果,实验重复3 次,取平均值。

表1 引物序列Tab. 1 Primer sequences
12 Western blot 检 测SEMA3A 和 成 骨 相 关 基因的蛋白表达 实验分为WT-BMMSCs、GKBMMSCs、WT-BMMSCs + OM 和GK-BMMSCs +OM 四组,将细胞以2 × 105/孔的密度接种于六孔板中培养于普通培养液,每组样本量为3,待细胞达70%融合后分别更换为普通培养液和成骨诱导液继续培养7 d。培养结束后用PBS 清洗2 遍,每孔加入RIPA 裂解液200 µL,冰上裂解5 min,4℃下12000 r/min 离心10 min,收集上清,即为总蛋白溶液。BCA 蛋白浓度测定试剂盒测定蛋白浓度。制备5% SDS-PAGE 凝胶电泳进行蛋白分离,PVDF 转膜,室温下5% 脱脂牛奶封闭30 min,加入一抗(一抗稀释比:Actin 1∶2000;ALP 1∶300; OCN 1∶300; RUNX2 1∶300;SEMA3A 1∶300) 4℃孵 育 过 夜;次 日TBST 洗3 次,每次5 min,加入二抗(二抗稀释比 1∶500)室温孵育30 min 后,TBST 洗3 次,ECL 发光液显影检测。使用Adobe 软件对Western blot 条带进行分析。实验重复3 次。
13 统 计 学 分 析 统 计 学 分 析 使 用Graphpad Prism 8 软件进行,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析,Tukey 多重比较检验,P<0.05 为差异有统计学意义。
结 果
1 2 型糖尿病大鼠造模成功 通过4 周高脂高糖饲料的喂养,GK 大鼠出现血糖上升及多饮、多食、多尿现象。12 周时GK 大鼠的体质量显著低于Wistar 大鼠(P<0.001;图1),同时血糖显著高于Wistar 大鼠并维持稳定(P<0.001;图2)。连续两次检测随机血糖≥16.7 mmol/L,GK 大鼠2 型糖尿病模型造模成功。
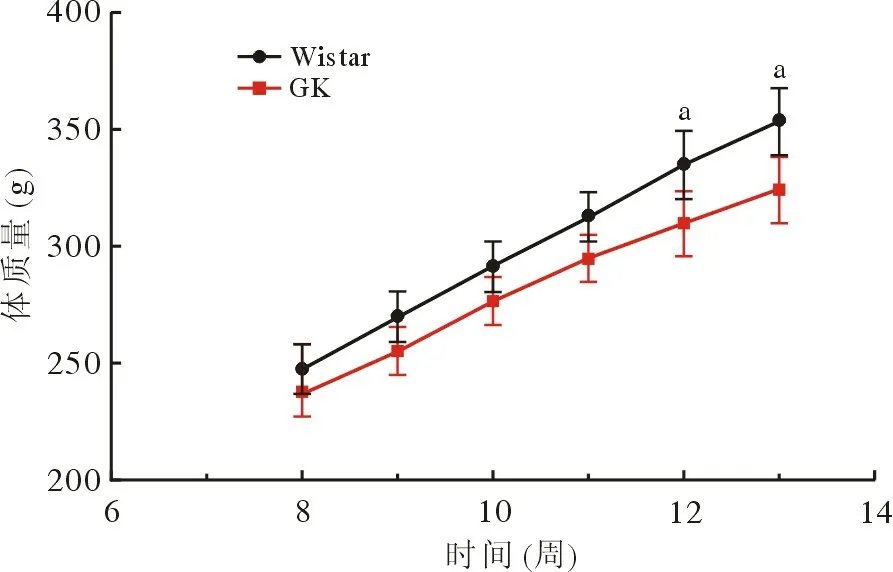
