
盐胁迫与供磷水平对柳枝稷苗期光合特性与总生物量的影响
2023-08-01 11:27:50麻仲花陈娟吴娜满本菊王晓港者永清刘吉利
中国农业科技导报 2023年6期
麻仲花, 陈娟, 吴娜, 满本菊, 王晓港, 者永清, 刘吉利
(1.宁夏大学农学院,银川 750021; 2.宁夏大学生态环境学院,银川 750021)
柳枝稷(Panicum virgatumL.)作为禾本科黍属多年生草本C4植物,适宜在盐碱地种植[1]。土地盐渍化是目前农业生产中普遍存在的问题,严重制约植物的生长发育,影响农作物的产量和品质。磷作为植物必需营养元素之一,对植物光合作用和生长发育起至关重要的作用。但盐胁迫抑制植物生长发育的同时,也显著降低了土壤中有效磷的利用率,甚至限制根系对磷的吸收利用,影响作物产量。在农业生产中通常用磷肥来补充土壤中有效磷的含量[2-3]。黄高鉴等[4]研究表明,盐胁迫显著抑制柳枝稷的生长发育,使其生物量显著降低。常雯雯等[5]研究表明,盐胁迫条件下柳枝稷的光合特性显著降低,但不同品种间存在差异。刘金彪等[6]认为,低磷胁迫环境下柳枝稷为吸收磷素而蒸腾损失水分,减弱光合作用,从而降低生物量。张蕊等[7]认为,在逆境胁迫下增施磷肥可增强植物光合生理特性及植物生长发育能力,从而缓解逆境对植物造成的伤害,进而提高产量。目前,在盐胁迫条件下,磷素的流失对柳枝稷光合特性及生物量影响的研究多集中在盐胁迫或供磷水平单因素上。柳枝稷苗期对盐和低磷胁迫均比较敏感[6,8],因此,本研究采用水培试验研究盐胁迫与供磷水平2种因素对不同品种柳枝稷苗期光合特性及生物量的影响,并进一步揭示苗期柳枝稷光合特性与总生物量的关系,为柳枝稷在盐碱地的推广种植提供理论依据。
1 材料与方法
1.1 试验材料
试验在北京市农林科学院温室大棚内开展,以3个不同品种柳枝稷(八倍体Alamo、Alamo和Pathfinder)为试验材料,品种特性详见表1。
1.2 试验设计
采用水培试验,设置盐胁迫和供磷水平2个因素,其中盐胁迫分别设置0(A1)和100 mmol·L-1NaCl(A2)共2个水平;磷肥施用量分别设置0(B1)、5(B2)、200(B3)和2 000 mmol·L-1KH2PO4(B4)4个水平,共计8个处理,分别为A1B1、A1B2、A1B3、A1B4、A2B1、A2B2、A2B3、A2B4,每个处理3次重复。
挑选大小一致饱满的柳枝稷种子,通过沙培在光照培养箱中育苗(培养条件为:昼/夜室温35 ℃/20 ℃,光周期14 h/10 h,光照强度400 μM·m-2·s-1),待幼苗长到3片展开叶,挑选生长一致的幼苗移栽至水培箱(箱体长41 cm,宽30.5 cm,高13.5 cm)中培养。每个水培箱内,幼苗基部用海绵包裹,放入聚乙烯泡沫板预先打好的孔中,然后固定于水培箱中。水培箱体用锡箔纸包裹,以防止光照,并避免藻类的繁殖。每个水培箱灌注营养液13.9 L,用气泵进行24 h充气,营养液每3 d更换1次。试验营养液配比采用不同磷水平的Hoagland营养液(配方详见表2)。KH2PO4为营养液中的磷素来源,为保持钾素的平衡,改变KH2PO4的含量时需调节相应KCl含量。柳枝稷材料先在正常条件下培养至3叶1心,然后进行不同盐胁迫和供磷水平处理,处理第7天进行各项指标测定。每个处理种植6盆,每盆种植16株。
