
玉米大豆间作对土壤真菌群落和作物生产力的影响
2023-06-07 03:44:30王兵兵李光文刘文娟何亚玲姜琴芳
农业科学研究 2023年1期
王兵兵, 李光文, 马 琨, 刘文娟,何亚玲, 姜琴芳, 米 扬
(1.宁夏大学 农学院,宁夏 银川 750021;2.宁夏大学 西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021)
间套作在资源高效利用、改善土壤肥力、提高作物产量和稳定作物生产力等方面具有较大潜力[1]。作为经典的间套作种植模式,禾、豆搭配具有提高资源利用率的生物学基础。Duchene 等[2]研究认为,禾本科与豆科作物间作可以提高作物生产力。禾、豆间套作能够改善土壤肥力、提高作物产量以及持续增加年际间的产量效益[3]。在禾豆间套作系统中,禾本科作物通过降低土壤氮素浓度,促进豆科作物结瘤固氮,实现了在氮素利用上的生态位分离[1]。Chen 等[4]在玉米||大豆的长期减氮定位试验中发现,因大豆自身的固氮特性、种间相互作用及间作系统对有限资源的高效利用,仍能实现高产。此外,禾、豆间作可以改善土壤养分有效性,提高作物对土壤养分的吸收利用。可见,土壤养分是引起土壤微生物群落演替的重要因素,而两者间的动态变化又影响了植物的生长发育,植物的生长状况又与土壤性质和土壤微生物直接相关[5]。因此,植物、土壤和微生物之间的相互作用应该是影响农田生态系统的主要驱动力。
土壤微生物在农业生态系统扮演着重要角色,具有维持土壤生态系统稳定和土壤可持续生产力的潜能[6]。Li 等[7]认为,间套作中作物地下交互作用能够改善土壤微生物群落组成和土壤微生物主要类群;植物多样性能增加根际对土壤微生物的碳输入,促进土壤微生物活性和碳储量增加[8];禾、豆间作能改善土壤微生态环境,提高土壤质量[9]。土壤真菌作为土壤微生物的重要组成部分,其群落组成及多样性在与植物互作、驱动碳氮循环和调节植物营养等方面发挥着独特作用[10—11]。有研究表明,植物多样性可通过增加土壤全氮提高土壤真菌丰富度[12]。Lian 等[13]发现,与单作相比,间作改变了土壤真菌群落组成且降低了土壤真菌的Alpha 多样性。但也有学者发现,间作系统能够改变土壤真菌群落的Alpha 和Beta 多样性,并认为这是由作物种间相互作用的差异性和土壤性质差异引起的[14]。可见,诸多关于土壤真菌群落、多样性变化规律及其与环境因子间驱动关系的研究结果并不完全一致。
本试验通过分析玉米大豆间作下作物生产力、土壤真菌群落组成、多样性及其与土壤性状间的相互关系,揭示间作对作物生产力以及土壤真菌群落的影响,阐明其驱动因素,以期为协调土壤资源高效利用和集约化农业可持续发展提供理论支撑。
1 材料与方法
1.1 试验地概况
试验地位于宁夏同心县王团镇(105°59′E,36°51′N),海拔1 373 m,年均降水量272.6 mm,年均蒸发量2 740 mm,年均气温8.7 ℃。供试土壤为淡灰钙土,试验前土壤w(全磷)=0.92 g/kg、w(碱解氮)=26.78 mg/kg、w(速效磷)=20.54 mg/kg、w(速效钾)=227.61 mg/kg、pH=8.50(水土比为5∶1)。
1.2 试验设计