图1 8 ~ 13 周Wistar 与GK 大鼠体质量(n=12;aP<0.001,vsGK)Fig.1 Changes in body weight of Wistar and GK rats from 8 to 13weeks (n=12; aP<0.001, vsGK)
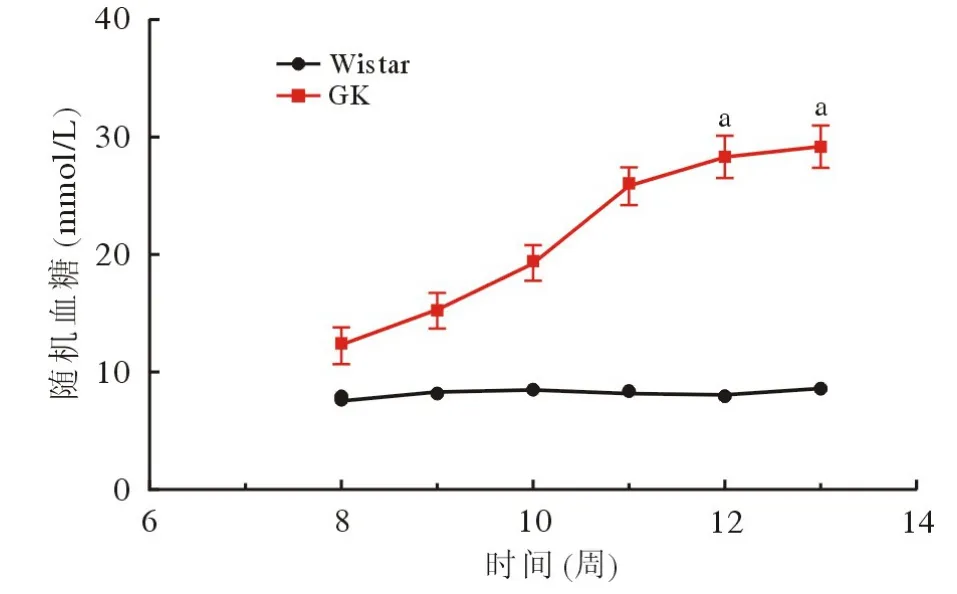
图2 8 ~ 13 周Wistar 与GK 大鼠血糖(n=12;aP<0.001,vsGK)Fig.2 Changes in blood glucose of Wistar and GK rats feom 8 to 13weeks (n=12; aP<0.001, vsGK)
2 股骨BMMSCs 分离培养及干细胞表型检测镜下见原代细胞呈长梭形或多角形,贴壁生长,夹杂着圆形高亮未贴壁的杂细胞,原代细胞培养6 ~ 8 d 可达融合,传代后细胞2 ~ 4 d 可达融合(图3)。流式细胞术进行干细胞表型检测,结果显示,体外培养的WT-BMMSCs 和GK-BMMSCs均阳性表达大鼠间充质干细胞表面标志物CD90(97.5%/99.6%) 、CD44 (98.8%/97.9%),阴性表达造血 干细 胞 标 志 物CD34 (1.49%/2.47%)、CD45(1.13%/1.55%)、CD11b/c (1.51%/1.87%)。符 合 干细胞分子表型(图4)。
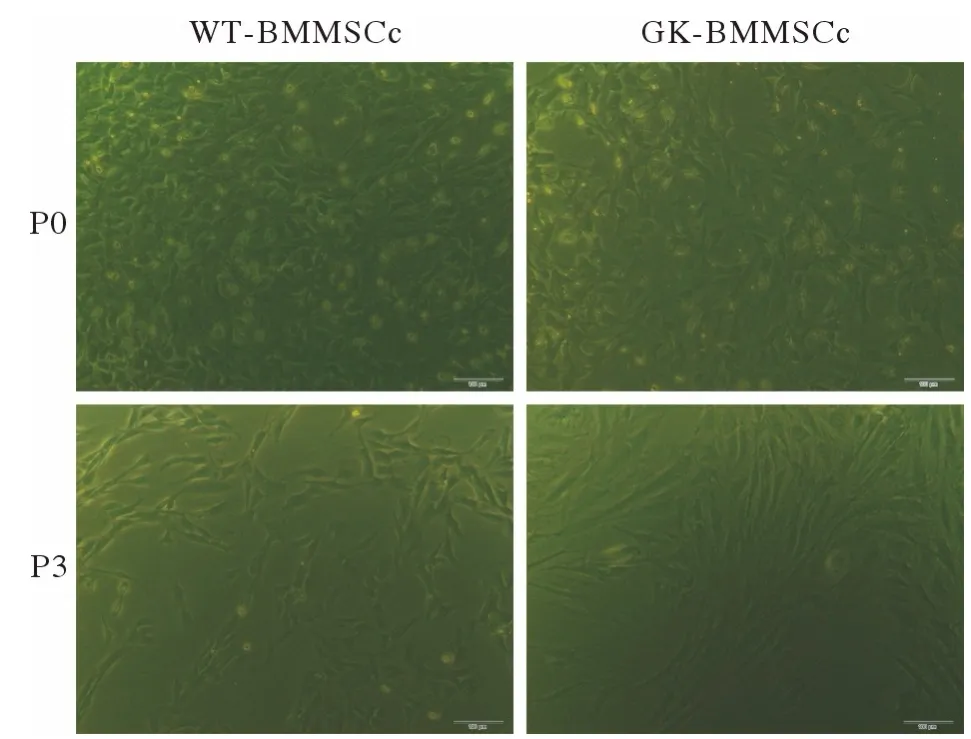
图3 倒置显微镜观察原代(P0)与第3 代(P3)正常和糖尿病大鼠BMMSCs (标尺=100 µm)Fig.3 Primary and the third generation BMMSCs derived fromnormal and type 2 diabetic rats (scale bar=100 µm)
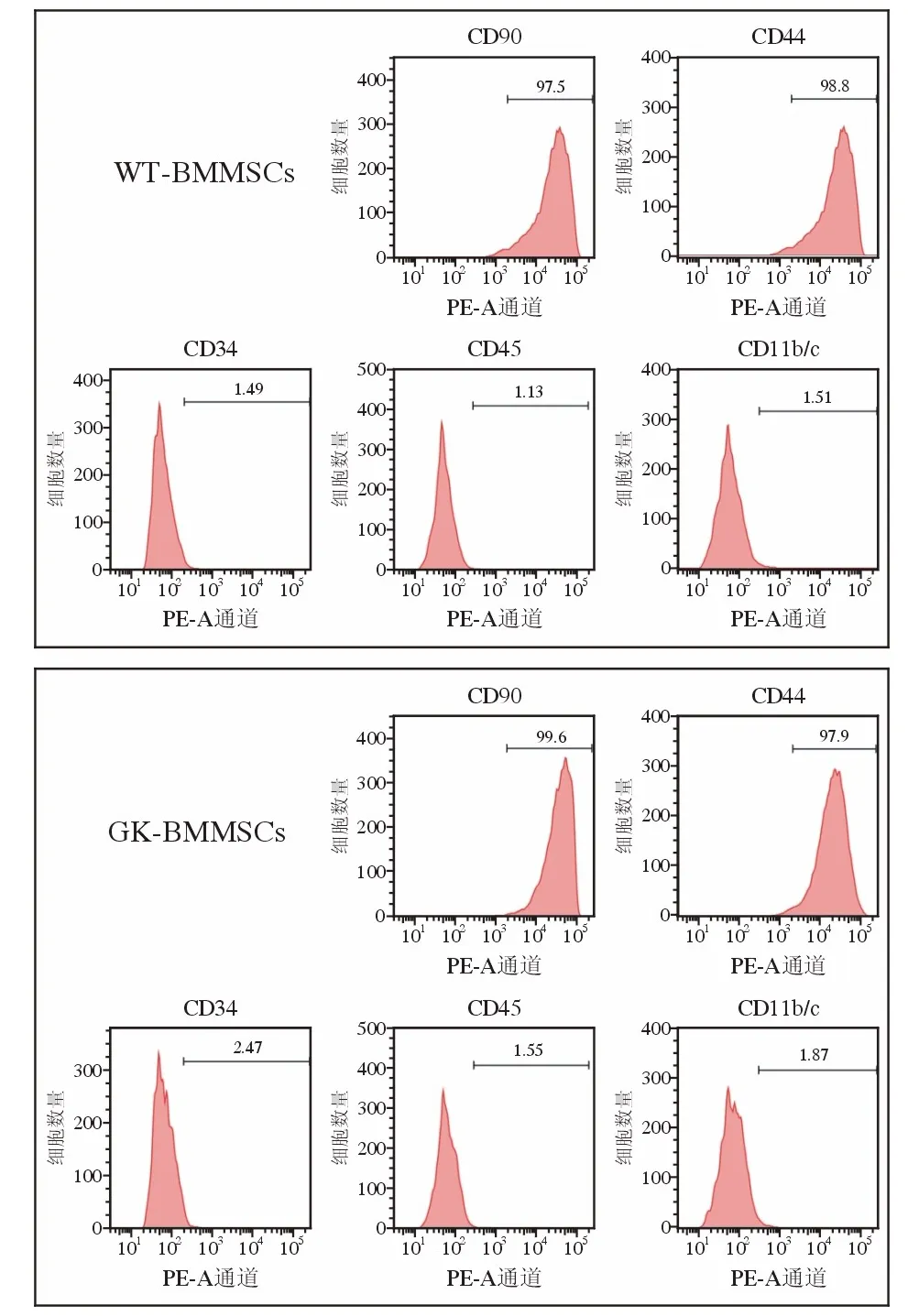
图4 流式细胞学鉴定BMMSCs 表面抗原F ig.4 Identification of BMMSCs surface antigens by flow cytometry
3 WT-BMMSCs 和GK-BMMSCs 的生物学特性比较
3.1 细胞生长曲线 CCK-8 结果显示,两组细胞的生长曲线均呈现出生长潜伏期、对数生长期和平台期,从4 d 开始,WT-BMMSCs 的增殖速率显著高于GK-BMMSCs (P<0.01;图5)。
