
β-甘露聚糖酶的生物学功能及其在动物生产中的应用
2023-07-14 05:37翟彩宁
饲料研究 2023年9期
翟彩宁 陈 佩
(陕西开放大学,陕西 西安 710119)
甘露聚糖广泛存在于豆粕、芝麻粕和棕榈仁粕等植物性饲料原料中,主要由β-1,4 糖苷键连接而成[1]。β-甘露聚糖作为一种抗营养因子,在单胃动物体内甘露聚糖酶的缺乏会影响畜禽对营养成分的消化吸收[2]。β-甘露聚糖酶是水解甘露聚糖的关键酶[3],可以分解细胞壁中的甘露聚糖,使消化酶与营养物质充分接触,促进营养物质的吸收。β-甘露聚糖酶还可直接作用于肠黏膜,减少病原微生物与肠黏膜上皮细胞的结合,改善肠道环境。β-甘露聚糖酶水解甘露聚糖的产物——甘露寡糖具有益生元的作用,可以调节免疫反应,提高机体免疫力[4-6]。目前,β-甘露聚糖酶广泛应用于饲料等领域,可降解饲料中的抗营养因子,提高能量的利用率,提高畜禽的生长和生产性能以及机体免疫能力[7-8]。文章主要综述了β-甘露聚糖酶的生物学功能及其在动物生产中的研究结果,为β-甘露聚糖酶进一步在动物养殖中的应用提供参考。
1 β-甘露聚糖酶的来源和结构
甘露聚糖是自然界中半纤维素的第二大组成部分,广泛存在于豆科植物中,是由β-1,4-D-吡喃甘露糖连接而成的线状多聚体[1]。甘露聚糖可作为多糖存储在植物种子的胚乳中。此外,甘露聚糖还是植物细胞壁的主要组成成分,作为结构多糖维持植物细胞结构的完整性[9]。甘露聚糖通常结构比较复杂,不同来源的甘露聚糖结构也不相同。按照不同的连接方式和单糖组成,甘露聚糖主要包括线性甘露聚糖、半乳甘露聚糖、葡甘露聚糖和半乳葡甘露聚糖等4 种[10](见图1)。
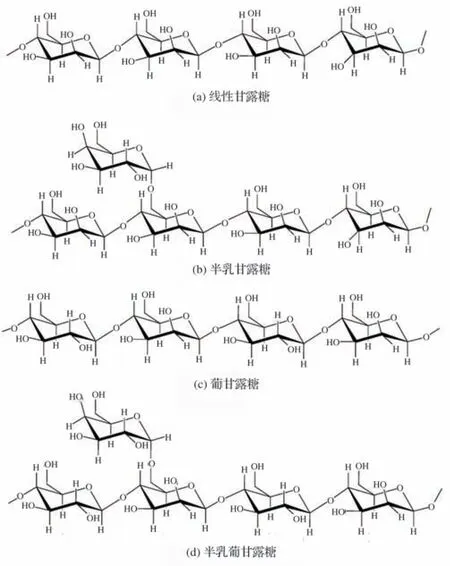
图1 甘露聚糖的分类
张建新等[11]研究表明,甘露聚糖作为一种抗营养因子影响动物对营养物质的吸收利用,完全被降解需要多种水解酶的协同作用。β-甘露聚糖酶是甘露聚糖水解过程中最关键的酶之一,通过内切的方式水解甘露聚糖的β-1,4-D-甘露糖苷键,得到不同类型的寡糖,之后在β-葡萄糖苷酶和β-甘露糖苷酶的协同作用下进一步水解寡糖的β-1,4 糖苷键,释放甘露糖和葡萄糖[12]。
β-甘露聚糖酶来源广泛,在一些植物(番茄、芝麻、豆儿瓜、长角豆)、微生物(细菌、真菌、放线菌)以及部分低等动物(蜗牛、翡翠贻贝)的肠道分泌液中均有存在[13]。饲用甘露聚糖酶主要由微生物产生,具有提取方便、生产成本低、pH 值和温度作用范围广、底物专一和酶活性高等优点[14-16],目前在理论研究和工业化生产中均得到广泛应用。
根据氨基酸序列的相似性,β-甘露聚糖酶可分为4 个糖苷水解酶家族(GH),即GH5、GH26、GH113 和GH134[17]。许多真菌、动植物来源的β-甘露聚糖酶和部分细菌来源的β-甘露聚糖酶均属于GH5 家族[18]。GH5、GH26 和GH113 家族的β-甘露聚糖酶均是GH-A 家族成员[19]。β-甘露聚糖酶由催化域和非催化域两部分组成,催化域经折叠形成典型的(β/α)8-TIM 桶状结构,由8 个α 螺旋和8 个β 折叠交替排列组成,8 个β 折叠在中部平行排列形成一个桶状结构,α 螺旋主要负责连接这些折叠片段,主要位于桶的外围[20](见图2)。β-甘露聚糖酶具有两个保守的活性位点(酸碱催化位点和亲核催化位点),分别位于第4 和第7 个β 折叠的C 末端[19]。除催化域外,部分β-甘露聚糖酶还存在非催化域,如碳水化合物结合模块(CBM) 中的经典β 三明治(β sandwich)结构,由13 个β 折叠、1 个α 螺旋和1 个钙离子构成[21],可增强酶对含纤维素甘露糖的水解能力。
