
冰封期乌梁素海浮游植物生态位和种间联结性研究
2023-04-29 09:05:50林岩丁晓宇吕航卢兴顺李佳赵子闻武琳慧
水生态学杂志 2023年3期
林岩 丁晓宇 吕航 卢兴顺 李佳 赵子闻 武琳慧


摘要:为了解乌梁素海冰封期浮游植物群落结构和生态位特征,于2018年和2019年冬季(1月)进行浮游植物采样调查。根据乌梁素海污染源分布、水文及环境特征,在湖泊入水口(N1~N6)、湖泊中部(N7~N14)和湖泊出水口(N15、N16)选取了16个典型样点,进行浮游植物和水样采集;运用改进的Levins公式和Petraitis指数测定浮游植物优势种的生态位宽度和生态位重叠度。结果表明,2018年和2019年冰封期共鉴定出浮游植物优势种2门6种和6门8种,伪鱼腥藻是2018年的绝对优势种,2019年的绝对优势种为蓝隐藻和旋转囊裸藻。2018年冰封期优势种生态位宽度为0.0711~0.5971,宽度最大的为球衣藻,其次是绿柄球藻和衣藻;2019年冰封期优势种生态位宽度为0.1265~0.5455,宽度最大的为绿柄球藻,其次是具尾逗隐藻和蓝隐藻。2018 年和2019年乌梁素海冰封期各优势种的生态位重叠值分别为0.0078~0.7967 和0.0448~0.9248,整体上偏小;2018年冰封期浮游植物优势种绿藻门的衣藻与球衣藻的重叠值最高,2019年隐藻门的具尾逗隐藻与蓝隐藻的重叠值最高。生态位重叠值和生态响应速率显示,2018年乌梁素海冰封期浮游植物群落整体上呈发展趋势,而2019年呈衰退趋势。方差比率法分析显示,优势种整体呈正关联;卡方检验发现,各优势种间的联结性均不显著,大多数藻类种间为负关联,少数为正关联。
关键词:浮游植物;优势种;生态位;种间联结性;乌梁素海
中图分类号:X172 文献标志码:A 文章编号:1674-3075(2023)03-0102-08
最早在生态学中使用生态位概念是为了划分环境的空间单位和物种在环境中的地位(Grinnell, 1917)。一个世纪以来,许多学者对生态位理论进行探索研究和补充完善,已成为现代生态学研究中不可忽视的一部分,其在理解群落结构功能、群落物种间关系、生物多样性、群落动态演替和种群进化方面有重要作用(田雅楠和艾尼瓦尔,2021)。
生态位及种间联结性已经在陆地动植物生态学领域中得到广泛应用(江焕等,2019;赵家豪等,2021);水域生态系统中主要应用于底栖动物和浮游动物群落(张琳琳等,2019;安瑞志等,2021),而浮游植物的相关研究较少(魏志兵等,2020;马一明等,2021)。浮游植物是水域生态系统中的初级生产者,对水体多种理化和生物因素的变化反应灵敏,常作为水环境评价的依据和指标(吴小伟和刘平,2015)。将湖泊中浮游植物生态位理论和种间联结关系结合起来,有助于了解浮游植物之间的相互关系,反映湖泊浮游植物群落的稳定性和发展趋势,对湖泊富营养化监控和治理具有指导意义。