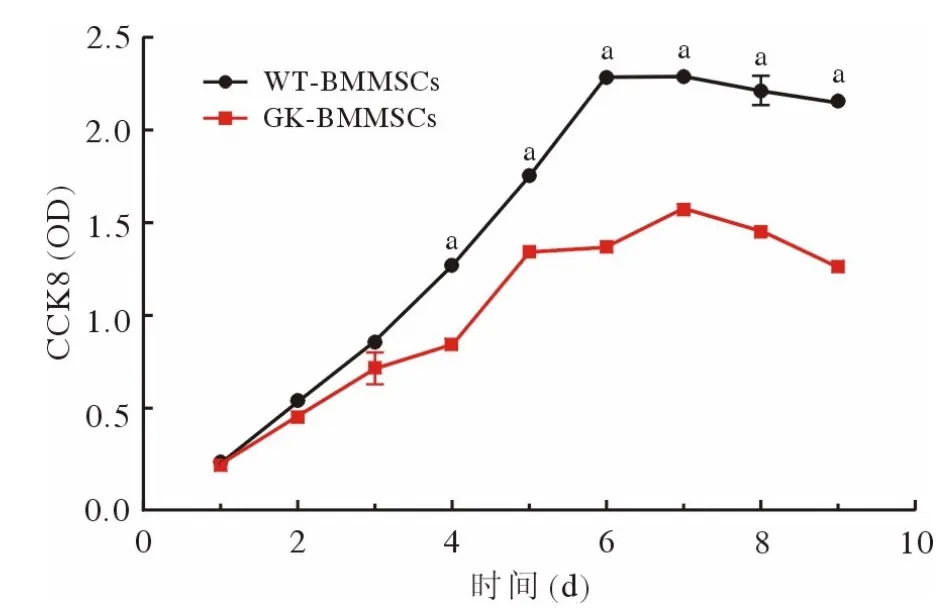
图5 CCK-8 试剂盒检测正常和糖尿病大鼠股骨BMMSCs 生长曲线(n=6;aP<0.01,vsGK)Fig.5 Detection of growth curve of BMMSCs derived from normaland type 2 diabetic rats by CCK-8 kit (n=6; aP<0.01, vsGK)
3.2 集落形成能力 结晶紫染色结果显示,WTBMMSCs 和GK-BMMSCs 培养10 d 时均有细胞集落形成,WT-BMMSCs 集落形成率显著高于GKBMMSCs (P<0.01;图6)。
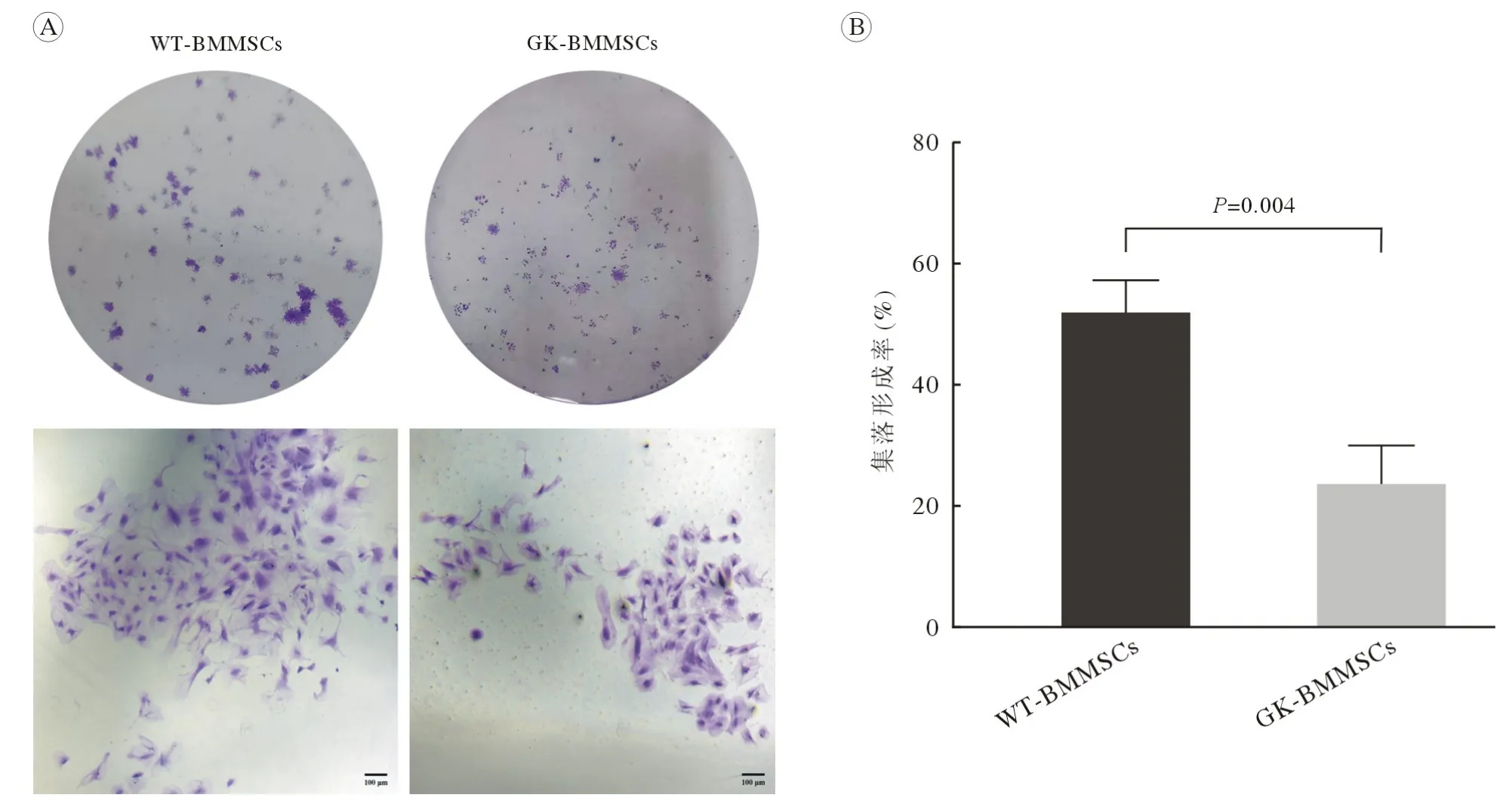
图6 正常和糖尿病大鼠股骨BMMSCs 集落形成实验A:结晶紫染色检测正常和糖尿病大鼠10 d 时股骨BMMSCs 集落形成能力(标尺=100 µm);B:正常和糖尿病大鼠股骨BMMSCs 集落形成率(n=6)Fig.6 Colony formation experiment of femoral BMMSCs in normal rats and type 2 diabetic rats A: Crystal violet staining were performed on day 10 (scale bar=100 µm); B: Colony forming efficiency of the two group (n=6)
3.3 成骨分化能力 成骨诱导7 d 后ALP 染色结果表明,WT-BMMSCs 染色较GK-BMMSCs 深,ALP 半定量检测结果与染色结果一致(P<0.001;图7A、图7B)。成骨诱导21 d 后茜素红染色结果显示,正常组和GK-BMMSCs 均出现大小不等的钙化结节,WT-BMMSCs 染色较GK-BMMSCs深(图7C)。
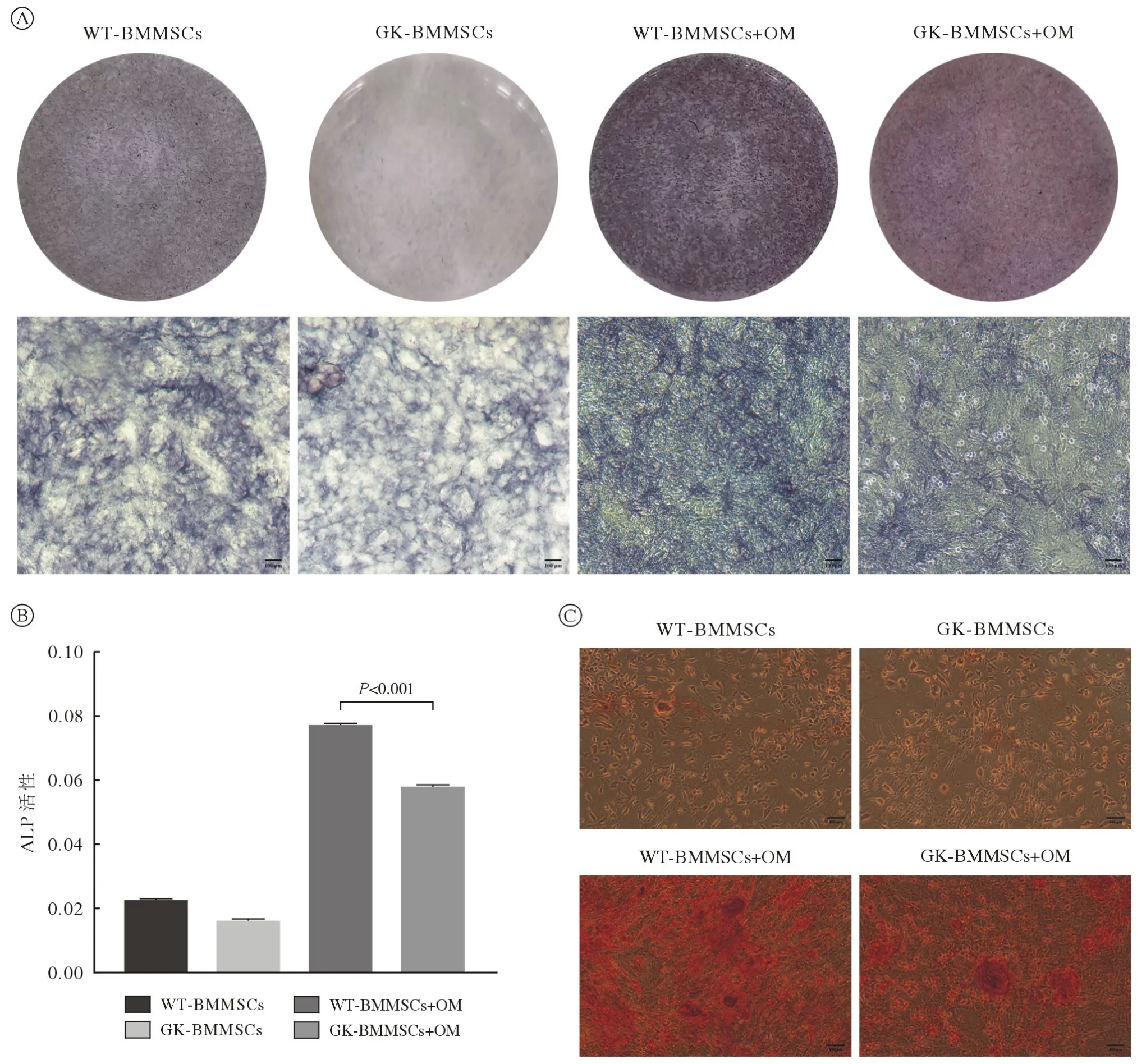
