
大渡河乐山段鱼类群落结构及其多样性分布特征
2023-04-29 00:50:35王文君方艳红杨钟黄晋何春华杜霞
水生态学杂志 2023年3期
王文君 方艳红 杨钟 黄晋 何春华 杜霞


摘要:了解安谷水电站建设后大渡河乐山段鱼类群落结构与多样性现状特征及变化,探讨鱼类群落对安谷水电站工程建设的响应,可为河流生态系统保护和恢复提供基础信息。2016-2018年利用地笼、多网目复合刺网等渔具对大渡河乐山段鱼类资源进行调查。结果表明,调查期间共采集到鱼类10 751尾,隶属于5目12科45属54种;其中,鲤科鱼类最多,有27种,占总种数的50.00%。水电站库区段、坝下段、左侧生态河道段共采集到鱼类36、50、35种,优势种分别为6、7、7种。根据鱼类个体生态学矩阵分析,大渡河乐山段鱼类以杂食性、砂砾底栖型、产粘沉性卵类群为主,安谷水电站坝下河段喜流水性鱼类更多;鱼类种类数和群落Shannon-Wiener多样性指数(H')、Pielou均匀度指数(J')、Margalef丰富度指数(R)均以安谷水电站坝下段最高,分别为50种和3.2033、0.8188、5.7234。水电站库区、坝下、生态河道渔获物体长在4~10 cm的数量分别占各河段总渔获物的69.17%、76.11%、78.36%,鱼类群落个体小型化、低质化明显,表明大渡河乐山段渔业资源呈现衰退,其主要原因是受捕捞强度过大及水电工程开发建设的影响,提出了恢复鱼类群落多样性的相关措施。
关键词:鱼类群落;分布特征;多样性;安谷水电站
中图分类号:Q145 文献标志码:A 文章编号:1674-3075(2023)03-0062-08
大渡河是长江上游的二级支流,岷江右岸的最大支流,干流全长1 062 km,天然落差4 175 m,流域面积77 400 km2(不含青衣江);其水能资源丰富,理论蕴藏量达33 679.7 MW,技术可开发量24 009 MW,经济可开发量17 791 MW,年发电量985.78亿kW·h,是四川省水能资源最丰富的三大河流之一,也是我国规划的十二大水电基地之一。
大渡河铜街子以下为下游,属四川盆地边缘,为丘陵宽谷区,河谷开阔,沙洲和岔流极为发育,特别是沙湾至乐山段,长约35 km,水流散乱,洲岛遍布,是典型的多汊滩险河道;该河段于2015年8月建成安谷水电站,是大渡河干流梯级开发的最后一级。安谷水电站具有发电、防洪、航运、灌溉和供水等综合利用功能,兼顾区域湿地与生态保护,属大(二)型水电站,正常蓄水位398 m,总利用落差36 m,电站总装机容量为772 MW,年均发电量32.93亿kW·h,为日调节运行水库。
安谷水电站建设改变了大坝下游的水流、水温、泥沙与生源要素的流动、运移模式等,影响着河流生物群落系统的结构和动态平衡,鱼类作为水生态系统的顶级类群,在河流生态系统中占据着重要位置(黎道丰和蔡庆华,2000);同时,鱼类群落通过上行和下行效应与其生存环境密切相关(刘恩生,2007)。当栖息环境发生变化时,鱼类群落组成及分布等在时间和空间上也将发生变化,鱼类群落与生存环境的耦合关系是目前国际淡水群落生态学领域研究的焦点之一(张觉民,1991;Feyrer & Healey,2003;Garcia et al,2003;Koel & Peterka,2003;Gerhard et al,2004;Maes et al,2004;刘恩生等,2005;刘恩生等,2007;Costa et al,2007;Leitao et al,2007;Sutela & Vehanen,2008)。本文通过对大渡河下游安谷水电站建设后的鱼类群落及其多样性变化规律研究,探讨鱼类群落对水电工程建设的响应,旨在为河流生态系统保护与恢复及管理提供基础信息。
