
中国牦牛种质资源及牦牛肉品质调控研究进展
2023-06-22 10:07王莉魏健李静王玉涛
甘肃农业大学学报 2023年2期
王莉,魏健,李静,王玉涛
(1. 喀什大学生命与地理科学学院,新疆帕米尔高原生物资源与生态重点实验室,新疆 喀什 844000;2. 喀什市第二中学,新疆 喀什 844000)
优异的种质资源是遗传育种及其他生命相关学科的物质基础,是国家战略性资源,事关种业振兴全局。牦牛(Bos grunniens)是分布于青藏高原及其毗邻高山、亚高山地区的特有畜种资源,能够适应高寒、缺氧、极端干燥等恶劣环境,是高原牧民生产生活重要的特有畜种[1-2]。牦牛的肉、乳、毛、粪等均是牧民主要的生产资料来源,同时在山区物资托运及固疆守边等方面扮演重要角色,其对生产区域的生态价值、经济价值和社会价值不可替代[3]。前期已有学者对我国牦牛种质资源及其肉品质改善方式的研究成果进行了总结。近年来,我国在牦牛优质地方品种遗传资源调查、申报、审定和鉴定方面又有了很大进展,同时在消费升级背景下,对牦牛肉品质要求不断提升,推动相应牦牛肉品质调控方式和相关研究不断革新升级,相关认识也更为深入和丰富。因此,本文通过对已有文献的梳理和归纳,对我国牦牛种质遗传资源现状及牦牛宰前宰后肉品质调控进展进行了较系统概述,并对牦牛肉品调控改善前景进行了讨论,以期为牦牛种质资源保护、牦牛肉品质改善提升及高质化利用和生产加工提供参考。
1 中国牦牛种质资源现状
2020 年末,我国牦牛存栏数约1 400 万头,约占世界总数的95%以上,是世界上牦牛数量和品种最多的国家,主要集中分布在青海、西藏、四川、甘肃、新疆和云南等地,少量分布于河北省北部山区[1,4-5]。2021 年1月公布的2021 版《国家畜禽遗传资源品种名录》中收录的地方牦牛品种有18个,培育品种有2个,分别是大通牦牛和阿什旦牦牛。2021年底,帕米尔牦牛和查吾拉牦牛作为10大新发现的优异畜禽遗传资源被列入畜禽遗传资源名单。帕米尔牦牛分布在新疆维吾尔自治区帕米尔高原东部海拔超过4 000 m的荒漠草场,中心产区主要在克孜勒苏柯尔克孜自治州(克州)及喀什地区塔什库尔干塔吉克自治县,现存栏约18.4 万头,其中克州12.8 万头、喀什地区5.6 万头[6]。查吾拉牦牛主要分布在西藏那曲海拔4 700 m左右的高原地区,聂荣县下曲乡和查当乡是查吾拉牦牛的中心产区,存栏约7.23 万头[7]。目前,被列入《国家畜禽遗传资源品种名录》的地方牦牛品种共20 个,具体分布及存栏情况见表1。此外,根据牦牛各品种间的遗传多样性,运用随机扩增片段长度多态性(RAPD)和微卫星(SSR)标记及牦牛线粒体基因组(mtDNA)D-loop区基因序列分析,支持将地方牦牛品种划分为青藏高原型和横断高山型2个类型的观点,但大多数研究涉及品种数量少,缺乏系统性,每个类型所包括的具体品种结果不尽相同,至今仍无定论[8]。