图7 正常与2 型糖尿病大鼠BMMSCs 成骨分化能力比较A:成骨诱导7 d 碱性磷酸酶染色(标尺=100 µm);B:成骨诱导7 d ALP 半定量分析 (n=6);C:成骨诱导21 d 茜素红染色显示钙化结节形成(标尺=100 µm)Fig.7 Comparison of osteogenic differentiation capacity of BMMSCs derived from normal rats and type 2 diabetic rats A: ALP staining was performed on day 7 after osteogenic differentiation (scale bar=100µm); B: Quantification of ALP activity was shown (n=6); C: ARS staining was performed on day 21 after osteogenic differentiation to show the formation of calcified nodules(scale bar=100µm)
4 WT-BMMSCs 和GK-BMMSCs 成骨分化相关基因的表达 qRT-PCR 结果显示,成骨诱导7 d时WT-BMMSCs 和GK-BMMSCs 成骨诱导组成骨分化相关基因ALP、OCN、RUNX2、COL1 的表达均显著高于未诱导组(P<0.05),而GK-BMMSCs成骨诱导组的表达显著低于WT-BMMSCs 成骨诱导组(P<0.01;图8)。Western blot 结果显示,WTBMMSCs 诱导组成骨基因ALP、OCN 表达均显著高于未诱导组,GK-BMMSCs 成骨诱导组的表达显著低于WT-BMMSCs 成骨诱导组(P<0.001),而GK-BMMSCs 成骨诱导组的表达与未诱导组无统计学差异。WT-BMMSCs 和GK-BMMSCs 成骨诱导组成骨基因RUNX2 表达均显著高于未诱导组,而GK-BMMSCs 成骨诱导组的表达显著高于WT-BMMSCs 成骨诱导组(P<0.001;图9)。
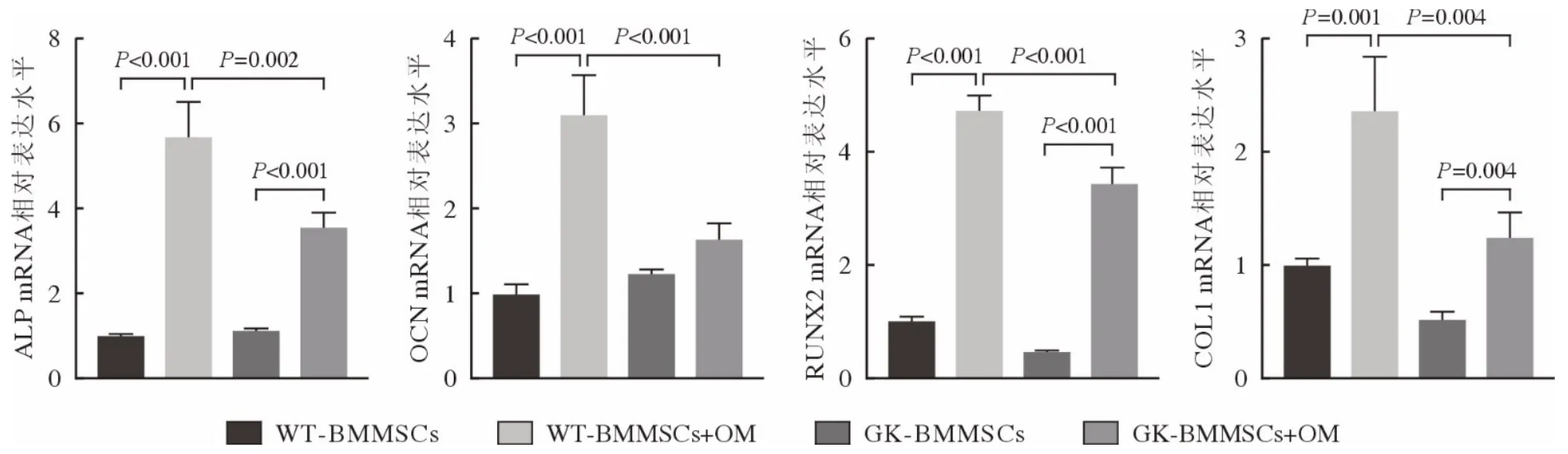