试验开始于2019 年4 月,采用单因素随机设计,3 个处理,4 次重复,小区面积96 m2(4.8 m×20.0 m),小区间距50 cm。处理1,单作玉米(M),行距60 cm,株距30 cm;处理2,单作大豆(S),行距30 cm,株距15 cm;处理3,玉米||大豆(间作,MS),玉米和大豆均采用条带种植,其中玉米带宽120 cm,大豆带宽60 cm,玉米带和大豆带间隔20 cm,带宽均为1.2 m,玉米、大豆行比为3∶3,行株距与对应单作相同。供试玉米品种为翔玉998,大豆品种为承豆6 号。玉米、大豆于当年4 月中旬同期播种,10 月1 日前后同期收获。2019 年、2020 年播种前均将磷酸二铵、复合肥(wN=14%、wP2O5=16%、wK2O=15%)和尿素(w总氮≥46.0%)一次性基施,用量分别为20、20、10 kg/666.7 m2,全生育期不再追肥。以滴灌方式在玉米苗期、拔节期、大喇叭口期和开花期各灌水1 次。
1.3 样品采集及处理
于玉米大喇叭口期、抽雄初期、灌浆期及成熟期等主要生育阶段,在每小区选取3 株长势一致的玉米、大豆植株,在105 ℃杀青30 min,随后在80 ℃烘干至恒质量,用于测定地上干物质积累量。于作物收获期将小区全部大豆去荚称质量;在单作玉米和间作玉米中随机选取3 行称质量,计算作物产量。连续试验2 年后,于作物收获期(2020 年10 月)采用5点取样法,采集不同处理0~10、11~20 cm 土层土壤样品。其中,玉米||大豆从2 种作物条带的行间采集,每小区采集相同土层5 份土壤样品并将其混合均匀,共采集独立土壤样品24 份。部分土壤样品低温储存带回实验室,过1 mm 筛,储存温度2~8 ℃,用于测定土壤硝态氮、铵态氮;一部分储存在-80 ℃冰箱,用于测定土壤真菌多样性;另一部分土壤样品阴凉风干,用于测定土壤理化性质。
1.4 研究方法
1.4.1 土壤理化性状的测定 采用元素分析仪(Elementar V MAX)测定土壤全氮、总碳;土壤全磷、速效磷、碱解氮、速效钾、含水量的测定分别采用H2SO4-HClO4消煮-钼锑抗比色法、NaHCO4浸提-钼锑抗比色法、碱解扩散法、NH4OAc 浸提-火焰光度法、烘干法;利用PHSJ-4F pH 计(水土比为5∶1)测定土壤pH[15];土壤硝态氮和铵态氮的测定采用1 mol/L KCl 浸提-流动注射分析仪(Futura 2)。
1.4.2 土壤样品的DNA 提取和PCR 扩增 采用CTAB 方法提取土壤真菌DNA,使用超微量紫外分光光度计(Nanodrop,ND-1000,德国)测定土壤样品DNA 浓度。土壤真菌扩增的正向引物为ITS1-1F-F(5′-CTTGGTCATTTAGAGGAAGTAA-3′),反向引物为ITS1-1F-R(5′-GCTGCGTTCTTCATCGATGC-3′)。PCR 扩增参数:98 ℃预变性1 min;98 ℃变性10 s,55 ℃退火30 s,72 ℃延伸30 s,最后72 ℃延伸5 min,循环30 次。PCR 反应体系为30 μL,2×Phusion Master Mix 15 μL,2 μmol/L Prime 3 μL,模板DNA 10 μL,ddH2O 2 μL。基于Illumina NovaSeq 测序平台对PCR 扩增产物进行双末端测序(Paired-End),委托天津诺禾致源生物信息科技有限公司完成。
1.5 数据分析与计算
1.5.1 数据处理 原始下机数据经Reads 拼接、Tags 过滤、质量控制和去嵌合体后得到有效数据。然后基于有效数据,利用Uparse 软件(Version 7.0.1001)将各样本按97%相似性序列聚类为操作分类单元(operational taxonomic units,OTUs),以其中出现频率最高的序列作为OTUs 代表序列。利用QIIME软件(Version 1.9.1)和Unit 数据库(Version 8.2)对各样本中OTUs 代表序列进行物种注释,使用QIIME 软件在界、门、纲、目、科、属、种各分类水平统计各样本的物种组成及其丰度分布。使用MUSCLE软件(Version 3.8.31)进行多序列比对。使用QIIME软件进行Alpha 多样性分析;使用R 软件对Unweighted 的Unifrac 距离矩阵进行NMDS 分析;使用Metastats 的统计学算法,对属水平各样本间的物种丰度数据进行比较检验。