1 材料与方法
1.1 调查方法
分别于2016-2018年4-5月和9-10月对大渡河下游乐山市至河口长约30 km的河段进行鱼类调查,采样方法参考《内陆水域渔业自然资源调查手册》(张觉民,1991)。通过购买渔获物和雇用渔民捕捞的方式开展鱼类样本采集,捕捞工具主要有定置刺网(网目2~10 cm,网高1~2 m,网长20~50 m)和地笼(网目1 cm)。采集到的鱼类样本现场进行种类鉴定及个体生物学数据测量,包括全长和体长(精确到0.1 cm)以及体重(精确到0.1 g)。对现场不能确定的种类,先对样本进行拍照,再用5%的甲醛溶液固定后带回实验室进一步鉴定,种类鉴定参考相关文献(湖北省水生生物研究所鱼类研究室,1976;丁瑞华,1994)。
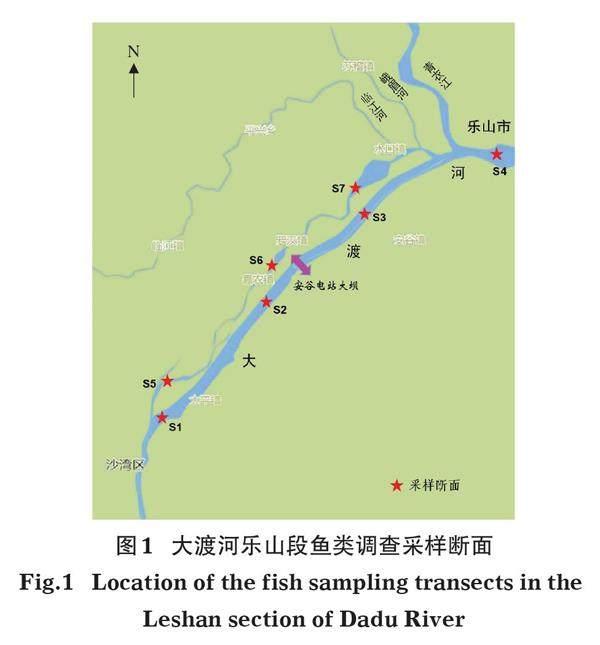
根据安谷水电站的工程特点,针对电站库区、坝下和左侧保留的生态河道段,分别布设鱼类资源采样断面(图1,表1)。其中,断面S1、S2位于安谷水电站坝上库区河段,断面S3、S4位于安谷水电站坝下河段,断面S5、S6、S7位于左侧生态河道段。
1.2 数据处理与统计分析
1.2.1 鱼类个体生态矩阵 鱼类个体生态矩阵是从鱼类生活史的特征考量,建立起鱼类与其所需栖息环境间的交互关系矩阵:
式中:S为某样点鱼类名录矩阵,S=[S1,S2,S3,…,Sj,…,Sn ],A为鱼类个体生态矩阵,ns为某样点采集到的鱼类总种类数。
R为某样点所有种类鱼类对栖息环境需求的矩阵,由[R1,R2,R3,…,Ri,…,Rn]矩阵组成;其中,Ri表示某样点中鱼类群落对某一栖息环境因子Ei的需要程度(以百分比表示)。Ri值越高,说明该鱼类受此环境因子的影响越大(蒋志刚和纪力强,1999;王丹等,2007)。
1.2.2 生物多样性指数 本文采用Shannon-Wiener多样性指数H′(Wilhm,1968;张雅芝和黄良敏,2009)、Pielou均匀度指数J′(Pielou,1975)、Margalef丰富度指数R(Margalef,1957;李博等,2000)评估鱼类群落的生物多样性,计算公式如下;
式中:Wi为群落中第i种鱼类的重量,W为群落中鱼类的总重量,S为群落中鱼类总种类数,N为群落中鱼类总尾数。
1.2.3 鱼类群落优势种 采用Pinkas(1971)相对重要性指数(Index of relative importance,IRI)确定鱼类群落的优势种类,计算公式如下:
IRI = (N+W)×F ⑤
式中:N为群落中某种鱼类尾数与总尾数的百分比值,W为群落中某种鱼类重量与总重量的百分比值,F为群落中某种鱼类出现的采样点数与总采样点数的百分比值。本文将IRI≥500的种类认定为优势种(程济生和俞连福,2004;熊飞等,2015)。
数据统计分析利用Excel 2019与软件SPSS 24.0完成。
2 结果与分析
2.1 鱼类群落种类组成