冰封期是寒区湖泊水质变化的重要时期。乌梁素海冰封期长,结冰后湖泊热交换过程、污染物分布及冰下底栖温度环境都将发生改变,从而影响湖中浮游植物的组成与结构(杨芳等,2016)。目前,已有关于各类湖泊中浮游藻类生态位特征的报道(李卫平等,2014,李兴等,2018),但对于高原冰封期湖泊藻类生态位的研究较为匮乏。本文通过生态位宽度和生态位重叠度相关理论,分析乌梁素海冰封期浮游植物生态位和种间联结度,旨在揭示浮游植物群落结构和分布特征,以期对湖泊营养状态评估与污染治理提供基础数据支持。
1 材料与方法
1.1 区域概况及样点布设
乌梁素海位于内蒙古自治区乌拉特前旗,属于典型的浅水草型湖泊(吕超,2013)。湖泊除了明水区外,还有大量以芦苇为主的挺水植物区。乌梁素海是黄河改道形成的河迹湖,现今主要靠乌加河和长济渠、民复渠等灌溉尾水补给。作为黄河流域最大的淡水湖,其具有净化水质、调节水量、防止河套灌区土壤盐碱化、减缓水体富营养化进程等多种生态功能,对调节河套地区生态平衡有重要作用(姜慧琴,2011)。
按照我国水生生态系统调查规范,根据乌梁素海污染源分布、水文及环境特征(李兴等,2012),选取了16个典型样点;其中,N1~N6为湖泊入水口,N7~N14为湖泊中部,N15和N16为湖泊出水口。具体位置如图1所示。
1.2 样品采集及处理
于2018年和2019年1月的乌梁素海冰封期进行采样。采用冰钻采集器破冰,浮游植物定性样品采集使用25号浮游生物网(60 μm)捞取,水样在现场分别使用5%的甲醛和1%的鲁哥试剂固定。浮游藻类种类鉴定和计数参考相关文献(金相灿和屠清瑛,1990;周凤霞和陈剑虹,2011)。
1.3 数据处理
1.3.1 优势度指数 依据McNaughton(1967)优势度指数(Y)确定优势种,计算公式如下:
式中:a为2个物种组成的种对同时存在的样点之和,b和c为其中仅出现一个物种的样点之和,d为一对物种共同出现频率为0的样点数,因 X2计算结果无法反映负值,利用ad-bc的符号对物种间的正、负联结性进行判断,若其值为正数,可判定为正联结,反之则判定为负联结,相等则判定为无联结。
在种间联结性中,AC能体现出那些由X2检验判断为不显著种对的联结性及大小(张金屯,2004):
式中:a、b、c 值同卡方检验;AC值域为[-1, 1],越接近1,说明物种间的正联结性越强;越接近-1,说明负联结性越强;AC=0说明物种间完全独立。
2 结果与分析
2.1 浮游植物群落结构及优势种分布
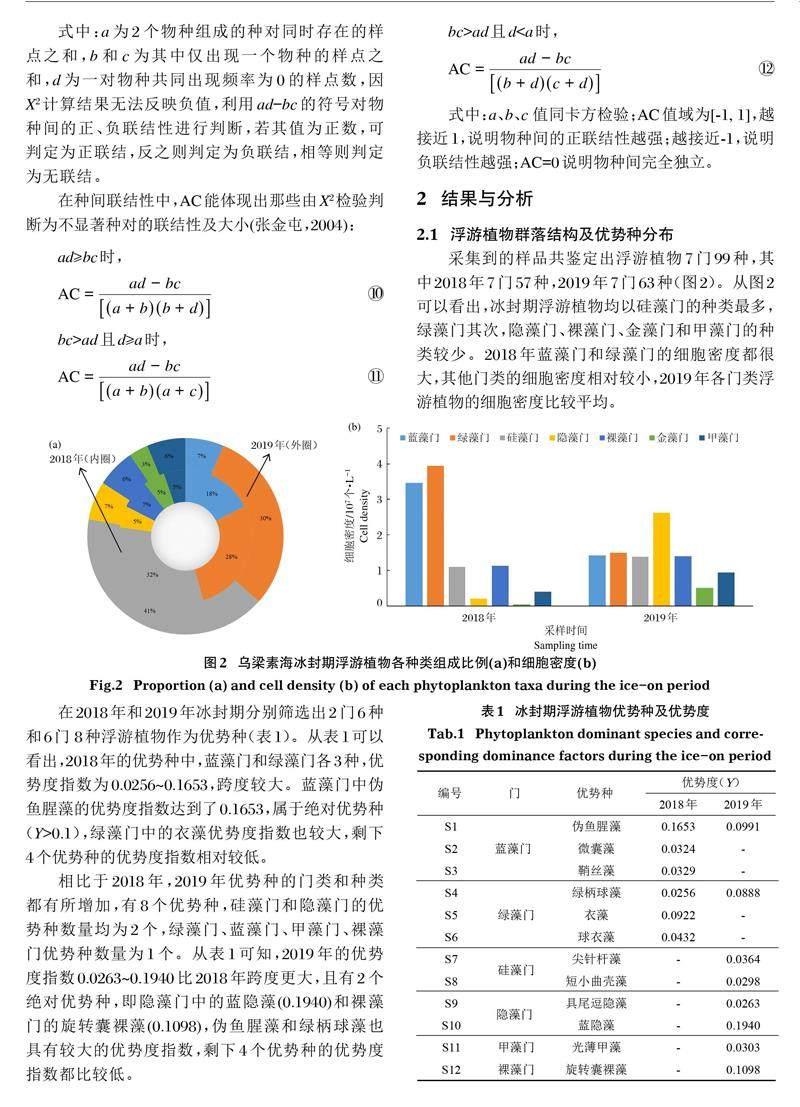
采集到的样品共鉴定出浮游植物7门99种,其中2018年7门57种,2019年7门63种(图2)。从图2可以看出,冰封期浮游植物均以硅藻门的种类最多,绿藻门其次,隐藻门、裸藻门、金藻门和甲藻门的种类较少。2018年蓝藻门和绿藻门的细胞密度都很大,其他门类的细胞密度相对较小,2019年各门类浮游植物的细胞密度比较平均。
