
云南咖啡锈菌生理小种鉴定
2023-04-27 03:35:56白学慧萧自位马关润陈振佳李锦红
热带农业科技 2023年2期
白学慧 ,萧自位 ,马关润 ,罗 琴 ,陈振佳 ,李锦红 *
(1.云南省德宏热带农业科学研究所,云南瑞丽 678600;2.中国咖啡工程研究中心,云南芒市 678400)
咖啡是世界上重要的饮料作物和热带经济作物,云南热区咖啡适种区面积约40 万hm2,截至2021 年底,咖啡种植面积9.29 万hm2、农业产值26.04 亿元,综合产值339.25 亿元。云南省咖啡面积、产量和农业产值均占全国的98%以上。咖啡产业是云南省打造世界一流“绿色食品牌”的高原特色农业产业之一。 由咖啡驼孢锈菌(Hemileia vastatrixBerk. & Br.)引起的咖啡锈病是咖啡生产中最重要的病害,在世界各主要咖啡产区均有不同程度发生,并以流行猛烈、传播迅速、损失惨重而著称[1],在适宜的气候条件下,咖啡锈病可以导致叶片损失达50%,浆果损失达70%[2]。咖啡锈病在中美洲及哥伦比亚、秘鲁和厄瓜多尔发生严重并广泛流行,造成了很大危害[3-5]。咖啡锈病对云南小粒种咖啡的产量和品质影响很大,是限制咖啡产业可持续发展的重要因素[6],种植咖啡抗病品种是控制该病害最为经济、有效和安全的措施[7]。
弄清病原菌生理小种分化情况是开展抗锈育种的前提和基础。在咖啡种植国当中,印度最先对咖啡锈病进行研究,印度学者Mayne 首次报道印度存在咖啡锈菌生理分化现象,鉴定出2 个小种[8],并于1939 年鉴定出另外两个生理小种(race3 和race4),其研究成果为后续研究奠定了基础[9]。国际咖啡锈病研究中心(CIFC)经过50多年的研究,对全世界46 个咖啡种植国的咖啡锈菌样品进行小种鉴定,已发现小种49 个,分别用罗马数字 I,II,III 等表示[10]。我国陈振佳等 1998年报道云南咖啡种植区有 I(v2,v5),II(v5),XV(v4,v5)和 XXIV(v2,v4,v5)4 个生理小种[11]。白学慧等2018 年报道云南咖啡植区有VIII(v2,v3,v5)、XXXIII(v5,v7 或 v5,v7,v9)、XXXIV(v2,v5,7 或 v2,v5,v7,v9)、XXXVII(v2,v5,v6,v7,v9)、XLI(v2,v5,v8)、XLII(v2,v5,v7,v8 或 v2,v5,v7,v8,v9),New race(v2,v5,v6,v7),New race(v1,v2,v5,v7 或 v1,v2,v5,v7,v9),New race(v1,v5,v7 或 v1,v5,v7,v9)9 个生理小种[12]。
Bettencourt 等[13]1985 年报道,咖啡种群有抗锈病基因9 个,用SH1~SH9 表示,锈菌小种的毒力基因也有9 个,用v1~v9 表示,且小种的毒性基因与咖啡种群的抗病基因对应。Van der Vossen 等[14]2005 年报道由于世界不同咖啡种植区新的锈菌生理小种不断出现,咖啡抗锈基因SH1~SH9 已经被陆续攻克。XXXIII(v5,v7 或 v5,v7,v9)、XXXIV(v2,5,7 或 v2,v5,v7,v9)、XXXVII(v2,v5,v6,v7,v9)、XXXIX(v2,v4,v5,v6,v7,v8,v9)等生理小种的出现,使抗病品种卡蒂姆7963 先后在印度、巴西、肯尼亚、坦桑尼亚、泰国、印度尼西亚、菲律宾、哥伦比亚和中国等咖啡种植区逐渐丧失抗锈性[15-20],由于咖啡锈菌群体遗传变异,培育具咖啡锈病持久抗病性的抗病品种一直是咖啡抗性育种的挑战。掌握咖啡锈菌生理小种分化及地理分布对该病的有效防治具有重要意义。本研究利用19 个国际通用的咖啡锈菌鉴别寄主,采用人工活体接种的方式,对2017—2019 年从云南咖啡植区15 个县(市)采集的锈菌样本进行生理小种鉴定,以期为云南咖啡品种合理布局及咖啡育种策略的调整提供重要依据。