4次调查共采集到鱼类54种,分属于5目12科45属(表1)。其中,鲤形目4科34属39种,占总种数的72.22%;鲇形目4科7属10种,占18.52%;鲈形目2科2属3种,占5.56%;合鳃目和鳉形目均仅采集到1科1属1种。在科级水平以鲤科种类最多,有27种,占总种数的50.00%,符合中国淡水鱼类主要以鲤科鱼类为主的组成特点(张春光等,2016);其次是鳅科和鲿科,均采集到7种,分别占12.96%。安谷水电站库区段共采集到鱼类36种,坝下段50种,左侧生态河道段35种。
2.2 鱼类生态类型
根据大渡河乐山段采集到的鱼类对不同水流类型、饵料资源、栖息水层、底质条件等栖息环境的需求,筛选出11个环境因子,建立起由594个生态矩阵因子组成的鱼类个体生态矩阵(表2)。结果表明,安谷水电站库区与左侧生态河道段鱼类均以适宜静缓流水域、杂食性、底栖生活、喜砂砾底质且产粘沉性卵的鱼类为主,坝下段鱼类以更适宜流水生境、杂食性、底栖生活、喜砂砾底质且产粘沉性卵的鱼类为主。总体上看,安谷水电站坝下段鱼类对栖息环境的需求较电站库区和生态河道段更高。
2.3 渔获物特征及优势种组成
IRI指数统计结果显示(表3),安谷水电站库区段优势种为蛇鮈、乐山小鰾鮈、鲤、棒花鱼、鲫、唇?共6种,优势种数量占渔获物总数量的83.17%,占渔获物总重的83.12%;坝下段优势种为短体副鳅、峨嵋后平鳅、白缘?、福建纹胸鮡、蛇鮈、山鳅、鲤共7种,优势种数量占比为85.03%,重量占比为45.37%;生态河道段优势种为宽鳍鱲、蛇鮈、乐山小鰾鮈、鲫、山鳅、钝吻棒花鱼、棒花鱼共7种,优势种数量占比为72.21%,重量占比为60.09%。
库区段渔获物优势种除了鲤以外,其他优势种平均体长不超过20 cm,平均体重不超过100 g,均为小型鱼类;坝下段渔获物优势种除了鲤以外,其他优势种平均体长不超过12 cm,平均体重不超过15 g,均为小型鱼类;生态河道段渔获物优势种平均体长不超过11 cm,平均体重不超过18 g,均为小型鱼类。各河段渔获物均以小型鱼类占绝对优势。
安谷水电站库区、坝下与生态河道段渔获物体长在4~10 cm的数量分别占各河段渔获物总数量的69.17%、76.11%、78.36%,体长超过20 cm的渔获物数量仅占4.40%、1.28%与0.42%(图2)。调查结果表明,渔获物种类及个体小型化趋势明显。
2.4 鱼类群落生物多样性指数
本次调查以安谷水电站坝下段鱼类群落生物多样性指数(3.2033)、均匀度指数(0.8188)以及丰富度指数(5.7234)最高,电站库区段鱼类群落生物多样性指数(2.0840)、均匀度指数(0.5816)以及丰富度指数(4.3347)最低;安谷水电站坝下河段鱼类群落种类丰富且分布均匀(表4)。
3 讨论
3.1 安谷水电站建设导致河道生境片段化
安谷水电站是大渡河干流开发的最后一个梯级,位于从山区向平原过渡的大渡河河口段。工程所在河段承接着上游来水来沙,同时受下游岷江和青衣江、峨眉河的顶托,使得该河段形成了开阔的河谷,河道流速减缓,泥沙沉积,心滩、漫滩发育,汊濠纵横。根据本课题组对安谷水电站工程建设前所在河段的卫星遥感影像资料加工、解译,同时结合工程建设布局特征进行的预测分析,发现安谷水电站所在河段工程建设前的洲岛数超过70个,自然河道蜿蜒度高达4.964,河网密度达3.384 km/km2,工程坝址所处的大渡河右侧天然主河道过流断面为150~400 m,为典型的河网型河道。