在2018年和2019年冰封期分别筛选出2门6种和6门 8种浮游植物作为优势种(表1)。从表1可以看出,2018年的优势种中,蓝藻门和绿藻门各3种,优势度指数为0.0256~0.1653,跨度较大。蓝藻门中伪鱼腥藻的优势度指数达到了0.1653,属于绝对优势种(Y>0.1),绿藻门中的衣藻优势度指数也较大,剩下4个优势种的优势度指数相对较低。
相比于2018年,2019年优势种的门类和种类都有所增加,有8个优势种,硅藻门和隐藻门的优势种数量均为2个,绿藻门、蓝藻门、甲藻门、裸藻门优势种数量为1个。从表1可知,2019年的优势度指数0.0263~0.1940比2018年跨度更大,且有2个绝对优势种,即隐藻门中的蓝隐藻(0.1940)和裸藻门的旋转囊裸藻(0.1098),伪鱼腥藻和绿柄球藻也具有较大的优势度指数,剩下4个优势种的优势度指数都比较低。
2.2 浮游植物优势种的生态位
2.2.1 生态位宽度 乌梁素海冰封期优势种的生态位宽度如表2所示。2018年优势种生态位宽度在0.0711~0.5971,且除了蓝藻门的鞘丝藻,都有较大的生态位宽度,最大的为球衣藻(0.5971)。2019年的优势种生态位宽度范围与2018年相差不大,在0.1265~0.5455,绿柄球藻的生态位宽度变化也较小,但新增隐藻门的优势种具尾逗隐藻和蓝隐藻都有较大的生态位宽度。
2.2.2 生态位重叠值 由表3和表4可知,2018年和2019年乌梁素海冰封期各优势种的生态重叠值分别在0.0078~0.7967和0.0448~0.9248。2018年球衣藻(S6)与衣藻(S5)的生态重叠值最高,为0.7967,鞘丝藻(S3)和衣藻(S5)的生态重叠值最低,仅为0.0078。2019年以隐藻门中的具尾逗隐藻(S9)与蓝隐藻(S10)的生态重叠值最高,为0.9248,接近完全重叠;短小曲壳藻(S8)与旋转囊裸藻(S12)的生态重叠值最小,仅为0.0448。
结合生态位宽度计算得到的生态响应速率(R)如表5所示。由表5可知,2018年鞘丝藻的生态重叠指数(ΔOik)达到-8.7798,表明冰封期的衰退程度很大,但R=-0.0081,说明继续衰退的趋势较弱。剩下5种优势种的生态重叠值都是正值,说明其在冰封期是发展型,伪鱼腥藻、微囊藻、球衣藻、绿柄球藻发展趋势较强,衣藻发展趋势稍弱。
2019年冰封期的优势种伪鱼腥藻(-1.2167)、尖针杆藻(-7.5959)、短小曲壳藻(-13.2815)、旋转囊裸藻(-4.8855)生态重叠值是负值,均属于衰退型,其中短小曲壳藻衰退程度最大;与同鞘丝藻一样,短小曲壳藻(R=-0.0095)的衰退趋势较弱,而伪鱼腥藻(R=-0.2452)的衰退趋势较为强烈。绿柄球藻(7.3545,R=0.0741)、光薄甲藻(3.6657,R=0.0561)、具尾逗隐藻(6.9561,R=0.0697)、蓝隐藻(9.0033,R=0.0554)的ΔOik都是正值,说明这4种优势种在冰封期发展型,但发展趋势较弱;蓝隐藻的ΔOik在4种优势种中最大,说明其在冰封期对环境的适应能力最好,对资源利用率最大。从整体上看,2018年冰封期浮游植物生态响应速率之和为正数(1.0204),说明浮游植物整体上呈发展趋势;而2019年浮游植物的生态响应速率之和为负数(-0.0950),说明其整体上呈较弱衰退趋势。