图8 成骨诱导7 d 正常与2 型糖尿病大鼠BMMSCs ALP、OCN 、RUNX2、COL1 的mRNA 表达(n=3)Fig.8 Expression level of the osteoblast differentiation-related genes ALP, OCN, RUNX2, COL1 on day 7 after osteogenic differentiation by real-time PCR (n=3)
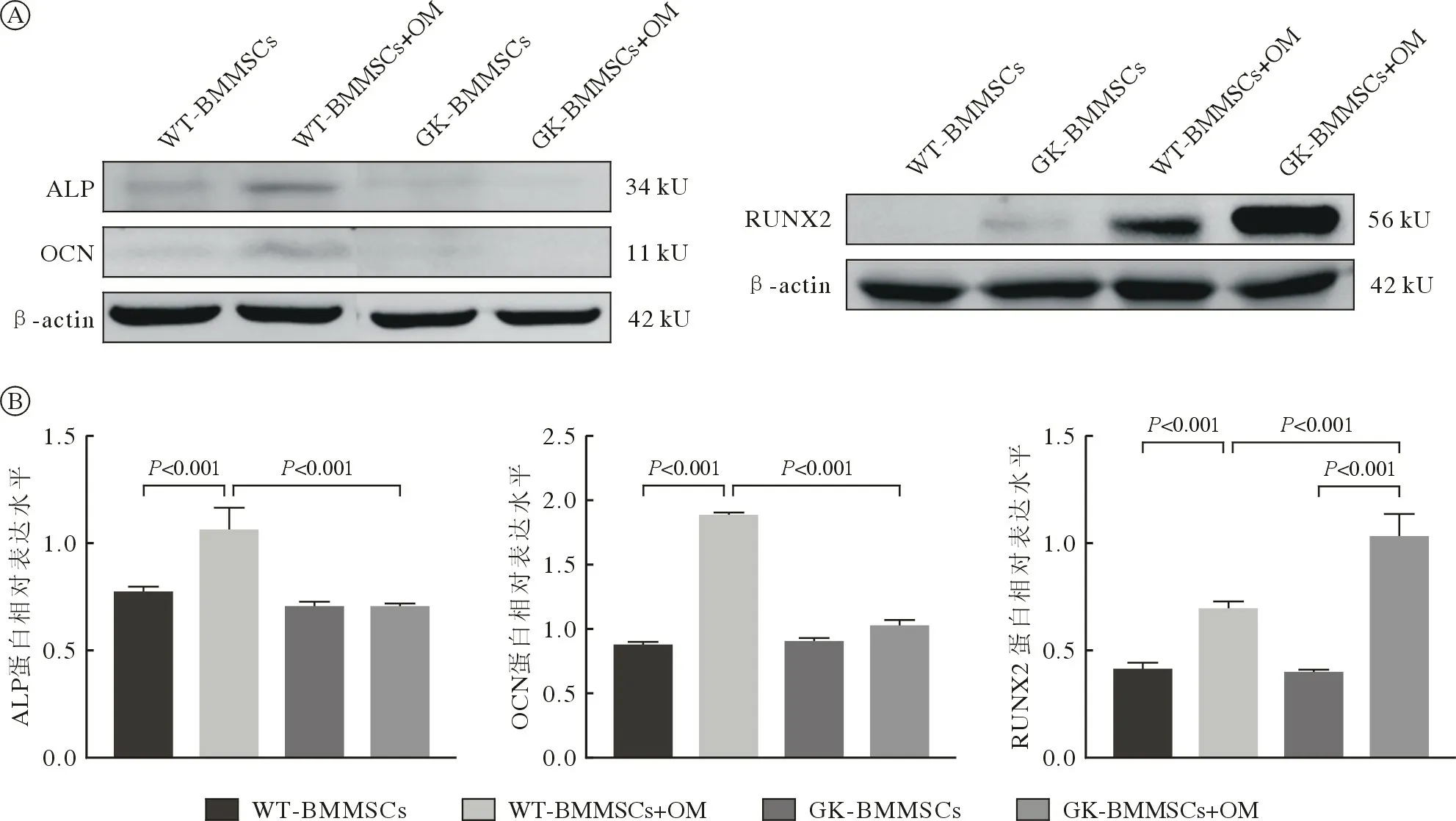
图9 成骨诱导7 d 正常与2 型糖尿病大鼠BMMSCs ALP、OCN、RUNX2 蛋白水平的表达A: ALP、OCN、RUNX2 蛋白的表达; B:ALP、OCN 、RUNX2 蛋白水平表达的定量分析(n=3)Fig.9 Western blot analysis of ALP, OCN, RUNX2 on day 7 after osteogenic differentiation A:Western blotting of ALP, OCN, RUNX2;B:Protein levels of ALP, OCN, RUNX2 quantified by densitometry (n=3)
5 WT-BMMSCs 和GK-BMMSCs成骨分化中miR-145-5p、SEMA3A 及β-catenin 的表达 qRT-PCR 结果显示,成骨诱导7 d 时WT-BMMSCs miR-145-5p表达显著低于未诱导组(P<0.05),GK-BMMSCs成骨诱导组的表达相反且显著高于WT-BMMSCs成骨诱 导 组(P<0.001)。WT-BMMSCs 和GKBMMSCs 成骨诱导组SEMA3A 的表达均显著低于未诱导组(P<0.001),GK-BMMSCs 成骨诱导组的表达显著低于WT-BMMSCs 成骨诱导组(P<0.05)。WT-BMMSCs 和GK-BMMSCs 成骨诱导组成骨通路关键蛋白β-catenin 的表达均显著高于未诱导组(P<0.01),GK-BMMSCs 成骨诱导组的表达显著低于WT-BMMSCs 成骨诱导组(P<0.01;图10)。Western blot 结果显示,WT-BMMSCs 和GK-BMMSCs 诱导组SEMA3A 表达均显著高于未诱导组,GK-BMMSCs 诱导组的表达与WTBMMSCs 诱导组无统计学差异(P<0.05;图11)。
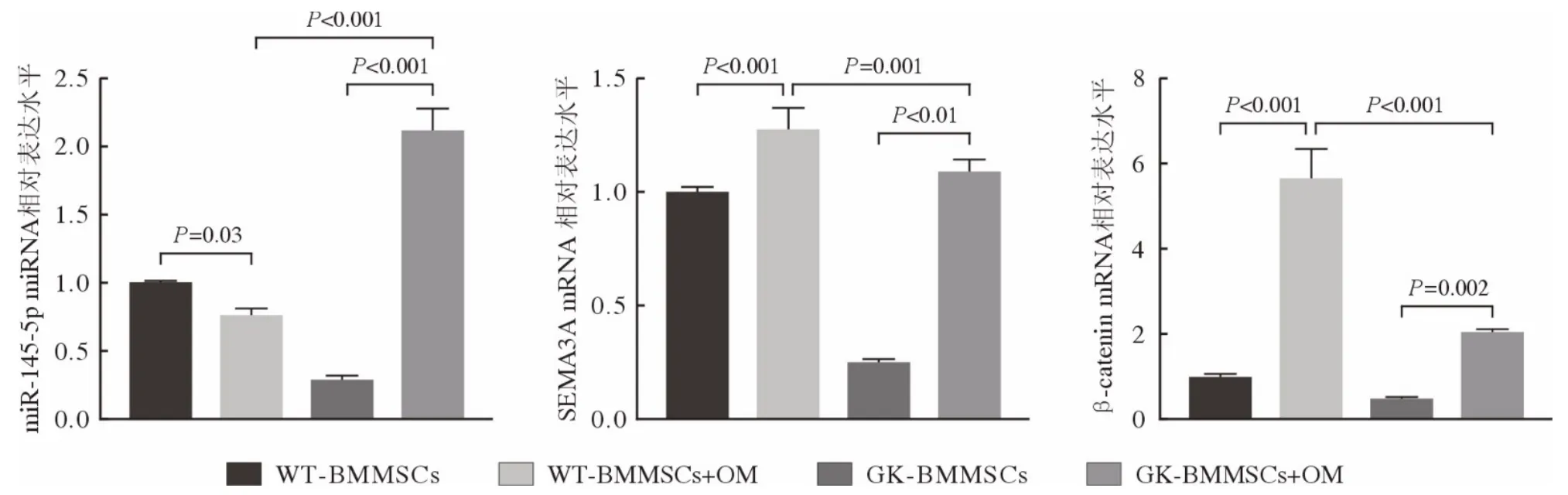