安谷水电站建成后,右侧河网原有河道渠化、顺直化,过水断面变宽,电站坝下形成长9 461 m、底宽91 m的尾水渠,还有与其并行的长9 205 m、底宽350~420 m泄洪渠,坝上库区过水断面500 m。水电站建设后,原有的河网结构受到严重影响,河心洲数量明显减少,河网密度降低。工程水库淹没、尾水渠以及泄洪渠、左侧河道弃渣造地占据了原天然河道中59.5%的河心洲,河网长度减少了34.2%,河网密度降低为1.87 km/km2,天然河流物理结构被完全改变,多样性明显降低,形成了生境条件差异显著的电站库区段、坝下段以及左侧生态河道段。随着栖息环境的变化,鱼类群落组成、多样性等在时空分布上也将呈现出不同程度的差异。因此,研究鱼类群落结构的演替趋势能够直接反映出生态系统对环境变化的响应(Guun et al,2008),也是评价水生态系统健康状况最直接、有效的指示因子(Paller et al,2000)。
3.2 鱼类群落结构空间分布呈现差异性
本次大渡河乐山段鱼类资源调查共采集到54种,10 751尾,总重80 421.5 g,以鲤科鱼类为主,占总种数的50.00%;其次以鳅科与鲿科种类较多,分别占总种数的12.96%。安谷水电站坝下段河面开阔,底质以卵石、砾石为主,常年保持流水态,与大渡河河口直接相连,上下交流通道畅通,采集到的鱼类种类(50种)以及多样性指数(H'=3.2033、J'=0.8188、R=5.7234)均明显高于电站库区段及左侧生态河道段。从鱼类个体生态学矩阵对比分析,电站坝下段鱼类以适宜流水生境种类较多,而电站库区与左侧生态河道段均以适宜静缓流水域的种类较多;同时,3段水域均以杂食性、底栖生活、喜砂砾底质、产粘沉性卵的鱼类为主。可见鱼类群落的组成分布呈现出空间差异性,主要体现在电站坝下段鱼类群落结构稳定与种类丰富,且对水流条件有一定需求,有别于电站库区和左侧生态河道段,与安谷水电站工程布置及调度运行特点密切相关。
3.3 鱼类小型化及渔业资源衰减原因
本次调查到的小型鱼类短体副鳅、蛇鮈、乐山小鰾鮈、宽鳍鱲、棒花鱼、山鳅等在个体数量上占比达90%以上,小型鱼类占据渔获物的优势地位;同时,从渔获物个体的大小上看,体长不足10 cm的鱼类占渔获物总数的77.42%,说明渔业资源正面临着小型化、低质化的压力(陈校辉,2007;刘飞等,2019),反映出大渡河乐山段渔业资源呈现逐渐衰退的趋势。随着大渡河、青衣江水电梯级以及大渡河汇入后的岷江下游航电梯级工程的规划开发,河流所承受的服务功能将越来越多,必将导致鱼类栖息地退化,造成鱼类资源衰竭(Kang et al,2009)。过度捕捞、水利工程建设、河道整治、水生态环境变化、饵料生物匮乏、产卵场损失等因素,都可能对鱼类资源造成一定的影响(江小寒,2019)。