表1 中国牦牛地方品种分布及存栏情况[4-5]Table 1 Distribution and stock of local yak breeds in China
2 牦牛肉品质调控
目前,消费者对肉产品的关注点正逐渐聚焦于质量安全、营养价值、生产体系以及来源信息等非价格因素。牦牛常年生活于高海拔无污染天然牧场,主要采食天然优质牧草,因此牦牛肉具有“优质绿色肉食品”的独特优势[9]。牦牛肉蛋白质含量较高(约21.6%),脂肪含量较低(1.6%~4.7%),8 种人体必需的氨基酸的含量占总氨基酸含量的35%以上,n-3系列功能性脂肪酸含量显著高于秦川牛、西门塔尔牛等黄牛品种,富含维生素和矿物质元素,尤其Fe含量远高于黄牛肉,保水性能好、熟肉率高[10-20]。牦牛肉的这些独特优点深受消费者亲睐,不过,牦牛肉也存在着明显缺陷,比如多不饱和脂肪酸和饱和脂肪酸比值偏小、饱和脂肪酸和n-6/n-3值较高,脂肪沉积效率和大理石花纹评分低,嫩度、肉色稳定性和总体感官较差等,同时传统放牧牦牛生产模式存在周期长、收益低、草场压力大等问题,导致加工及食用途径受到一定限制,缺乏市场竞争力[11,21-24]。因此,进一步探究提升牦牛肉品质调控措施已成为牧民、科研人员和消费者重点关注的问题,目前主要通过宰前遗传育种改良调控和营养调控及宰后物理化学调控三种途径对牦牛肉品质进行改善提升。
2.1 遗传育种改良调控
遗传性状是影响畜禽肌肉品质的重要因素,牦牛育种改良及肉质优良品种培育是提高生产性能和改善牦牛肉品质的有效途径和发展方向[25-27]。目前,我国已有大通牦牛和阿什旦牦牛2个完全自主知识产权的人工培育肉用型牦牛新品种。大通牦牛和阿什旦牦牛分别于2005年和2019年通过国家鉴定,是世界上仅有的2个人工培育牦牛新品种,大通牦牛是在家牦牛群体(母本)中导入野牦牛(父本)优良遗传基因,适度利用近交进行闭锁四级繁育体系培育的含1/2野牦牛基因的杂种牦牛。阿什旦牦牛主要采用群体继代选育法,以更适合舍饲化养殖管理的无角牦牛为父母本,通过完善的四级繁育体系育成。这2 种牦牛均具有生长发育快(18 月龄体质量比当地牦牛提高24%以上),繁殖率高(比当地同龄牦牛提前1~1.5 岁投入繁殖,繁殖成活率为60%~85%)、产肉性能高(成年牦牛屠宰率为47%~50%),抗逆性强(越冬死亡率小于1%)等特点[28-29]。Zhang等[30]认为,大通牦牛肌间脂肪酸构成优于当地黄牛,营养价值更高。侯丽等[20]研究发现,与成年环湖牦牛、青南牦牛和秦川牛相比,大通成年牦牛肉中除色氨酸、天冬氨酸和脯氨酸外其他各种氨基酸、必需氨基酸(6.061 g/100 g)、油酸(41.05±1.41)%和亚油酸(23.00%以上)的含量均比较高。李维红等[31]和席斌等[32]对大通牦牛肉和青海高原牦牛肉的氨基酸进行比较,发现大通牦牛肉的必需氨基酸比例更为合理。