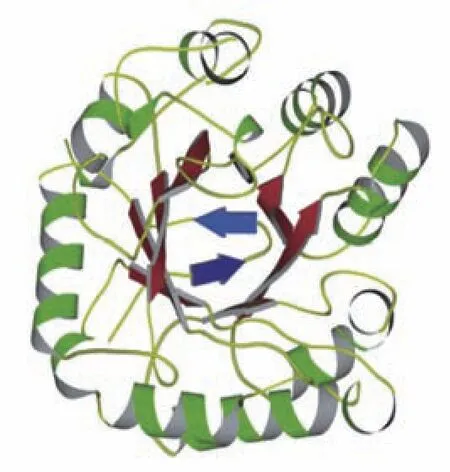
图2 β-甘露聚糖酶的分子结构
2 β-甘露聚糖酶的生物学功能
2.1 促进营养物质的消化吸收
饲料中主要的抗营养因子是非淀粉多糖,β-甘露聚糖普遍存在于植物性饲料中,占畜禽饲粮中非淀粉多糖的15%~37%。甘露聚糖酶能够消除β-甘露聚糖的抗营养作用,促进畜禽生长[22]。β-甘露聚糖因其高亲水性,易在单胃动物的消化道内吸水形成凝胶状结构,增加消化道中内容物的黏度,影响消化酶与营养物质的相互作用[23]。β-甘露聚糖酶能够水解β-甘露聚糖分子中的β-1,4 糖苷键,水解得到甘露寡糖,降低食糜黏度,促进消化酶(淀粉酶、胰蛋白酶等)的分泌,利于机体对营养物质的消化吸收[24]。此外,β-甘露聚糖酶能够降解植物性饲料细胞壁中的β-甘露聚糖,促进营养物质的释放,提高饲料的营养价值[25]。薛瑞婷等[26]研究表明,在玉米-豆粕型基础饲粮中添加β-甘露聚糖酶能够提高生长猪的养分表观消化率。
2.2 改善肠道健康
在畜禽饲粮中添加β-甘露聚糖酶后,甘露聚糖被水解为甘露寡糖。甘露寡糖能够调节肠道内菌群平衡,促进肠道中益生菌等有益菌的生长,抑制腐败菌和外源菌在消化道的定植,保障肠道黏膜的完整性[27-28]。此外,β-甘露聚糖酶直接作用于肠黏膜,通过与病原微生物表面的特异性蛋白结合,减少细菌与肠黏膜上皮细胞的结合,改善肠道健康[11]。乔海云等[29]研究发现,在肉仔鸡的基础饲粮中添加0.06%的β-甘露聚糖酶,能够显著降低大肠杆菌的数量,维持肠道酸碱度平衡,增加肠道乳酸杆菌等有益菌的数量,改善肉鸡肠道菌群平衡。李路胜等[30]研究发现,与对照组相比,在低能量饲粮中添加500 g/t 的β-甘露聚糖酶,肉鸡的胸腺指数显著提高29.70%。高会战[31]研究发现,在低豆粕饲粮中添加β-甘露聚糖酶能够显著增加十二指肠和回肠的长度,降低肉鸡十二指肠指数,改善肉鸡的肠道形态。
2.3 提高机体免疫功能
β-甘露聚糖酶能够降解甘露聚糖缓解机体肠道的免疫应激,提高动物机体免疫力。饲料中的β-甘露聚糖是一种病原相关分子结构(PAMP),在肠道中可以被模式识别受体(PRR)识别,刺激肠道中的先天免疫细胞,诱发一系列的免疫反应,导致生产效益下降[32]。在饲料中添加β-甘露聚糖酶可以消除甘露聚糖的负面作用,提高动物的生产性能[33]。此外,甘露寡糖可以促进细胞分泌糖蛋白,这些含有甘露糖基的糖蛋白能够与入侵的病原菌结合,激发机体的免疫反应,提高动物免疫力[25]。Arsenault 等[34]研究发现,饲粮中添加β-甘露聚糖酶可增加肉鸡淋巴细胞的数量,调节肠道炎症反应,提高机体的免疫力。Roque 等[32]研究发现,在荷斯坦奶牛的基础饲粮中添加0.1%的β-甘露聚糖酶能够减轻产奶牛的炎症反应。乔海云等[29]研究发现,在玉米-豆粕型饲粮中添加0.01%的β-甘露聚糖酶后,肉仔鸡的法氏囊指数和脾脏指数均显著提高,对免疫性能起到积极的促进作用。黄铁生等[35]在肉鸡饲粮中添加β-甘露聚糖酶,1~21 日龄肉鸡的脾脏指数、法氏囊指数、胸腺指数分别提高3.37%、4.20%、4.93%,一定程度上提高了机体的免疫力。
3 β-甘露聚糖酶在动物生产中的应用