图10 成骨诱导7 d 正常与2 型糖尿病大鼠BMMSCs miR-145-5p、SEMA3A、β-catenin 的表达(n=3)Fig.10 Expression levels of miR-145-5p, SEMA3A, β-catenin on day 7 after osteogenic differentiation by real-time PCR (n=3)
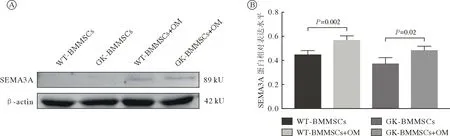
图11 成骨诱导7 d 正常与2 型糖尿病大鼠BMMSCs SEMA3A 的蛋白表达A:SEMA3A 蛋白的表达;B:SEMA3A 蛋白水平表达的定量分析(n=3)Fig.11 Western blot analysis of SEMA3A on day 7 after osteogenic differentiation A: Western blotting of SEMA3A; B: Protein levels of SEMA3A quantified by densitometry (n=3)
讨 论
糖尿病作为一种常见代谢性疾病,其引起的骨质疏松等疾病严重危害人类健康[14]。BMMSCs是理想的骨再生医学种子细胞,具有自我更新和多向分化潜能,其成骨分化功能降低是骨质疏松症发病的重要机制[13]。以往研究表明,糖尿病高糖微环境会导致BMMSCs 成骨分化能力降低从,而导致骨密度降低和骨再生障碍,其具体机制尚不十分清楚[5-6]。研究显示,miRNAs 在干细胞成骨分化中具有重要的作用。因此,本研究探讨了正常大鼠与2 型糖尿病大鼠BMMSCs 成骨分化能力及在成骨分化中miR-145-5p、SEMA3A、β-catenin表达的差异。
GK 大鼠是非肥胖自发Ⅱ型糖尿病大鼠模型,表现出与人类糖尿病相似的代谢、内分泌、血管疾病和糖尿病并发症。高血糖和胰岛素抵抗是T2DM 的主要症状。本研究中高脂高糖饲料喂养后GK 大鼠的高血糖符合T2DM 的特点。实验成功建立了稳定的2 型糖尿病大鼠模型[15]。我们培养的正常大鼠和2 型糖尿病大鼠BMMSCs 阳性表达间充质干细胞表面标志物,阴性表达造血干细胞表面标志物并能向成骨分化,符合干细胞特性[16]。