冶成君[21]指出,大通犊牦牛肉的肌纤维较细,肉色和保水性较好,开发利用价值高,这与罗毅皓等[22]、刘勇[33]和张永辉[34]的研究结果相似。吴晓云等[35]发现,4 岁龄阿什旦牦牛经170 d 舍饲育肥后,背最长肌pH45min值、亮度L*24h、黄度b*24h、熟肉率和剪切力均显著降低,食用品质明显改善。当前对大通牦牛肉质性状研究较多,但对新品种阿什旦牦牛肉品质及与其他品种牦牛肉品质的相关对比分析较少。
此外,有很多学者对其他品种牦牛的肉质性状相关功能基因也在不断深入发掘、筛选和验证,为牦牛遗传育种奠定基础,以实现肉质优良品种选育。如表2所示,刘自增和阎萍[36]研究发现,除了硬脂酰辅酶A 脱氢酶(SCD1)基因会影响天祝白牦牛背最长肌脂肪酸去饱和作用外,瘦素(Leptin)基因外显子2 和3 上7 个SNPs 位点中4 个错义突变也会对脂肪酸的组成产生影响。同年刘自增[37]在甘南、大通和天祝牦牛背最长肌肥胖基因(OB)与SCD1中获得13 处影响肌肉脂肪酸含量的SNPs 位点。曹健等[38-39]分析脂肪型脂肪酸结合蛋白(A-FABP和HFABP)基因多态性与甘南牦牛肉质性状的关联表明,4.5~5岁龄A-FABP基因的AA型嫩度极显著高于BB 型,H-FABP基因的AA 型熟肉率显著高于AB 型,A-FABP和H-FABP基因突变可作为牦牛肉质性状潜在分子标记位点。焦斐[40]通过对甘南、天祝和大通牦牛编码一磷酸腺苷激活蛋白激酶γ3亚基的PRKAG3基因和脂肪酸合成酶(FASN)基因多态位点与肉质性状关联分析发现,PRKAG3基因P1位点在第3内含子发现A→G 突变,AA 型个体的失水率显著高于AB型,pH24h值显著低于AB型;P4位点在第9 内含子发现G→A 突变,PQ 型个体的失水率显著高于PP型,而熟肉率、pH24h值、剪切力和脂肪含量差异不显著。FASN基因F3B 位点M 等位基因、F34B位点上H等位基因和F35位点上E等位基因均对牦牛肉脂肪含量起正面效应。褚敏等[41]在研究FASN基因第3 内含子SNPs 位点与牦牛部分肉质性状相关性时也得出了类似的结论。秦文[42]发现,甘南牦牛背最长肌FASN基因mRNA表达量与C10:0 呈极显著正相关。陈海青等[43-44]、潘红梅[45]、牛晓亮[46]和陈杰[47]研究发现,细胞骨架蛋白(ANK1)基因、钙蛋白酶(CAPN)基因家族中的CAPN1、CAPN3、CAPN4和钙蛋白酶抑制蛋白(CAST)基因的多个SNPs 位点均与甘南牦牛肉的嫩度和保水性显著相关,且相关性受年龄影响。Temizkan等[48]指出,ANK1对牛肉嫩度的影响较大,且ANK1与肉质性状的关联性较钙蛋白酶1(CAPN1)基因更强。根据官久强等[49]和张海波等[50]的研究,通过抑制激素敏感脂肪酶(HSL)和肉碱转移酶-1(CPT-1)基因的表达,可促进麦洼牦牛肌内脂肪沉积,提高嫩度。