常规饲料原料中均含有β-甘露聚糖,β-甘露聚糖在豆粕中含量最高,为1.1%~1.3%[36]。在饲料中添加豆粕能够降低饲料成本,但甘露聚糖酶的缺乏使畜禽无法充分利用甘露聚糖。因此,在畜禽饲料中添加β-甘露聚糖酶可以提高饲料的利用率,改善动物健康[37]。
3.1 β-甘露聚糖酶在家禽生产中的应用
刘远斌等[38]在肉鸡低能量饲粮中分别添加36、72、108 U/g 的耐热β-甘露聚糖酶,与对照组相比,72 U/g添加组肉鸡回肠隐窝深度降低22.65%,干物质、粗蛋白、粗脂肪和能量的表观代谢率分别提高6.84%、23.00%、5.04%、6.33%,表明低能量饲粮中添加适量的β-甘露聚糖酶对肉鸡的小肠形态和表观代谢率均有改善作用。张荣春等[39]研究发现,在适当降低代谢能水平的肉鸡饲粮中添加100 g/t 的β-甘露聚糖酶,料重比有所降低,肉仔鸡的成活率和日增重有所提高,试验组比对照组多盈利0.51 元/只,提高了养殖效益。郑允志等[40]研究发现,在夏季高温时期,向低能饲粮(代谢能11.34 MJ/kg,粗蛋白质水平15%)中添加500 mg/kg 的β-甘露聚糖酶,蛋鸡的代谢能显著提高2.41%,蛋壳强度也有所改善,但对高温热应激引发的蛋鸡生产性能无缓解作用。万新颖等[41]研究3 种酶制剂(复合酶、葡萄糖氧化酶、β-甘露聚糖酶)对肉鸡生长性能的影响,发现β-甘露聚糖酶组肉鸡平均日增重提高了9.38%,料重比降低了8.86%,经济效益提高了22.86%;与其他试验组相比,β-甘露聚糖酶对肉鸡生长性能和经济效益的影响效果最好。辛禹秀等[42]在对不同水平(50、70、100 g/kg)β-甘露聚糖酶的肉鸡饲粮的研究中发现,与对照组相比,70 g/kg β-甘露聚糖酶添加组显著提高了肉鸡的日增重和体重均匀度,同时料重比显著降低了5.43%。黄李蓉等[43]在产蛋后期鹌鹑的基础饲粮中添加120 mg/kg 的β-甘露聚糖酶,结果发现在不影响其生产性能的前提下具有降低软蛋率的趋势。Carvalho 等[44]在蛋鸡的饲粮中添加300 g/t 的β-甘露聚糖酶并观察母鸡的行为变化,发现与对照组相比,添加β-甘露聚糖酶能够使蛋鸡的取食频率显著提高49%,减少蛋鸡的啄食和颈部损伤,因此β-甘露聚糖酶成为改善禽类福利的有效策略。Yaqoob 等[45]研究发现,在肉鸡饲粮中添加600 mg/kg 的β-甘露聚糖酶对生长性能和屠体品质无负面影响,但肉仔鸡回肠干物质和粗纤维的消化率分别提高7.68%和14.48%,表明β-甘露聚糖酶能够提高肉仔鸡对营养物质的消化率。
3.2 β-甘露聚糖酶在猪生产中的应用
薛瑞婷等[26]在玉米-豆粕型基础饲粮中分别添加0、150、300、450 U/kg 的β-甘露聚糖酶,发现随着β-甘露聚糖酶添加量增加,猪的日增重分别提高了4.59%、6.53%、3.42%,同时也提高了猪对粗蛋白质、干物质、钙、磷、中性洗涤纤维的表观消化率以及生长性能。余璐璐等[46]在仿生消化试验和断奶仔猪生产试验中发现,当酸性与中性β-甘露聚糖酶酶活比为50∶50,断奶仔猪饲粮中的添加量为6 000 U/kg 时,与对照组相比,β-甘露聚糖酶对饲粮中还原糖的释放量提高7.64%,断奶仔猪的末重、日均采食量、日增重分别显著提高了3.17%、5.51%、5.24%;同时显著提高了断奶仔猪血清中的血糖含量。朱晓彤等[47]研究发现,在豆粕添加水平为37%的断奶仔猪饲粮中添加0.02%的β-甘露聚糖酶,可显著降低断奶仔猪血清中半乳甘露聚糖的含量,显著增加前腔静脉血清中葡萄糖、钙、磷和尿素氮的含量,空肠氨基酸转运载体基因SLC7A1、SLC7A11 和SLC38A2 的相对表达量均显著升高,提高了仔猪对氨基酸的吸收利用。