1.5.2 数据统计 数据整理采用Excel 2010,单因素方差分析的Duncan 多重比较采用SPSS 25.0。利用Origin 2021 和Canoco 5.0 绘图。
2 结果与分析
2.1 玉米大豆间作对作物地上干物质积累和产量的影响
由图1 可知,随着作物生育期推移,不同种植方式下玉米地上干物质积累量均呈增加趋势,间作玉米地上干物质积累量均高于对应单作玉米;大豆地上干物质积累量却呈现出先增加后减少的变化趋势。与单作大豆相比,2019 年在大豆生长前期(6 月11 日),间作大豆的地上干物质积累量显著增加了27.87%;在中后期单作大豆地上干物质积累量却高于间作大豆。2019—2020 年,间作下的玉米产量比单作玉米分别高26.10%、30.56%,差异达显著水平(P<0.01);与单作相比,间作大豆产量极显著降低了54.33%、58.08%。相比2019 年,2020 年单作大豆、间作玉米和单作玉米的产量均有所提高,但年际间差异不显著。

图1 玉米||大豆对作物地上干物质积累及产量的影响
2.2 玉米大豆间作对不同土层土壤真菌群落组成的影响
土壤真菌参与有机质分解、腐殖质合成以及土壤碳氮循环等[16]。0~10 cm 土层(图2a),土壤赤霉菌属(Gibberella)和核瑚菌属(Typhula)真菌为优势属,其相对丰度所占比例为40.75%~52.53%。与单作玉米相比,玉米||大豆提高了链格孢属(Alternaria)(P=0.692)、光黑壳属(Preussia)(P=0.063)、未分类(P=0.054)和丝状菌属(Titaea)相对丰度,降低了镰孢菌属(Fusarium)和附球霉属(Epicoccum)等真菌的相对丰度(图2a),其中镰孢菌属(P=0.242)和附球霉属(P=0.724)分别降低了61.96%和43.19%;与单作大豆相比,玉米||大豆提高了假雨盘菌属(Pseudombrophila)(P=0.941)和核瑚菌属(P=0.372)相对丰度,但总体上不同处理间的差异不显著。

图2 玉米||大豆对0~10 cm(a)和11~20 cm(b)土层土壤真菌群落组成的影响
11~20 cm 土层中链格孢属、粪锈伞科-未分类(Bolbitiaceae-U)和枝孢属(Cladosporium)真菌为优势属(图2b),所占比例为34.65%~77.16%。与单作玉米相比,玉米||大豆显著提高了粪锈伞科-未分类(P=0.043)相对丰度,降低了链格孢属、枝孢属和附球霉属等相对丰度,其中链格孢属(P=0.001)达极显著水平;与单作大豆相比,玉米||大豆系统显著降低了枝孢属(P=0.043)相对丰度。单作和间作种植方式对11~20 cm 土层其他属真菌相对丰度的影响,在不同处理间未达显著水平。
赤霉菌属、链格孢属、假雨盘菌属、光黑壳属、附球霉属和镰孢菌属是同一种植方式下不同土层前10 属中的共有真菌属;随土层深度增加,共有真菌属相对丰度均有所降低,其中单作玉米中的链格孢属(P=0.033)、单作大豆中的赤霉菌属(P=0.012)及玉米||大豆系统中的光黑壳属(P=0.015)达显著水平。不同土层属水平真菌群落组成及其相对丰度存在差异,说明随土层深度改变,基于属水平的土壤真菌群落组成也会发生改变。
2.3 玉米大豆间作对不同土层土壤真菌群落Alpha多样性的影响