石斌刚[51]以不同月龄天祝白牦牛为研究对象,通过转录组、蛋白质组学和荧光定量等方法分析获得了FASN、FABP3、FABP4等24个与肌内脂肪沉积相关的基因和蛋白。上述研究均是从遗传角度筛选与牦牛肉质性状相关的候选基因,可为后续牦牛选育改良、遗传分子育种等工作提供参考,但对牦牛肉品质关键调控基因、品质性状与各基因之间的关联及各基因的网络调控关系的研究还不够深入,相关基因的功能和调控机制仍然需要进一步探索和精确验证。
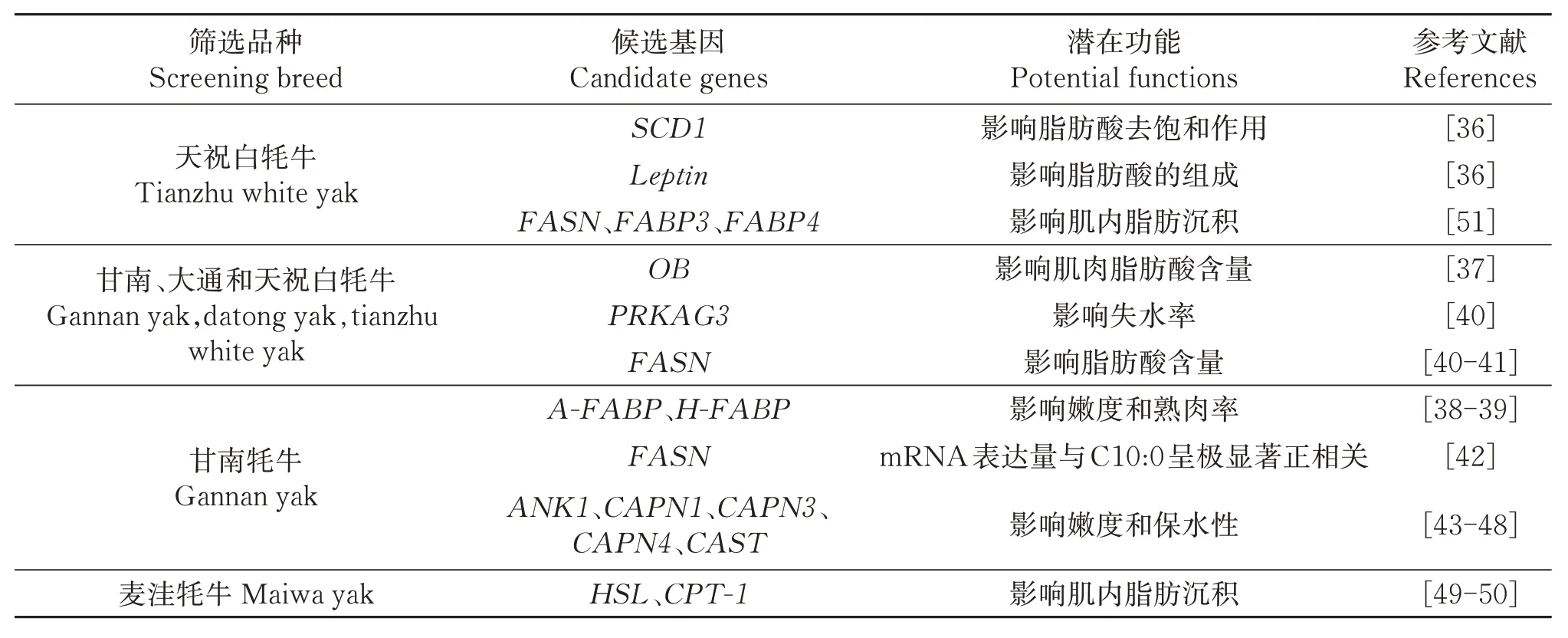
表2 牦牛肉质性状相关候选基因及其潜在功能Table 2 Candidate genes and potential functions of yak meat quality
2.2 营养调控
通过营养调控改善牦牛肉品质已成为牦牛高效生产过程中的共识。目前营养调控肉品质主要是在冷季或暖季对牦牛进行补饲或舍饲,根据牦牛生长期能量代谢、氮代谢及其他营养需要,通过适宜添加形式和添加量,向牦牛日粮中增加能量水平、蛋白质、脂肪、矿物质、氨基酸和生物活性物等营养物质,以达到牦牛高效生产及改善肉质的目的[52-54]。姚喜喜等[55]研究发现,在饲喂80%全贮玉米和20%小麦秸秆的基础上,以玉米、麸皮、菜粕、小苏打、盐、预混料为主配制净能6.94 MJ/kg 的日粮水平,经15 d 预饲期和90 d 正试期后,能显著提高舍饲大通牦牛的眼肌面积和pH值,但会使牦牛肉色L*值降低。陈光吉[56]以麦洼牦牛为研究对象,以玉米秸秆和鲜酒糟为日粮粗料,以玉米、麦麸、豆粕、发酵酒糟、碳酸钙、磷酸氢钙、硫酸钠、小苏打、食盐、氯化胆碱、矿舔及多维1为精料配制综合净能水平(预饲期7 d、正试期90 d),发现不同能量水平日粮对麦洼牦牛肉的pH值、保水性、常规营养成分及氨基酸含量均无显著影响,但净能4.48 MJ/kg 的日粮水平可以提高牦牛肉的不饱和脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸相对含量。这与周立业[57]、孙鹏飞[58]和方雷[59]的研究结果相似,但与杨媛丽[60]和郝力壮[61]研究结果有差异,后两位学者均认为,对放养牦牛进行舍饲或补饲能显著提高牦牛肉的食用品质和营养品质,同时会调节参与牦牛机体蛋白质和氨基酸代谢、脂肪酸代谢、碳水化合物代谢、维生素代谢、矿物质代谢及免疫代谢途径等功能相关的代谢通路。马志远[62]在牦牛饲料中添加不同水平的过瘤胃氨基酸和矿物质缓释丸,预饲期7 d,正试期90 d 后,牦牛肉的肉色,滴水损失及剪切力均有改善。王鸿泽[63]发现,增加环湖牦牛的日粮能量水平会促进背最长肌脂肪合成基因、脂肪转运基因及减缓脂肪分解基因mRNA的表达,促进肌内脂肪沉积,进而改善嫩度、保水性、风味及营养成分的组成和含量。由以上研究结果可知,通过营养调控方式可改善牦牛肉的食用品质,但对肉的常规营养成分影响程度不尽相同,需进一步结合肉品质研究相关调控机理,制定相应补饲标准,在保留牦牛肉现有优点的基础上,提升牦牛肉品质。