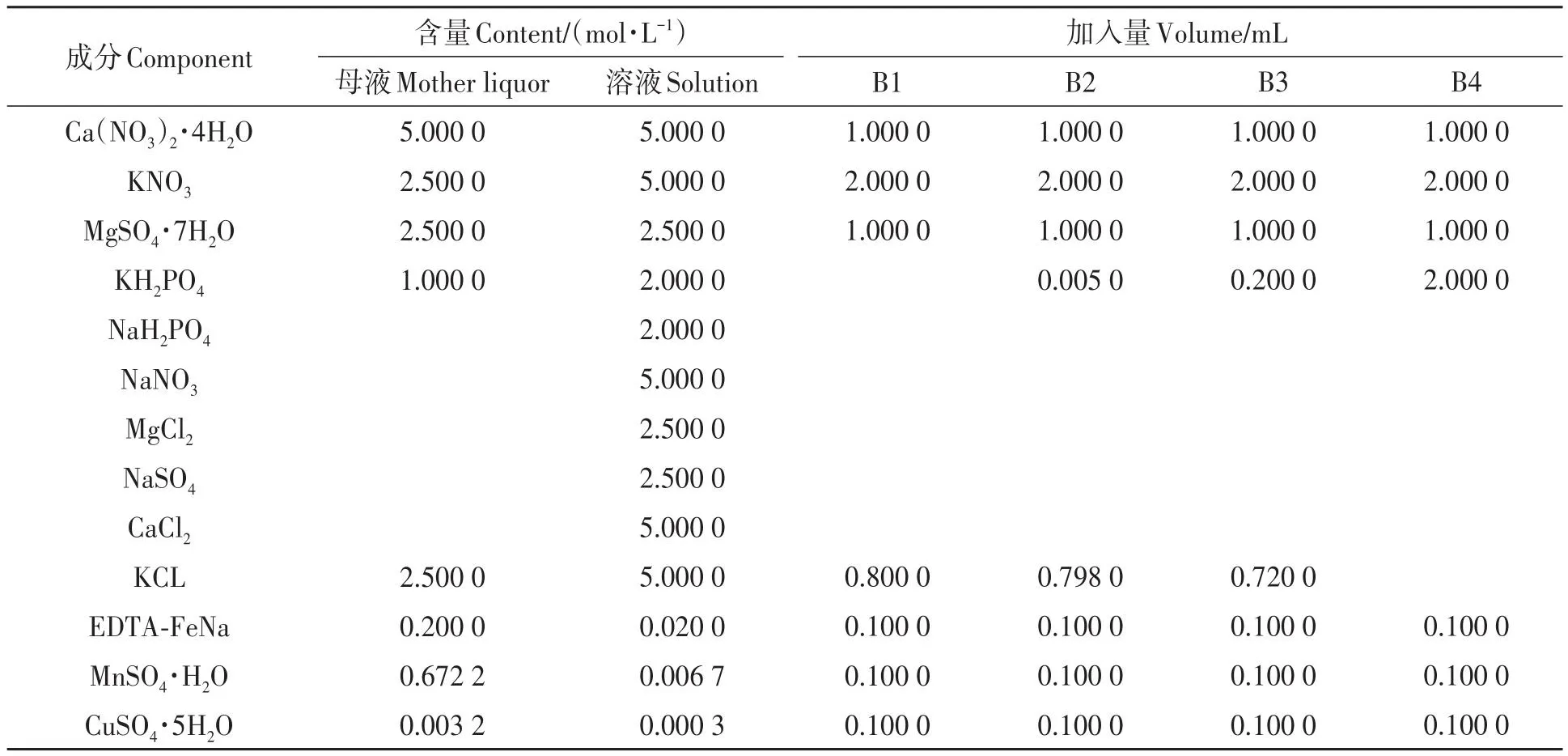
表2 不同磷水平的Hoagland营养液配置Table 2 Hoagland nutrient solution configuration table with different phosphorus levels
1.3 测定指标及方法
(1)叶绿素含量采用乙醇浸提法进行测定:称取叶片 0.05~0.08 g,记下具体质量,剪碎后置于离心管中;加入8 mL 95%的乙醇,避光静置24 h;分别于665、649 nm 下测定其吸光度值,并计算叶绿素含量。
(2)净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、蒸腾速率(transpiration rate,T)r、胞间CO2浓度(intercellular CO2concentration,C)i采用LI-6400光合仪进行测定。具体方法为:于晴朗天气9∶00—10∶00,选取其各株从上往下数第1片叶进行测定,每个处理随机测量5株。
(3)生物量测定:将整株取出,测定单株地上部分生物量鲜重;将待测植株冲洗干净,置于105 ℃鼓风干燥箱中杀青30 min,然后在65 ℃下烘干至恒重,测定单株地上部分生物量干重。
1.4 数据处理
采用Excel 2016、SPSS 25.0和0rgin 2020统计软件进行方差分析、相关性分析、模糊数学隶属函数分析。隶属函数的计算方法[9]如下。
式中,Xi为i指标测定值,Ximin和Ximax分别为i指标的最小值和最大值。
2 结果分析
2.1 盐胁迫与供磷水平对苗期柳枝稷光合特性及总生物量的影响
2.1.1 盐胁迫与供磷水平对叶绿素含量的影响由图1可知,3个柳枝稷品种的叶片叶绿素含量在同一盐处理下均随着供磷水平的增加呈上升趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder的叶绿素含量随着磷水平增加均呈现上升的趋势,其中全磷处理的叶绿素含量较无磷处理分别提高17.45%、95.16%、86.87%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder叶绿素含量随着磷水平增加均呈现上升的趋势,其中全磷处理的叶绿素含量较无磷处理分别提高115.45%、188.33%、108.20%。
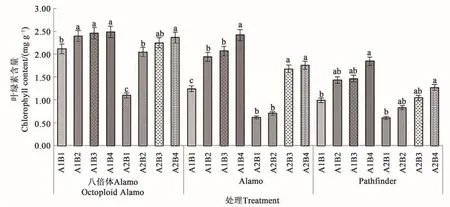
图1 不同盐胁迫与供磷水平下3个柳枝稷品种的叶绿素含量Fig.1 Chlorophyll content of 3 switchgrass varieties under different salt stress and phosphorus supply
由表3可知,盐胁迫显著降低了Alamo和Pathfinder叶绿素含量,但对八倍体Alamo影响不显著;供磷显著增加了Pathfinder叶绿素含量,但对八倍体Alamo和Alamo影响不显著;两因素间的交互作用对八倍体Alamo和Alamo的叶绿素含量影响显著,但对Pathfinder影响不显著。