由图3 可知,3 种处理下土壤0~10 cm 土层真菌多样性指数(Shannon 和Simpson)大小依次为S、MS、M,而丰富度指数(Chao1 和ACE)大小依次为S、M、MS,并且单作大豆Chao1 指数显著高于单作玉米。不同种植模式下11~20 cm 土层的多样性指数(Shannon 和Simpson)大小依次为M、MS、S,而丰富度指数(Chao1 和ACE)大小依次为M、S、MS,且单作玉米和单作大豆的Chao1(P<0.01)和ACE(P<0.01)指数极显著高于玉米||大豆系统。在单作大豆中,随土层深度增加,多样性指数(Shannon 和Simpson)和Chao1 指数均显著降低;玉米||大豆处理0~10 cm 土层的ACE 指数(P<0.05)显著高于11~20 cm 土层;单作玉米下不同土层的多样性指数和丰富度指数也存在差异,但未达显著水平。


图3 玉米||大豆对不同土层土壤真菌群落Alpha多样性的影响
2.4 玉米大豆间作对不同土层土壤真菌群落Beta多样性的影响
非度量多维尺度(NMDS)分析表明,不同种植方式下基于属水平的土壤真菌群落组成在空间分布上存在一定差异(图4)。与玉米||大豆相比,单作玉米下土壤真菌群落样本组内差异性较小;在0~10 cm(图4a)和11~20 cm(图4b)土层中,单作玉米与玉米||大豆的空间位置明显分开。这说明玉米||大豆影响了不同土层深度土壤真菌的群落组成。单作大豆下,土壤真菌群落Beta 多样性在空间上与单作玉米、玉米||大豆均重叠在一起,空间位置无明显分异。ANOSIM 相似性分析表明,在0~10 cm 和11~20 cm土层,M 与S 组间均无显著差异;尽管M 与MS 及S 与MS 的组间差异大于组内差异,但组间均未达显著水平。
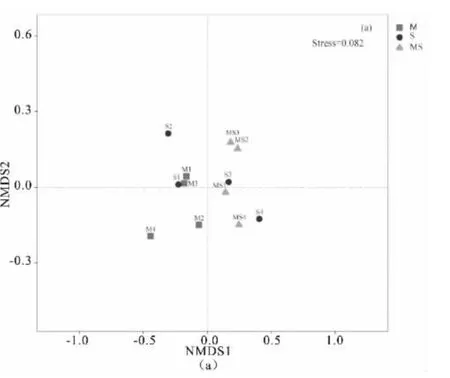
图4 玉米、大豆单作及间作下0~10 cm(a)和11~20 cm(b)土层真菌群落组成的NMDS 分析
2.5 玉米大豆间作下不同土层土壤真菌群落组成及Alpha 多样性与土壤理化性状间的相互关系
RDA 分析表明,第1、2 排序轴能够分别在累积变量23.28%、21.85%上揭示0~10 cm 土层土壤理化性状与土壤真菌群落组成及Alpha 多样性间的相关性(图5a)。利用向前选择,经蒙特卡洛置换检验发现,连续栽培2 年后,土壤C/N(P=0.029)和TC(P=0.039)是影响0~10 cm 土层土壤真菌群落组成及Alpha 多样性的主要因子,解释率分别达15.6%和14.5%。土壤真菌链格孢属、附球霉属、丝状菌属、光黑壳属相对丰度、Chao1 指数和多样性指数(Shannon 和Simpson)与C/N 呈正相关,与TC呈负相关;假雨盘菌属、未鉴定类、核瑚菌属的相对丰度和ACE 指数与TC 呈正相关,与C/N 呈负相关。

图5 玉米、大豆单作及间作下0~10 cm(a)和11~20 cm(b)土层真菌群落组成、Alpha 多样性与土壤理化性状间的RDA 分析
通过对11~20 cm 土层土壤理化性状与土壤真菌群落组成及Alpha 多样性间的RDA 分析发现,第1、2 排序轴能分别在累积变量24.80%、21.30%上揭示11~20 cm 土层土壤理化性状与土壤真菌群落组成及Alpha 多样性间的相关性(图5b)。AN(P=0.001)是影响11~20 cm 土层土壤真菌群落组成及Alpha 多样性的主要环境因子,解释率为20.5%。AN与赤霉菌属、光黑壳属、假雨盘菌属、枝孢属、接合地衣属、链格孢属、附球霉属的相对丰度和ACE 指数呈正相关,与侧盘菌科-未分类、镰孢菌属、粪锈伞科-未分类的相对丰度、Shannon 指数、Simpson 指数及Chao1 指数呈负相关。此外,不同土层C/N 与ACE 指数呈负相关,与Chao1 指数、Shannon 指数和Simpson 指数呈正相关,而NO3--N与之相反。