2 型糖尿病大鼠BMMSCs 的增殖能力、细胞集落形成能力和成骨分化能力降低,这与以往研究结果一致[5,17],提示2 型糖尿病导致股骨BMMSCs 的功能障碍。有研究证实,干细胞在成骨分化过程中经历增殖期、分化期、矿化期和凋亡期四个阶段,而后形成骨细胞,在增殖期(1 ~7 d),增殖相关基因、COL1 和成骨分化相关基因RUNX2、OPN 高表达,ALP、OCN 低表达;在分化期(7 ~ 14 d),增殖相关基因、COL1 和成骨分化相关基因RUNX2、OPN 表达下降,ALP、OCN表达升高[18]。值得注意的是,在本研究中,培养7 d 时,2 型糖尿病大鼠BMMSCs 的增殖能力、细胞集落形成能力和成骨分化相关标志物ALP、OCN 的蛋白表达水平低于正常大鼠BMMSCs,而RUNX2 蛋白表达水平却高于正常大鼠BMMSCs。这些结果提示2 型糖尿病大鼠BMMSCs 从增殖期向分化期转变晚于正常大鼠BMMSCs。
研究表明,miR-145-5p 抑制细胞成骨成软骨分化。Zhu 等[8]报道miR-145-5p 靶向SOX9 抑制正常人脂肪间充质干细胞COL2、聚集蛋白聚糖的表达。Yu 等[19]报道miR-145-5p 抑制TLR4 信号通路降低正常人脂肪间充质干细胞成骨分化能力。Liu 等[10]报道miR-145-5p 靶向SEMA3A 降低Wnt通路基因 Wnt3a、Wnt10a 的表达,抑制2 型糖尿病大鼠脂肪间充质干细胞成骨分化相关基因ALP、RUNX2、COL1 的表达。我们的研究发现,在正常大鼠BMMSCs 成骨分化中,成骨分化相关标志物ALP、OCN、COL1、RUNX2 表达升高,miR-145-5p 表达降低,SEMA3A 和β-catenin 表达升高,这些结果与以上研究一致。提示miR-145-5p 可能通过靶向SEMA3A 调控β-catenin 来抑制正常大鼠BMMSCs 成骨分化。而在2 型糖尿病大鼠BMMSCs 成骨分化中,miR-145-5p、SEMA3A和β-catenin 表达均升高;miR-145-5p 和SEMA3A不是负向调控,可见miR-145-5p 在糖尿病源MSCs 成骨分化中有多种调控途径,其具体的调控网络仍存在争议,我们后续研究将进一步探讨。