2.2.3 种间联结性 通过计算得到2018年和2019年的V为4.0787和8.2643,说明2年间乌梁素海冰封期浮游植物的总体联结性属于正联结,且2019年的联结性比2018年强(表6,表7)。2018年6个优势种之间只有球衣藻(S6)和衣藻(S5)的X2统计量大于6.635,表明这2个优势种之间的联结性极强。通过比较15个种对之间ad-bc的正负性,可以判断有5对优势种为正联结,占总种对数的33.3%;有10对优势种为负联结,占66.7%。联结系数也表明(图3,图4),5个种对为正联结,10个种对为负联结,有6个种对的AC指数小于-0.5,说明这6个种对负联结性较强。
3 讨论
3.1 浮游植物优势种表明乌梁素海营养状态好转
蓝-绿藻型湖泊的水质营养化程度较高,优势种的种类越多,则说明水体中的群落结构越复杂,群落结构越稳定(柳丽华等,2007;赵秀侠等,2021)。2018年冰封期的优势种只有2门,浮游植物的细胞密度都远超其他门,说明乌梁素海水体处于富营养化状态或趋于富营养化状态,群落结构简单;而2019年的优势种门类较多,有6种,各门细胞密度占比比较平均,且占优势的是硅藻门和隐藻门,说明2019年冰封期乌梁素海的富营养状态有所好转,群落结构较为稳定。
3.2 浮游植物环境适应力与生态位宽度和重叠性
生态位宽度反映了各优势种对资源的利用程度和对环境的适应能力(汪志聪等,2010)。2018年冰封期绿藻门优势种的生态位宽度比蓝藻门大,说明绿藻门中的优势种占据了较大的空间且在资源竞争中处于优势地位。2019年生态位宽度最大的是绿藻门的绿柄球藻,其次是隐藻门的蓝隐藻和具尾逗隐藻。证明乌梁素海冰封期隐藻门在竞争资源中处于优势地位,也占据了较大的生存空间,这也和优势种的研究结论一致。
Wathne等(2000)认为ΔOik>0.6优势种的重叠性较显著。本次研究中,2018年共有5对优势种显示了较强重叠度,其中球衣藻和衣藻之间有最大的生态重叠值,这可能与二者都属于绿藻门,其生活习性和资源利用相似有关,因此其生态位重叠显著,这与马一明等(2021)的研究结论一致。而鞘丝藻和其他所有优势种的生态重叠值都很低,说明其生活习性与其他优势种都不相似,且生态位宽度也很窄,表明其对环境变化比较敏感,只有特定的环境适宜其生长。2019年28个种对中,只有4对生态重叠值大于0.6,说明各优势种之间在空间和资源利用上有所区别,这也与2018年优势种门类多相呼应。隐藻门的蓝隐藻和具尾逗隐藻具有最大的生态位宽度,且生态重叠值非常接近1,说明这2种藻类在冰封期的空间分布大致相同且更多地利用同种资源,生态位分化不明显,这可能导致资源利用竞争较为激烈,种间竞争强度较大。
3.3 营养盐高低影响浮游植物优势种变化态势