3 讨论
3.1 玉米大豆间作对作物生产力的影响
有关研究发现,禾、豆间作既能增强地上复合群体光截获及利用能力,也能通过地下种间协同作用提高土壤养分利用效率[7,17]。试验结果表明,间作玉米产量显著高于单作,单作大豆产量极显著高于间作。Liu[5]等认为,玉米与大豆间作优势是地上、地下相互作用的结果,这种优势可能源于土壤性状的改善和植物有益土壤微生物群落的构建。试验中,在作物生育期内,玉米、大豆干物质积累量呈上升趋势;不同生育期内间作玉米干物质积累量均高于单作玉米;前期间作大豆的生物量高于单作大豆,但生物量在中、后期的变化趋势与此相反。有研究表明,玉米大豆套作系统中,玉米和大豆不同时期干物质积累量均呈先快后慢的增加趋势[18];与单作相比,间作玉米和间作大豆均有产量优势[19],这与本试验结果不完全一致。分析认为,玉米大豆间作下,玉米在其拔节期后比大豆有更强的光截获能力和更大的生长空间,从而玉米成为优势作物;由于玉米对大豆的遮阳效果会伴随其以后的生育阶段,这使得间作大豆干物质积累量的增加受到抑制,进而影响其产量。
Wang[20]等认为,玉米||大豆系统中的大豆能为间作系统提供氮源,从而减弱了作物种间对氮素的竞争,利于玉米生长。本试验也发现,间作系统中土壤NO3--N 含量显著低于单作玉米(P<0.05),且玉米大豆间作中11~20 cm 土层土壤AN 含量比单作玉米降低了32.90%(P<0.05),分析认为,间作下作物的种间相互作用促进了植物对氮素的利用能力。Li 等[21]研究也证实,间作系统比单作具有更高的养分吸收量,且物种间的需肥特性存在差异,能够在时间生态位上形成互补[22—23]。因此,间作可通过充分利用物种间的协同作用,促进土壤养分吸收,提高间作系统生产力。此外,禾、豆间作系统中玉米根系通过分泌柠檬酸、黄酮类化合物和染料木素[24—25]等物质促进豆科根瘤菌固氮也可能是一些原因。本研究表明,与单作玉米相比,间作使得11~20 cm 土层土壤AP 含量提高了40.56%(P<0.05),Wang 等[26]也在玉米大豆间作中发现,禾、豆间作系统能够提高土壤中有效磷含量,进而改善玉米磷营养。分析认为,玉米||大豆系统中玉米群体植株较高,对光热资源截获能力强,且玉米、大豆物种间的协同作用和根生态位的分离,是土壤养分利用效率和玉米生产力提高的主要原因,高生产力的间作系统必然会从土壤中带出更多的土壤养分。但是,Ryan[27]认为适宜的作物间作能够增加产量、作物稳定性和土壤肥力。因此,土壤肥力的改善在提高作物产量方面也有积极作用。
3.2 玉米大豆间作对土壤真菌群落结构的影响
试验中,玉米大豆间作下不同土层间土壤真菌组成和相对丰度存在差异。在0~10 cm 土层赤霉菌属(Gibberella)和核瑚菌属(Typhula)为优势属。在11~20 cm 土层,粪伞科-未分类(Bolbitiaceae-U)、链格孢属(Alternaria)和枝孢属(Cladosporium)为优势属。分析认为,不同土层土壤真菌优势属间的差异与所处环境为其提供的碳源种类有关。Challacombe等[28]证实,子囊菌门有分解纤维素和其他植物碳水化合物的能力,是驱动生态系统碳氮循环的重要参与者;由于间作玉米对间作大豆的遮阳作用,在生育后期大豆叶片因早衰提前脱落,为表层土壤微生物提供了更丰富的碳源。这可能就是其对应分类水平下土壤真菌以较高丰度存在于表层土壤的原因。试验中,玉米||大豆系统中作物物种间的根系结构和空间分布差异以及间作系统引起的根系分泌物和土壤性质的变化,可能就是引起不同土层土壤真菌群落组成优势属出现差异的主要原因。此外,与单作玉米相比,间作系统能够显著提高粪锈伞科-未分类(P=0.043)相对丰度,降低赤霉菌属、镰孢菌属和核瑚菌属等病原菌的相对丰度。作物收获期,土壤主要真菌类群中含有较多的病原菌,这可能是由于玉米||大豆系统中大豆早衰及玉米代谢减缓引起的,要揭示其具体原因还有待进一步分析间作下不同作物生育时期根系分泌物组成和土壤真菌群落组成。