另有研究者报道在骨关节炎患者的软骨组织中,miR-145-5p 的表达显著高于非骨关节炎患者,miR-145-5p 靶向Smad4 降低 Sox-9、Col-2a1、聚集蛋白聚糖的表达,抑制骨关节炎患者源骨髓间充质干细胞的增殖和成软骨分化[9]。在我们的研究中,经成骨诱导后,2 型糖尿病大鼠BMMSCs miR-145-5p 的表达显著高于正常BMMSCs,SEMA3A和β-catenin 表达显著低于正常BMMSCs。这些结果提示miR-45-5p 在2 型糖尿病大鼠BMMSCs 成骨分化关键途径中也发挥重要的抑制作用。
综上所述,本研究结果提示,2 型糖尿病大鼠BMMSCs 细胞增殖、集落形成及成骨分化能力降低,我们推测正常大鼠BMMSCs 成骨分化中miR-145-5p 起抑制作用,2 型糖尿病大鼠BMMSCs 成骨分化中miR-145-5p 起促进作用,miR-145-5p 是否靶向SEMA3A 通过Wnt/β-catenin 调控成骨仍需要进一步研究。miR-145-5p 或许可作为治疗糖尿病引起的骨代谢疾病的关键靶点。
作者贡献戴雅文:细胞及分子生物学实验,文章撰写;鄂玲玲:课题设计,课题指导;郑颖、马小草:文章修改,文章审阅,文章校正;张戎:文献收集和整理;时权:统计分析;刘洪臣:写作指导,修订,终审论文。
利益冲突所有作者声明无利益冲突。
数据共享声明本篇论文相关数据可依据合理理由从作者处获取,Email:395298305@qq.com。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:21:20
口腔医学(2021年10期)2021-12-02 02:08:00
中学生物学(2021年8期)2021-11-02 04:53:14
生物学通报(2020年10期)2020-08-13 08:52:26
中国果业信息(2019年1期)2019-01-05 17:41:42
中国畜牧兽医文摘(2018年6期)2018-07-28 02:30:16
知识经济·中国直销(2017年10期)2017-11-07 02:39:52
中华老年口腔医学杂志(2016年2期)2017-01-15 14:24:47
中国病理生理杂志(2015年8期)2015-12-21 12:38:14
天津护理(2015年4期)2015-11-10 06:11:41