当i=k时,通过比较ΔOik大小来说明不同的环境条件下不同种属的生态习性变化和发展衰退状况。差值的正负表明该种属处于发展还是衰退状态,差值的大小反映了该种发展或衰退的程度,为0时则表明该种为中性种(夏霆等,2014)。结合生态响应速率之和为正数(1.0204),2018年的优势种整体上是发展的,只有鞘丝藻衰退且程度很大,这可能是由于其对环境的要求较高,环境发生改变就会导致鞘丝藻的减少。2019年生态响应速率之和为负数(-0.0951)说明浮游植物群落整体上处于衰退阶段。冰封期气温低,湖泊逐渐结冰后形成的冰层阻碍了水体与大气的复氧过程,减弱了冰下浮游植物的光合作用,形成了较为极端的生境(何婷婷,2017),优势种的ΔOik有正有负,说明乌梁素海冰封期浮游植物群落结构发生了改变,一些不适应低温的藻类生长发育受到影响,数量变少;另一些耐低温或者对温度和溶解氧需求不高的浮游植物,在其他藻类减少的情况下,对空间和资源的占有率得到提高。2018年和2019年的优势种趋势分别为发展和衰退,造成这种情况的原因是2018年的总氮和总磷含量较高,优势种的营养元素充足;而2019年的总氮和总磷含量较低,资源匮乏,优势种对营养元素的竞争较为激烈,导致整体上优势种的衰退。
3.4 乌梁素海浮游植物的种间联结性总体不显著
X2检验能定性分析物种的关联性,进而反映出乌梁素海冰封期浮游植物各个优势物种间的联结性。2018年只有球衣藻和衣藻具有极显著的联结性,结合联结系数,两者为极显著的正联结性关系,其余优势种种间无极显著和显著关联的种对。2019年的8个优势种的种间均无显著关联,结合生态重叠值总体较小,说明这些种对的优势种相似的生态习性少。联结系数表明,2018年和2019年各个种对多数表现出较强的负联结性,说明冰封期浮游植物竞争激烈,群落结构属于演替阶段初期,而一般认为种对间正联结越多,群落结构越稳定(周先叶和王伯荪,2000)。总体来看,主要浮游植物之间以负联结关系为主,种对之间联结关系不显著。但种对间正联结关系越强,生态位宽度重叠程度并不一定越高,这可能与种对的分布频率、生境选择和对资源的利用能力不同等因素有关(丁朋朋等,2019)。
参考文献
安瑞志,张鹏,达珍,等,2021. 西藏麦地卡湿地不同水文期原生动物优势种生态位及其种间联结性[J]. 林业科学, 57(2):126-138.
丁朋朋,高春霞,彭欣,等,2019. 浙江南部近海主要虾类的时空生态位及种间联结性[J]. 应用生态学报, 30(11):3942-3950.
何婷婷,2017. 寒区湖泊冰封期藻类及污染物冰——水介质分布特征及响应关系研究[D]. 呼和浩特:内蒙古师范大学.
江焕,张辉,龙文兴,等,2019. 金钟藤入侵群落的种间联结及生态位特征[J]. 生物多样性, 27(4):388-399.
姜慧琴,2011. 乌梁素海营养盐在冰体中的空间分布及其在冻融过程中释放规律的试验研究[D]. 呼和浩特:内蒙古农业大学.