2.3 宰后物理化学调控
刚屠宰完的牦牛肉最明显的缺陷是嫩度差、肉色不稳定、风味不佳,而嫩度、肉色和风味均是影响牦牛肉品质最直观重要的感官因素,既会影响消费者的接受性和购买欲,又会降低牦牛肉的食用价值、商品价值和加工利用性。因此,如何改善牦牛肉的嫩度、维持肉色稳定性及提高牦牛肉的风味是当前宰后调控改善牦牛肉品质的研究热点。宰后牦牛肉嫩度可通过物理和化学两类方式调控改善。物理嫩化法主要有成熟、电刺激和振动处理,化学嫩化法主要是通过向牦牛肉中添加钙盐、外源酶和磷酸盐等改善嫩度。Zhang 等[64]将牦牛背最长肌在4 ℃成熟21 d 后,剪切力值显著降低了60%。成熟期间嫩度的变化与肌原纤维小片化、结构弹性网状蛋白含量及内源蛋白酶的降解作用等有关[10]。郑祖林等[65]对宰后天祝白牦牛胴体立即进行低压电流刺激发现,电刺激处理能明显加速牦牛肉成熟期间糖酵解速度,使pH急速下降,肌原纤维小片化程度增大,剪切力值显著降低,使嫩度、多汁性及风味得到改善,特别对品质较差的后半躯部位肉效果尤为显著。Lang等[66]发现,电刺激结合成熟能显著提高冷热剔骨甘南牦牛肉的嫩度。这可能是电刺激加速宰后牦牛肉ATP 和糖原消耗,加快pH 下降速率,使钙蛋白酶(calpain)被更早激活,进而加快骨骼肌肌原纤维骨架蛋白—肌钙蛋白-T(Troponin-T)和肌间线蛋(Desmin)降解,破坏肌纤维结构,提高嫩化速率,达到改善肉品质的目的[67-70]。杨璐[71]利用加氯盐振动技术对牦牛肉进行嫩化处理,发现加盐振动处理可显著破坏牦牛背最长肌肌原纤维结构,加大肌原纤维水解程度,嫩化效果明显。马秀利[72]用CaCl2溶液处理宰后牦牛背最长肌发现,Ca2+可加快肌肉能量代谢活动,使糖酵解加速,降低肌肉内环境pH值,激活并提高成熟前期钙激活酶和细胞凋亡酶活性,加快成熟速度,使牦牛肉在72 h内完成成熟,剪切力值(4.24 kgf)不足非CaCl2组(8.65 kgf)的一半,达到较好的嫩度和口感,这与高永芳[73]、Wang 等[74-75]的研究结果相似。可能与单磷酸腺苷活化蛋白激酶(AMPK)及其级联激活效应相关[76-77]。陈一萌等[78]将超声波、木瓜蛋白酶、超声辅助木瓜蛋白酶、发酵、超声辅助木瓜蛋白酶+发酵这5种牦牛肉嫩化方法进行了比较,5个处理组的牦牛肉肌原纤维小片化指数均显著高于对照组,硬度、弹性、内聚性和咀嚼性显著低于对照组,其中超声波辅助木瓜蛋白酶组最低。杨敏等[79]用磷酸盐处理牦牛肉发现,磷酸盐对牦牛肉的嫩度也有改善作用,但嫩化效果较CaCl2差。目前在牦牛肉嫩度改善方面就宰后内源酶对肌纤维组织结构及肌原纤维骨架蛋白的作用研究较为深入,而对宰后牦牛肉肌内结缔组织特性、变化及调控改善方式缺乏系统和深入研究。