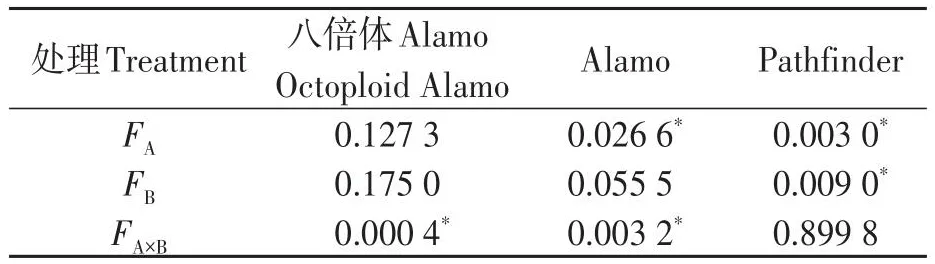
表3 叶绿素含量方差分析Table 3 Analysis of variance of chlorophyll content
2.1.2 盐胁迫与供磷水平对净光合速率的影响由图2可知,3个柳枝稷品种的叶片净光合速率在同一盐处理下均随着供磷水平的增加呈上升趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder净光合速率随着磷水平增加均呈现上升的趋势,其中全磷处理的净光合速率较无磷处理分别提高21.84%、15.89%、18.33%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder净光合速率随着磷水平增加均呈现上升的趋势,其中全磷处理的净光合速率较无磷处理分别提高14.63%、15.28%、23.35%。

图2 不同盐胁迫与供磷水平下3个柳枝稷品种的净光合速率Fig.2 Pn of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表4可知,盐胁迫显著降低了八倍体Alamo、Alamo和Pathfinder的净光合速率;供磷显著增加了Alamo和Pathfinder的净光合速率,但对八倍体Alamo净光合速率影响不显著;两因素的交互作用对八倍体Alamo、Alamo和Pathfinder的净光合速率影响均不显著。

表4 净光合速率方差分析Table 4 Analysis of variance of Pn
2.1.3 盐胁迫与供磷水平对气孔导度的影响 由图3可知,3个柳枝稷品种苗期叶片气孔导度在同一盐处理下均随着供磷水平的增加呈上升趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder气孔导度随着磷水平增加均呈现上升的趋势,其中全磷处理的气孔导度较无磷处理分别提高42.86%、28.57%、75.00%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder气孔导度随着磷水平增加均呈现上升的趋势,其中全磷处理的气孔导度较无磷处理分别提高16.67%、100.00%、100.00%。
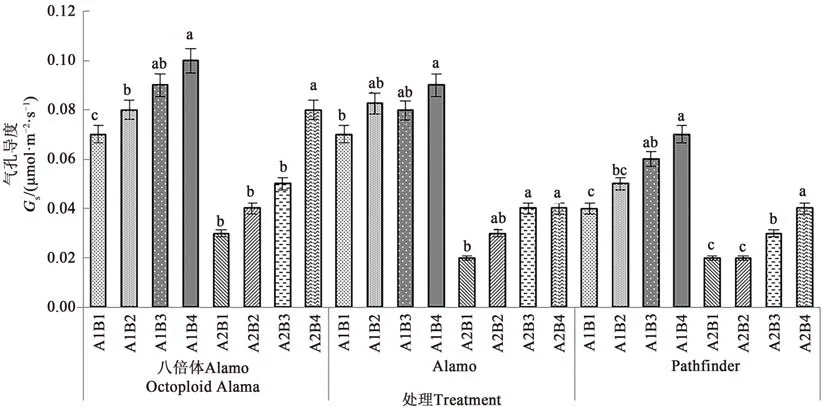
图3 不同盐胁迫与供磷水平下3个柳枝稷品种的气孔导度Fig.3 Gs of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表5可知,盐胁迫显著降低了八倍体Alamo、Alamo和Pathfinder苗期叶片气孔导度;供磷显著增加了Pathfinder气孔导度,但对八倍体Alamo和Alamo的影响不显著;两因素的交互作用对八倍体Alamo和Pathfinder影响均不显著,但对Alamo影响显著。
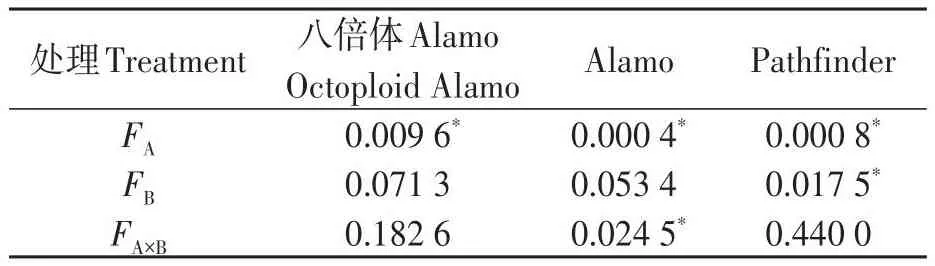
表5 气孔导度方差分析Table 5 Analysis of variance of Gs