金相灿,屠清瑛,1990. 湖泊富营养化调查规范[M]. 北京:中国环境科学出版社.
李卫平,徐静,于玲红,等,2014. 乌梁素海冰封期营养盐及浮游植物的分布特征[J]. 生态环境学报, 23(6):1007-1013.
李兴,何婷婷,勾芒芒,2018. 乌梁素海冰封期浮游植物群落特征与环境因子CCA分析[J]. 东北农业大学学报, 49(4): 67-78.
李兴,张树礼,李畅游,等,2012. 乌梁素海浮游植物群落特征分析[J]. 生态环境学报, 21(11):1865-1869.
柳丽华,左涛,陈瑞盛,等,2007. 2004年秋季长江口海域浮游植物的群落结构和多样性[J]. 海洋水产研究, (3):112-119.
吕超,2013. 乌梁素海冰封期富营养化特征研究[D]. 呼和浩特:内蒙古科技大学.
马一明,李秋华,潘少朴,等,2021. 贵州高原花溪水库浮游植物优势种生态位及种间联结性动态分析[J]. 湖泊科学, 33(3):785-796.
田雅楠,艾尼瓦尔·吐米尔,2021. 新疆博格达峰北坡岩面生壳状地衣的生态位特征[J]. 干旱区资源与环境, 35(1):108-113.
汪志聪,吴卫菊,左明,等,2010. 巢湖浮游植物群落生态位的研究[J]. 长江流域资源与环境, 19(6):685-691.
魏志兵,柴毅,罗静波,等,2020. 长湖浮游植物优势种季节演替及生态位分析[J]. 水生生物学报, 44(3):612-621.
吴小伟,刘平,2015. 扬州境内湖泊浮游植物群落结构及其与环境因子的关系[J]. 水资源保护, 31(5):47-52,71.
夏霆,陈静,曹方意,等,2014. 镇江通江城市河道浮游植物优势种群生态位分析[J]. 长江流域资源与环境, 23(3):344-350.
杨芳,李畅游,史小红,等,2016. 乌梁素海冰封期湖泊冰盖组构特征对污染物分布的影响[J]. 湖泊科学, 28(2):455-462.
张金屯,2004. 数量生态学[M]. 北京:科学出版社.
张琳琳,蒋日进,印瑞,等,2019. 乐清湾主要游泳动物空间生态位及其分化[J]. 应用生态学报, 30(11):3911-3920.
赵家豪,叶钰倩,陈斌,等,2021. 江西武夷山南方铁杉针阔混交林主要植物生态位特征[J]. 林业科学, 57(1):191-199.
赵秀侠,卢文轩,梁阳阳,等,2021. 安徽三座通江湖泊秋季浮游植物群落结构特征及其影响因子[J]. 生态学杂志, 40(1):67-75.
周凤霞,陈剑虹,2011. 淡水微型生物与底栖动物图谱[M]. 北京:化学工业出版社.
周先叶,王伯荪,2000. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性[J]. 植物生态学报, 24(3):332-339.
Colwell R K, Futuyma D J, 1971. On the measurement of niche breadth and overlap[J]. Ecology, 52(4):567-576.
Grinnell J, 1917. The Niche-Relationships of the California Thrasher[J]. The Auk, DOI:10.2307/4072271
McNaughton S J, 1967. Relationship among functional prosperities of California grassland[J]. Nature, 216:168-169.
Pianka E R, 1973. The structure of lizard communities[J]. Annual Review of Ecology and Systematics, 4:53-74.
Schluter D, 1984. A variance test for detecting species associations, with some example applications[J]. Ecology, 65(3): 998-1005.
Wathne J A, Haug T, Lydersen C, 2000. Prey preference andniche overlap of ringed seals Phoca hispida and harpseals P. groenlandica in the Barents Sea[J]. Marine Ecology Progress Series, 194:233-239.