牦牛宰后成熟除了能改善肉的嫩度外,还可对肉色和肉的风味产生影响。王琳琳等[80]发现,肌肉部位和肌纤维类型均可显著影响麦洼牦牛肉成熟过程中的肉色稳定性。陈骋[81]将甘南牦牛背最长肌在4 ℃成熟7 d后,发现L*值、a*值和氧合肌红蛋白相对含量分别显著降低8.22%、34.53%、33.66%,H*值和高铁肌红蛋白相对含量分别显著升高37.73%、315.89%,肉色主要特征为红色度降低,肉色劣变的根本原因是肌红蛋白氧化和线粒体高铁肌红蛋白还原能力降低,同时伴随着脂质氧化程度的升高,脂质氧化产物己醛和壬醛是影响肉色稳定性的主要因子,而内源抗氧化物质α-生育酚和肌肽能够抑制微粒体脂质氧化、降低肌红蛋白氧化速率。马骋[82]研究发现,宰后牦牛肉成熟期间,用60%O2和40%CO2气调包装,肉色较稳定,还原型辅酶(NADH)和乳酸脱氢酶(LDH)是影响肉色的酶促因素。陈炼红等[83]指出,通过增加NADH、糖原含量、LDH活性和降低乳酸含量可抑制单磷酸腺苷激活蛋白激酶(AMPK)活性,而抑制AMPK活性有利于维持牦牛背最长肌在成熟期间肉色的稳定性。已有研究表明,宰后成熟有利于牦牛肉风味的改善,李永鹏等[84]、师希雄等[85]和牛珺等[86]采用固相微萃取(HSSPME)和气相色谱-质谱(GC-MS)联用技术检测分析甘南牦牛肉和西藏斯布牦牛肉成熟过程中的风味物质,发现成熟过程中总体风味物质增多,且宰后成熟可增强了牦牛肉的肉香味,减弱牦牛肉的脂肪味和牧草中特殊成分对牦牛肉风味的不良影响。
3 结论与展望
我国牦牛种质资源丰富,经济潜力巨大,但牦牛肉在嫩度、肉色和脂肪沉积等方面存在一定缺陷,这些因素在一定程度上限制了牦牛肉产业发展,通过科学的肉品调控方式可提升牦牛肉品质、增强市场竞争力。目前虽然对宰前遗传育种改良、营养调控和宰后物理化学调控独立改善牦牛肉的品质有较深入的认识和研究,但对牦牛肉品质宰前和宰后的系统关联调控研究较少,应结合宰后牦牛肉品质调控方式,对牦牛肉质关键调控基因与品质性状之间的关联开展完整系统深入研究,以揭示各基因对牦牛营养调控及宰后肉质调控的网络关系和机理,制定相应饲养标准,达到提升牦牛肉品质的目的。
猜你喜欢
西藏人文地理(2020年4期)2020-11-19
西南民族大学学报(自然科学版)(2019年5期)2019-11-12
幼儿教育·父母孩子版(2018年12期)2018-04-20
恋爱婚姻家庭(2017年18期)2017-12-26
肉类研究(2017年10期)2017-11-20
中国农业科学(2016年17期)2016-12-01
广东饲料(2016年7期)2016-12-01
浙江农业科学(2016年11期)2016-05-04
西藏科技(2015年10期)2015-09-26
畜牧兽医学报(2015年3期)2015-07-05