2.1.4 盐胁迫与供磷水平对胞间CO2浓度的影响 由图4可知,3个柳枝稷品种苗期叶片胞间CO2浓度在同一盐处理下均随着供磷水平的增加呈下降趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder胞间CO2浓度随着磷水平增加均呈现下降趋势,其中全磷处理的胞间CO2浓度较无磷处理分别降低50.12%、68.07%、63.11%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder胞间CO2浓度随着磷水平增加均呈现下降趋势,其中全磷处理的胞间CO2浓度较无磷处理分别降低49.68%、40.41%、40.21%。

图4 不同盐胁迫与供磷水平下3个柳枝稷品种的胞间CO2浓度Fig.4 Ci of 3 switchgrass varieties under different salt stress and phosphorus supply level
由表6可知,盐胁迫增加了八倍体Alamo、Alamo和 Pathfinder胞间CO2浓度;供磷显著降低了Pathfinder胞间CO2浓度,但对八倍体Alamo和Alamo影响不显著;两因素的交互作用对八倍体Alamo、Alamo和 Pathfinder影响显著。
2.1.5 盐胁迫与供磷水平对蒸腾速率的影响 由图5可知,3个柳枝稷品种蒸腾速率在同一盐处理下均随着供磷水平的增加呈上升趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder蒸腾速率随着磷水平增加均呈现上升的趋势,其中全磷处理的蒸腾速率较无磷处理分别提高21.98%、37.62%、42.60%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder蒸腾速率随着磷水平增加均呈现上升的趋势,其中全磷处理的蒸腾速率较无磷处理分别提高71.10%、37.20%、50.35%。

图5 不同盐胁迫与供磷水平下3个柳枝稷品种的蒸腾速率Fig.5 Tr of 3 switchgrass varieties under different salt stress and phosphorus supply level
由表7可知,盐胁迫显著降低了八倍体Alamo、Alamo和Pathfinder蒸腾速率;供磷对八倍体Alamo和Pathfinder蒸腾速率影响显著,但对Alamo影响不显著;两因素的交互作用对Alamo和Pathfinder蒸腾速率影响显著,但对八倍体Alamo影响不显著。

表7 蒸腾速率方差分析Table 7 Analysis of variance of Tr
2.1.6 盐胁迫与供磷水平对总生物量的影响 由图6可知,3个柳枝稷品种总生物量在同一盐处理下均随着供磷水平的增加呈上升趋势。在非盐胁迫下,八倍体Alamo、Alamo和Pathfinder总生物量随着磷水平增加均呈现上升的趋势,其中全磷处理的总生物量较无磷处理分别提高246.52%、115.32%、145.85%。在盐胁迫条件下,八倍体Alamo、Alamo和Pathfinder总生物量随着磷水平增加均呈现上升的趋势,其中全磷处理的总生物量较无磷处理分别提高91.00%、113.97%、405.88%。
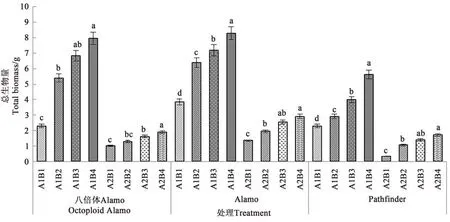