大渡河乐山段鱼类资源小型化的主要原因,一方面是由于小型鱼类的生活史属于r-对策者(Mathieu et al,2008),其种群特点是生命周期短、年龄结构简单、繁殖能力强、饵料来源丰富、栖息环境需求低(卢继武等,1995),对安谷水电站建设后形成的新环境适应快,群落结构更稳定;而大型鱼类则相反,对环境变化更敏感,种群恢复能力较弱。另一方面,长江十年禁渔前,大渡河乐山段野生鱼类经济效益远高于养殖鱼类,某些江鱼的价格可达400元/kg,高额利润驱使下的捕捞强度不断增加,捕捞网目甚至缩小至1~2 cm。Pauly等(1998)指出,在酷渔滥捕的影响下,大个体高营养级捕食者持续减少、衰退,降低了对小型鱼类种群的捕食与竞争压力。十年禁渔前,大渡河乐山段的过度捕捞导致鱼类群落结构转变(毛志刚等,2016),渔获不断向小个体、营养层次低、经济价值不高的种类转变,渔业资源明显衰退。预计随着长江十年禁渔制度的实施,渔民上岸,公众生态保护意识提高,大渡河鱼类资源将得到有效恢复;再者,随着大渡河流域水电梯级工程的开发,河流生态系统结构及功能被改变,鱼类生存环境由河流相向湖泊相转化,能量流、信息流被阻隔,鱼类有效栖息空间萎缩,其群落结构组成、分布及资源量均受到一定影响。
3.4 大渡河鱼类资源保护对策与建议
在大渡河乐山段的调查中,小型鱼类在种类和数量上均占优,在乐山段的能量流动与物质循环中具有十分重要的作用。但小型鱼类在摄食河流中大量饵料资源的同时,也会与许多经济鱼类形成食物和空间的竞争关系(梁彦龄和刘伙泉,1995),其种群资源的不断扩张会造成渔业资源的衰退。 因此,应该合理控制小型鱼类资源,防止其种群过度增长,保护鱼类群落多样性,特提出如下保护建议:
(1)加强对大渡河乐山段鱼类群落组成及多样性变动规律方面的监测与研究,以及时掌握渔业资源结构与发展趋势,准确评估河流生态系统健康水平。
(2)加强人工繁殖研究以及增殖放流效果评估,特别是对胭脂鱼、长薄鳅、泉水鱼、唇?、花?、黄颡鱼、大鳍鳠等大中型经济鱼类以及食肉性鱼类资源增殖需求的研究。
(3)开展科学生态调度措施研究,保证流水环境栖息鱼类的生存需求以及大中型经济鱼类等集中繁殖期3-7月对流水脉冲刺激信号的需求。
(4)严格控制沿河周边工矿企业的污水排放,保持优良的鱼类栖息水质环境。
参考文献
陈校辉,2007. 长江江苏段水生生物调查与研究[D]. 南京:南京农业大学.
程济生,俞连福,2004. 黄、东海冬季底层鱼类群落结构及多样性变化[J]. 水产学报, 28(1):29-34.
丁瑞华,1994. 四川鱼类志[M]. 成都:四川科学技术出版社.
湖北省水生生物研究所鱼类研究室,1976. 长江鱼类[M]. 北京:科学出版社.
江小寒,2019. 航道整治工程对水域生态环境影响探讨[J]. 中国水运, (12):102-103.
蒋志刚,纪力强,1999. 鸟兽物种多样性测度的G-F指数方法[J]. 生物多样性, 7(3):220-225.
黎道丰,蔡庆华,2000. 不同盐碱度水体的鱼类区系结构及主要经济鱼类生长的比较[J]. 水生生物学报, 24(5):493-501.
李博,杨持,林鹏,2000. 生态学[M]. 北京:高等教育出版社.
梁彦龄,刘伙泉,1995. 草型湖泊资源、环境与渔业生态学管理[M]. 北京:科学出版社:205-210.
刘恩生,刘正文,陈伟民,等,2005. 太湖鱼类产量、组成的变动规律及与环境的相互关系[J]. 湖泊科学, 17(3):251-256.
刘恩生,2007. 鱼类与水环境间相互关系的研究回顾和设想[J]. 水产学报, 31(3):391-399.
刘恩生,刘正文,鲍传和,2007. 太湖鲫数量变化的规律及与水环境间关系的分析[J]. 湖泊科学, 19(3):340-345.