Eisenhauer[29]研究发现,植物多样性是决定土壤微生物多样性的关键因素,并且土壤微生物多样性随植物多样性的增加而增加。但本试验中,与单作玉米相比,间作系统提高了0~10 cm 土层的Shannon 指数和Simpson 指数,而降低了11~20 cm 土层真菌多样性指数和丰富度指数,其中丰富度指数(Chao1 和ACE)(P<0.01)达极显著水平。在玉米大豆间作下,随土层深度增加,ACE 指数显著降低;在单作大豆下, 随土层深度增加, 多样性指数(Shannon 和Simpson)和Chao1 指数显著降低。植物根系分泌物因根系生态位和发育阶段而异,且根系分泌物可以改变根周围的土壤性质和土壤微生物群落组成[30—32]。
Zheng[33]证实,土壤微生物易受环境干扰,其群落结构的改变会影响土壤碳氮循环及其他养分的有效性,进而调控植物营养供给。本试验中,TC、C/N 和AN 是驱动土壤真菌群落结构发生变化的主要土壤因子,这与前人研究结果一致[34]。此外,Li 等[35]发现,土壤有机碳的化学组分与土壤真菌群落组成密切相关,且土壤有机碳组分含量的差异是改变不同土层土壤真菌群落组成的主要原因,Li 等还发现土壤真菌群落组成和有机碳组分在表层土壤中更敏感。本试验中连续2 年的间作与单作栽培对土壤TC、C/N并未产生显著影响。分析认为,单作玉米、单作大豆枯落物及间作系统中因玉米遮阳作用而早衰脱落的大豆叶片能为土壤真菌提供丰富碳源,但枯落物的种类、性质及作物种间交互作用均会影响土壤中碳源存在的形态。可见,不同单作和间作下土壤碳源的变化及差异有可能是引起土壤真菌群落结构及多样性变化的原因。Zhang 等[36]也证实,间作系统能够显著提高苜蓿根际土壤TN、TC、AN 和AK 含量,改善土壤真菌多样性和丰富度。此外,也有研究指出,土壤有机质、铵态氮、速效磷和有效氮含量是影响土壤真菌群落结构的主要因素[37]。由此可知,种植方式引起的土壤理化性状的变化,可能是驱动土壤真菌群落组成及其多样性变化的另一重要因素。
4 结论
玉米大豆间作提高了玉米产量而降低了大豆产量。与单作玉米相比,玉米大豆间作能够降低土壤赤霉菌属、镰孢菌属和核瑚菌属等病原菌的相对丰度,显著提高11~20 cm 土层粪锈伞科-未分类真菌的相对丰度而极显著降低链格孢属相对丰度。土壤TC和C/N 是驱动0~10 cm 土层土壤真菌群落组成和Alpha 多样性变化的主要土壤因子,而在11~20 cm土层土壤真菌群落结构的改变主要是由土壤AN差异引起的。
猜你喜欢
经济林研究(2022年4期)2023-01-12 09:14:46
广西林业科学(2022年2期)2022-05-09 07:49:54
中草药(2021年19期)2021-10-12 03:22:08
河北果树(2020年4期)2020-11-26 06:05:18
山东农业科学(2019年11期)2019-12-24 01:11:27
江苏农业科学(2019年11期)2019-07-22 01:32:45
天然产物研究与开发(2018年7期)2018-08-21 02:04:02
干旱地区农业研究(2017年5期)2017-12-18 06:18:18
新疆农垦科技(2016年2期)2016-08-21 13:50:18
西藏大学学报(自然科学版)(2015年1期)2015-12-29 07:37:22