图6 盐胁迫与供磷水平下3个柳枝稷品种的总生物量Fig.6 Biomass of 3 switchgrass varieties under different salt stress and phosphorus supply levels
由表8可知,盐胁迫显著降低了八倍体Alamo、Alamo和Pathfinder总生物量;供磷对八倍体Alamo、Alamo和Pathfinder总生物量均无显著影响;两因素的交互作用对八倍体Alamo、Alamo和Pathfinder总生物量有显现影响。

表8 总生物量方差分析Table 8 Analysis of variance of total biomass
2.2 不同柳枝稷品种苗期总生物量与光合特性的相关性分析
分析不同柳枝稷品种苗期总生物与光合特性之间的关系,结果(表9)表明,八倍体Alamo、Alamo和Pathfinder的总生物量与叶绿素含量、蒸腾速率、气孔导度、净光合速率均呈正相关关系,与胞间CO2浓度呈负相关关系。其中,八倍体Alamo总生物量与蒸腾速率、气孔导度、净光合速率呈显著正相关,相关系数分别为0.88、0.85、0.89,与胞间CO2浓度呈显著负相关,相关系数为0.83;Alamo总生物量与叶绿素含量呈显著正相关,相关系数为0.86,与胞间CO2浓度呈显著负相关,相关系数为0.92。

表9 3个柳枝稷品种苗期总生物量与光合特性的相关性分析Table 9 Correlation analysis of total biomass and photosynthetic characteristics of 3 switchgrass varieties at different seedling stage
2.3 不同柳枝稷品种苗期光合特性及总生物量的隶属函数分析
不同盐胁迫与供磷水平下各光合指标及总生物量隶属函数值的平均值如表10所示。3个柳枝稷品种各光合指标及总生物量的隶属函数值平均值表现为八倍体Alamo>Alamo>Pathfinder。其中,八倍体Alamo的光合指标及总生物量隶属函数值的综合平均值最高,为0.50,最适宜在盐胁迫与低磷胁迫条件下种植。
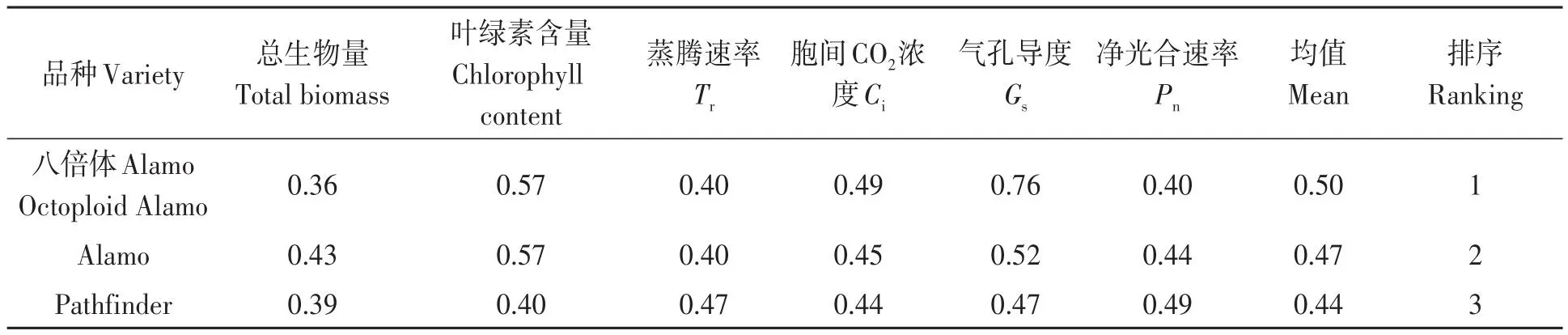
表10 3个柳枝稷品种苗期光合特性及总生物量的隶属函数值Table 10 Membership function values of photosynthetic characteristics and total biomass of 3 switchgrass varieties at seedling stage
3 讨论
植物光合作用对光能的利用是从对光合色素吸收开始的[10],叶绿素是叶片吸收、传递和转化光能的基础[11-12]。叶绿素含量可以直接反映植物光合能力的强弱,也是衡量植物抗逆能力的重要指标之一[13-14]。研究发现,盐胁迫下植物叶片的叶绿素含量显著降低[15]。本研究结果表明,盐胁迫下柳枝稷苗期叶片叶绿素含量低于非盐胁迫处理,这可能是叶绿体色素合成酶活性降低、叶绿素分解加速、叶绿体功能紊乱或结构受损等原因造成[16]。在相同盐处理下,3个柳枝稷品种苗期叶片的叶绿素含量均随着供磷水平的提高而增加。这说明适量供磷可以提高植物叶绿素含量,增强光合吸收能力[17],进而提高苗期柳枝稷的光合作用。