(责任编辑 万月华)
Ecological Niche and Interspecific Association of Phytoplankton during
the Ice-on Period of Ulansuhai Lake
LIN Yan1, DING Xiao‐yu1, LV Hang1, LU Xing‐shun1, LI Jia1, ZHAO Zi‐wen1, WU Lin‐hui1,2
(1. College of Ecology and Environment, Inner Mongolia University, Hohhot 010021, P.R. China;
2. Key Laboratory of Environmental Pollution Control and Waste Recycling
in Inner Mongolia Autonomous Region, Hohhot 010021, P.R. China)
Abstract:In this study, we explored the phytoplankton community structure and ecological niche characteristics of Ulansuhai Lake during the ice-on period and analyzed interspecific associations of phytoplankton based on theoretical conceptions of ecological niche breadth and overlap. Our objective was to provide basic data to support nutrient evaluation and pollution treatment of the lake. Based on the distribution of pollution sources, hydrology and environmental characteristics of Ulansuhai Lake, 16 representative sites were selected, including the inlet (N1-N6), open area (N7-N14) and outlet (N15, N16) of the lake for sampling phytoplankton and water in January (winter) of 2018 and 2019. The Improved Levins formula and Petraitis Index were used to determine niche breadth and niche overlap of dominant phytoplankton species. A total of 99 phytoplankton species from 7 phyla were identified during the two investigations, with 57 species from 7 phyla in 2018 and 63 species from 7 phyla in 2019. Among them, six species from two phyla in 2018 and 8 species from 6 phyla in 2019 were dominant during the ice-on period, with the absolute dominance by Pseudoanabaena sp. in 2018 and the absolute dominance by Chroomonas sp. and Trachelomonas volvocina in 2019. The niche width of the dominant species in the ice-on period was 0.0711-0.5971 in 2018, and Chlamydomonas globosa had the largest niche width, followed by Stylosphaeridium stipitatum and Chlamydomonas sp. In 2019, the niche width of the dominant species was in the range of 0.1265-0.5455, and Stylosphaeridium stipitatum had the largest niche width, followed by Komma caudata and Chroomonas sp.. In 2018 and 2019, the niche overlap values of the dominant species were 0.0078-0.7967 and 0.0448-0.9248, respectively, and the niche overlap values were generally small. The overlap degree of Chlamydomonas sp. and Chlamydomonas globosa was highest in 2018, and the overlap degree of Komma caudata and Chroomonas sp. was highest in 2019. The overlap of ecological niches and the rate of ecological response indicates that phytoplankton community showed an overall developing trend in 2018 and a declining trend in 2019. The analysis of the variance ratio method showed that, overall, the dominant species were positively correlated. The chi-square test indicated that the association between dominant species was not significant, and most algae species were negatively correlated, with a few species displaying positive correlations.
Key words: phytoplankton; dominant species; niche; interspecific association; Ulansuhai Lake
收稿日期:2021-09-17 修回日期:2023-01-13
基金项目:国家自然科学基金(42067037,41701281);内蒙古“一湖两海”科技重大专项(ZDZX2018054)。
作者简介:林岩,1997年生,男,硕士研究生,研究方向为环境微生物学。E-mail:imulinyan1997@163.com
通信作者:武琳慧,1982年生,女,博士,研究方向为环境微生物学。E-mail:imuwulinhui@163.com
猜你喜欢
中国三峡(2017年4期)2017-06-06 10:44:22
科教导刊·电子版(2016年28期)2017-01-10 21:27:23
商业经济研究(2016年22期)2016-12-27 10:48:00
农家科技下旬刊(2016年9期)2016-12-15 13:23:52
时代金融(2016年27期)2016-11-25 17:07:08
水利科技与经济(2016年8期)2016-04-22 03:41:12
河北渔业(2015年11期)2015-11-18 16:37:49
河北渔业(2015年10期)2015-10-15 11:14:57
湖北农业科学(2015年18期)2015-10-13 21:36:01
科技视界(2015年25期)2015-09-01 15:49:44