刘飞,林鹏程,黎明政,等,2019. 长江流域鱼类资源现状与保护对策[J]. 水生生物学报, 43(S1):144-156.
卢继武,罗秉征,兰永伦,等,1995. 中国近海渔业资源结构特点及演替的研究[J]. 海洋科学集刊, (10):198-210.
毛志刚,谷孝鸿,曾庆飞,2016. 呼伦湖鱼类群落结构及其渔业资源变化[J]. 湖泊科学, 28(2):387-394.
王丹,赵亚辉,张春光,等,2007. 广西野生淡水鱼类的物种多样性及其资源的可持续利用[J]. 动物分类学报, 32(1):160-173.
熊飞,刘红艳,段辛斌,等,2015. 长江上游宜宾江段渔业资源现状研究[J]. 西南大学学报(自然科学版), 37(11):43-50.
张春光,张亚辉,等,2016. 中国内陆鱼类物种与分布[M]. 北京:科学出版社.
张觉民, 1991. 内陆水域渔业自然资源调查手册[M]. 北京:科学出版社.
张雅芝,黄良敏,2009. 厦门东海域鱼类的群落结构及种类多样性研究[J]. 热带海洋学报, 28(2):66-76.
Costa M J, Vasconcelos R, Costa J L, et al, 2007. River flow influence on the fish community of the Tagus estuary (Portugal)[J]. Hydrobiologia, 587:113-123.
Feyrer F, Healey M P, 2003. Fish community structure and environmental correlates in the highly altered southern Sacramento-San Joaquin Delta[J]. Environmental Biology of Fishes, 66:123-132.
Garcia A M, Raseira M B, Vieira J P, et al, 2003. Spatiotemporal variation in shallow-water freshwater fish distribution and abundance in a large subtropical coastal lagoon[J]. Environmental Biology of Fishes, 68:215-228.
Gerhard P, Moraes R, Molander S, 2004. Stream fish communities and their associations to habitat variables in a rain forest reserve in southeastern Brazil[J].Environmental Biology of Fishes, 71:321-340.
Gunn J S, Clear N P, Carter T I, et al, 2008. Age and growth in southern bluefin tuna, Thunnus maccoyii (Castelnau): Direct estimation from otoliths,scales and vertebrae[J]. Fisheries Research, 92:207-220.
Kang B, He D M, Perrett L, et al, 2009. Fish and fisheries in the Upper Mekong: current assessment of the fish community, threats and conservation[J]. Reviews in Fish Biology and Fisheries, 19(4):465-480.
Koel T M, Peterka J J, 2003. Stream fish communities and environmental correlates in the Red River of the North, Minnesota and North Dakota[J]. Environmental Biology of Fiehes, 67:137-155.
Leitao R, Martinho F, Cabral H N, et al, 2007.The fish assemblage of the Mondego estuary:composition, structure and trends over the past two decades[J]. Hydrobiologia, 587:269-279.
Maes J, Damme S V, Meire P, et al, 2004. Statistical modeling of seasonal and environmental influences on the population dynamics of an estuarine fish community[J]. Marine Biology, 145:1033-1042.
Margalef D R, 1957. Information theory in ecology[J]. International Journal of General Systems, (3):36-71.
Mathieu B, Clement C, Eric M, et al, 2008. Assessing habitat selection using multivariate statistics: Some refinements of the ecological-niche factor analysis[J]. Ecological Modelling, 211:233-240.
Paller M H, Reichert M J M, Dean J M, et al, 2000. Use of fish community data to evaluate restoration success of a riparian stream[J]. Ecological Engineering, 15:431-446.
Pauly D, Christensen V, Dalsgaard J, et al, 1998. Fishing down marine food webs[J]. Science, 279(6):860-863.
Pianka E R, 1971. Ecology of the Agamid Lizard Amphibolurus isolepis in Western Australia[J]. Copeia, 527-536.
Pielou E C, 1975. Ecological diversity[M]. New York: John Wiley:46-49.