植物通过光合作用积累能量与贮藏营养物质,光照强弱直接影响植物生长发育[18]。气孔导度是反映光、水、气交换能力的重要生理指标,其直接影响着植物的光合速率和蒸腾速率[19-22]。研究表明,叶片的胞间CO2浓度越小,气孔内外CO2浓度差值越大,气孔能吸收的CO2越多,光合速率就越高;光合速率与气孔导度、蒸腾速率呈显著正相关,与胞间CO2浓度呈显著负相关[23]。本研究结果表明,八倍体Alamo和Alamo的净光合速率、气孔导度和蒸腾速率三者呈正相关关系,而与胞间CO2浓度呈负相关关系。研究表明,净光合速率和气孔导度升高时,胞间CO2浓度随之降低,这是由非气孔因素引起的光合作用变化[24];盐胁迫对植物幼苗的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率有抑制作用[25];补施磷肥可以提高柳枝稷光合特性[6]。本研究表明,八倍体Alamo、Alamo和 Pathfinder在盐胁迫下的净光合速率、气孔导度、蒸腾速率均低于非盐胁迫处理,而胞间CO2浓度高于非盐胁迫处理;但相同盐处理下,随着供磷水平的增加净光合速率、气孔导度、蒸腾速率随之增大,胞间CO2浓度随之减小。在同一盐处理下,全磷处理下的净光合速率、气孔导度、蒸腾速率最高,胞间CO2浓度最低,这表明相同盐处理下供磷越多柳枝稷品种苗期叶片的光合特性越强,与姜宗庆等[26]研究结果一致。
黄高鉴等[4]研究表明,盐胁迫条件下柳枝稷的生长发育明显被抑制,生物量显著降低。本研究表明,3个柳枝稷品种在盐胁迫下的生物量低于非盐胁迫,与前人研究结果一致,这是由于盐胁迫下叶绿素含量明显降低,影响了光合作用,从而使生物量显著降低[27]。陈永亮等[28]研究表明,缺磷时植物幼苗生长明显受限,生物量累积速率降低。本研究表明,3个柳枝稷品种在相同盐处理下其生物量均随着供磷水平的增加而增加,均在全磷处理下达到最大值。盐胁迫下适当施用磷肥可以补充土壤中的有效磷含量,有利于提高植物生物量[7]。本研究表明,光合作用也是决定柳枝稷品种生物量的关键因素之一,其中八倍体Alamo总生物量的增加还与蒸腾速率、气孔导度、净光合速率呈正相关,与胞间CO2浓度呈负相关;Alamo总生物量与叶绿素含量呈正相关,与胞间CO2浓度呈负相关,与常雯雯等[5]研究结果一致。3个柳枝稷品种中,八倍体Alamo的光合指标及总生物量隶属函数值的平均值最高,表明其适应性最强,最宜在盐胁迫与低磷地区推广种植。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
湖北农业科学(2021年13期)2021-08-04 03:56:18
新疆农业科技(2021年2期)2021-07-03 06:39:34
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中学生物学(2020年12期)2020-03-30 07:27:46
中国农业科技导报(2020年12期)2020-03-15 10:50:08
生物技术通报(2014年10期)2014-03-21 06:56:38