Kim 等[48]在基础饲粮中分别添加400、800、1 600 U/kg 的β-甘露聚糖酶,发现随着β-甘露聚糖酶浓度的增加,猪的末重、平均日增重和血糖均显著提高;当添加量为400 U/kg 时,对提高猪生产性能的作用效果最好。Jeon 等[22]研究发现,当猪基础饲粮中β-甘露聚糖酶的添加水平为0.5 g/kg 时,生长猪的料重比和粪氨浓度显著降低,猪回肠的氨基酸消化率显著提高。此外,Vangroenweghe 等[49]研究发现,添加β-甘露聚糖酶可缓解仔猪断奶后引起的腹泻。
3.3 β-甘露聚糖酶在反刍动物生产中的应用
Seo 等[50]在母牛的玉米-大豆基础饲粮和含有高甘露聚糖的饲粮中添加1 g/kg 的β-甘露聚糖酶浓缩液,发现与对照组相比,试验组母牛的平均日增重分别提高111、90 g/d,表明在母牛饲粮中添加β-甘露聚糖酶有利于提高母牛的饲料利用率和生长速度。Tewoldebrhan等[51]研究发现,在荷斯坦奶牛干物质添加量为0.1%的基础饲粮中补充0.2%的β-甘露聚糖酶,可以提高奶牛的饲料转化效率,奶牛每千克干物质产奶量提高90 g,同时降低牛乳中的体细胞计数,但对奶牛的产奶量无显著影响。Roque 等[32]研究发现,在奶牛的基础饲粮中添加0.1%的β-甘露聚糖酶,奶牛血清中结合珠蛋白水平显著降低,表明β-甘露聚糖酶在抗炎方面发挥积极作用。Jeong 等[52]研究发现,向犊牛的基础饲粮中添加0.1%的β-甘露聚糖酶能够显著提高精料的干物质质量采食量,犊牛的平均日增重与对照组相比提高148 g/d,但差异不显著。Lee 等[53]在山羊的基础饲粮中分别添加0.1%和0.3%的β-甘露聚糖酶,结果发现,与对照组相比,添加β-甘露聚糖酶组的山羊平均日增重、饲料转化率和对氮的利用率均显著提高,不同添加水平的β-甘露聚糖酶对山羊的影响效果无显著差异,表明饲料中添加β-甘露聚糖酶可以改善山羊的生长性能和对养分的消化率。
4 结论
近年来,通过添加外源酶制剂提高畜禽饲料的饲用价值受到越来越多研究者的关注。β-甘露聚糖酶能够有效降解β-甘露聚糖释放营养成分,利于营养成分的消化吸收,提高能量利用率。β-甘露聚糖的水解产物甘露寡糖能够有效改善畜禽肠道菌群环境,缓解肠道免疫应激,提高机体免疫力。将β-甘露聚糖酶用于动物生产,能够提高动物的生长性能、改善机体健康、减少畜禽粪便中氮、磷的排量。但目前我国对β-甘露聚糖酶的研究体系还不够成熟,特别在提高菌种产酶水平、评定酶制剂在饲粮中的作用效果、提高酶的耐热性和酶活力、改善产酶发酵工艺等方面仍需继续努力。同时,继续深入了解β-甘露聚糖酶的作用机制,也是未来需要重点研究的方向。综上所述,β-甘露聚糖酶作为一种新型的促生长饲料添加剂,未来具有广阔的应用前景。
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
动物营养学报(2022年8期)2022-09-01
湖南饲料(2019年5期)2019-10-15
中国化肥信息(2018年6期)2018-08-23
动物营养学报(2017年8期)2017-08-16
中国酿造(2016年12期)2016-03-01
现代农业(2016年6期)2016-02-28
动物营养学报(2015年9期)2016-01-07
中国医科大学学报(2015年10期)2015-03-01
河南科技(2014年7期)2014-02-27