Sutela T, Vehanen T, 2008. Effects of water-level regulation on the nearshore fish community in boreal lakes[J]. Hydrobiologia, 613:13-20.
Wilhm J L, 1968. Use of biomass units in Shannon's formula[J]. Ecology, 49:153-156.
(责任编辑 万月华)
Fish Community Structure and Diversity in the Leshan Section of Dadu River
WANG Wen‐jun1, FANG Yan‐hong1, YANG Zhong1, HUANG Jin2, HE Chun‐hua3, DU Xia3
(1. Key Laboratory of Ecological Impacts of Hydraulic-Projects and Restoration of Aquatic Ecosystem
of Ministry of Water Resources, Institute of Hydroecology,
Ministry of Water Resources and Chinese Academy of Sciences, Wuhan 430079, P.R. China;
2. China Hydropower Construction Group Shengda hydropower Company, Leshan 614013, P.R. China;
3.Yalong River Hydropower Development Company, LTD. Chengdu 610051, P.R. China)
Abstract:In this study, we investigated fish community structure and diversity in the Leshan section of Dadu River after the construction of Angu hydropower station. The results were then used to analyze the response of the fish community to the construction of the hydropower station. The aim was to provide a basic reference for the protection and restoration of the river ecosystem. From April to May and September to October of 2016-2018, a fishery resource investigation was carried out across 7 transects (S1, S2 above the dam, S3, S4 below the dam, S5, S6, S7 in the left ecological river) on four occasions, along 30 km of the Leshan section. Fish samples were collected using multi-mesh fixed gill nets and fixed cages. Biological measurements of the total length, body length, body weight and species of all specimens collected were determined in the field. A total of 10 751 specimens from 54 species, 45 genera, 12 families and 5 orders were collected, with absolute dominance of Cyprinids (27 species), which accounted for 50.00% of the total fish species. The number of fish species collected in Angu reservoir, below the dam and in the left ecological river was 36, 50 and 35, with corresponding dominant species number of 6, 7 and 7. Ecological types of fish in the section investigated consisted primarily of omnivorous groups, preferring a gravel substrate and laying adhesive eggs. In general, the proportion of species preferring lotic habitats below Angu dam were higher than in Angu Reservoir and the ecological river. The Shannon-Wiener diversity (H'), Pielou evenness (J') and Margalef richness (R) indices in the river section below Angu dam were highest, with respective values of 3.2033, 0.8188 and 5.7234. The proportion of individuals in the body length range of 4-10cm for the reservoir, river section below the dam and ecological river were, respectively 69.17%, 76.11% and 78.36% of the total catches. A tendency towards miniaturization and lower quality fish was evident in the river section investigated, indicating a decline of the fish resource. Overfishing and hydroelectric development are the primary factors threatening fish resources in the Leshan section of Dadu River. Based on the results, we recommend measures to protect fish diversity.
Key words:fish community; distribution characteristics; diversity index; Angu hydropower station
收稿日期:2021-08-23 修回日期:2022-11-23
基金项目:水利部重大科技项目(SKR-2022015);水利部公益性行业科研专项(200901009);中国水利水电建设集团公司科研项目(AG2012/S-46-A)。
作者简介:王文君,1980年生,女,高级工程师,主要从事水生态保护与修复工作。E-mail:26759040@qq.com
猜你喜欢
安徽农学通报(2017年1期)2017-02-15 18:21:58
中国医药导报(2016年28期)2017-01-06 19:52:54
农业与技术(2016年20期)2016-12-08 21:37:07
戏剧之家(2016年22期)2016-11-30 15:13:39
散文百家·下旬刊(2016年9期)2016-11-23 22:23:14
散文百家·下旬刊(2016年10期)2016-11-17 21:22:22
科技资讯(2016年19期)2016-11-15 10:39:12
人间(2016年27期)2016-11-11 16:27:23
农业与技术(2016年15期)2016-11-09 07:11:20
赤峰学院学报·哲学社会科学版(2015年11期)2015-12-10 14:13:58