1 材料和方法
1.1 材料
1.1.1 菌株来源
菌株样本主要为2017/2018 年度、2018/2019 年度、2019/2020 年度从云南15 个县(市)采集的锈菌孢子,采集地咖啡栽培品种及生态环境较具代表性。孢子经纯化、扩繁、保存了58 份用于生理小种鉴定,其中 2017/2018 年度 24 份、2018/2019 年度25 份、2019/2020 年度9 份。样本采集信息详见表1。
1.1.2 鉴别寄主
采用19 个通用的鉴别寄主,包括15 个小粒咖啡(Coffea arabica)或四倍体杂交咖啡(Tetrapliod interspecific hybrids),4 个 二 倍 体 咖 啡(Coffeaspp.),品种所含抗病基因详见表2。
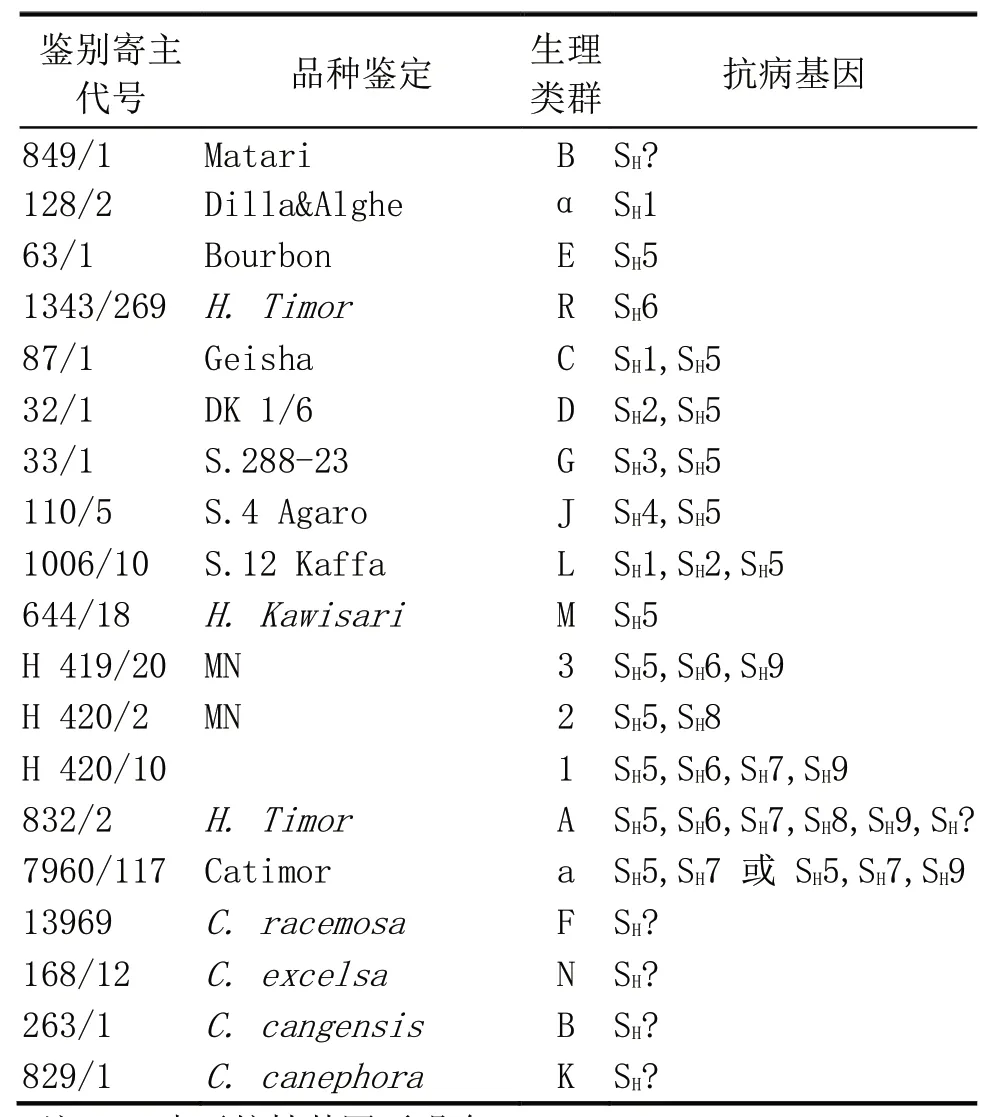
表2 咖啡锈菌小种鉴定所用鉴别寄主及其携带抗病基因
1.2 鉴定方法
鉴定在云南省德宏热带农业科学研究所温室进行,菌株的保存、扩繁和鉴定方法按照中华人民共和国农业行业标准(NY/T3331—2018)《热带作物品种资源抗病虫鉴定技术规程咖啡锈病》中描述的方法进行[21]。将采集的锈菌样本用解剖刀轻轻刮下单个夏孢子堆,接种在鉴别品种Matari(849/1)幼嫩的新叶背面,用毛刷轻轻涂抹均匀,并轻轻喷施水雾,至有水膜产生为止,然后套塑料袋保湿遮光48 h。约30 d 后收集夏孢子并接种至不同鉴别品种,接种后30~45 d 开始调查,间隔1 周调查2 次,每个锈菌样本重复3 次。
1.3 抗病性分级标准
参照中华人民共和国农业行业标准(NY/T3331-2018)《热带作物品种资源抗病虫鉴定技术规程 咖啡锈病》的分级标准[21],按10 级划分(表 3,图 1)。其中 0~3 级为抗病(R)、4~5 级为中抗(MR)、6~7 级为中感(MS)、8~9 级为感病(S)。

图1 咖啡锈病不同病级症状图

表3 咖啡锈病病情分级标准
2 结果与分析
2.1 锈菌生理小种类型及地理分布
根据咖啡锈菌样本与鉴别接种的抗感反应,鉴定出12 个生理小种(表4),分别为I(v2,v5)、II(v5)、VIII(v2,v3,v5)、XVII(v1,v2,v5)、XXIV(v2,v4,v5)、XXXIII(v5,v7 或 v5,v7,v9)、XXXIV(v2,v5,v7 或 v2,v5,v7,v9)、XXXV(v2,v4,v5,v7,v9)、XXXVIII(v2,v5,v6,v7,v9)、XXXIX(v2,v4,v5,v6,v7,v8,v9)、XLI(v2,v5,v8)和 XLII(v2,v5,v7,v8 或 v2,v5,v7,v8,v9)。其中:云南德宏植区有 7 个,分别为 I、VIII、XVII、XXIV、XXXIV、XXXV 和 XLII 型 小 种;大 理 1 个,仅 有 II 型小 种 ;普洱 6 个,分别为 I、II、XXXIV、XXXVII、XXXIX 及XLII 型小种;保山 5 个,分别为 I、II、XXXIII、XXXIV 及 XLII 型 小种 ;临 沧 5 个 ,分 别为 II、XXXIII、XXXIV、XLI 及 XLII 型小种(表1,表5)。
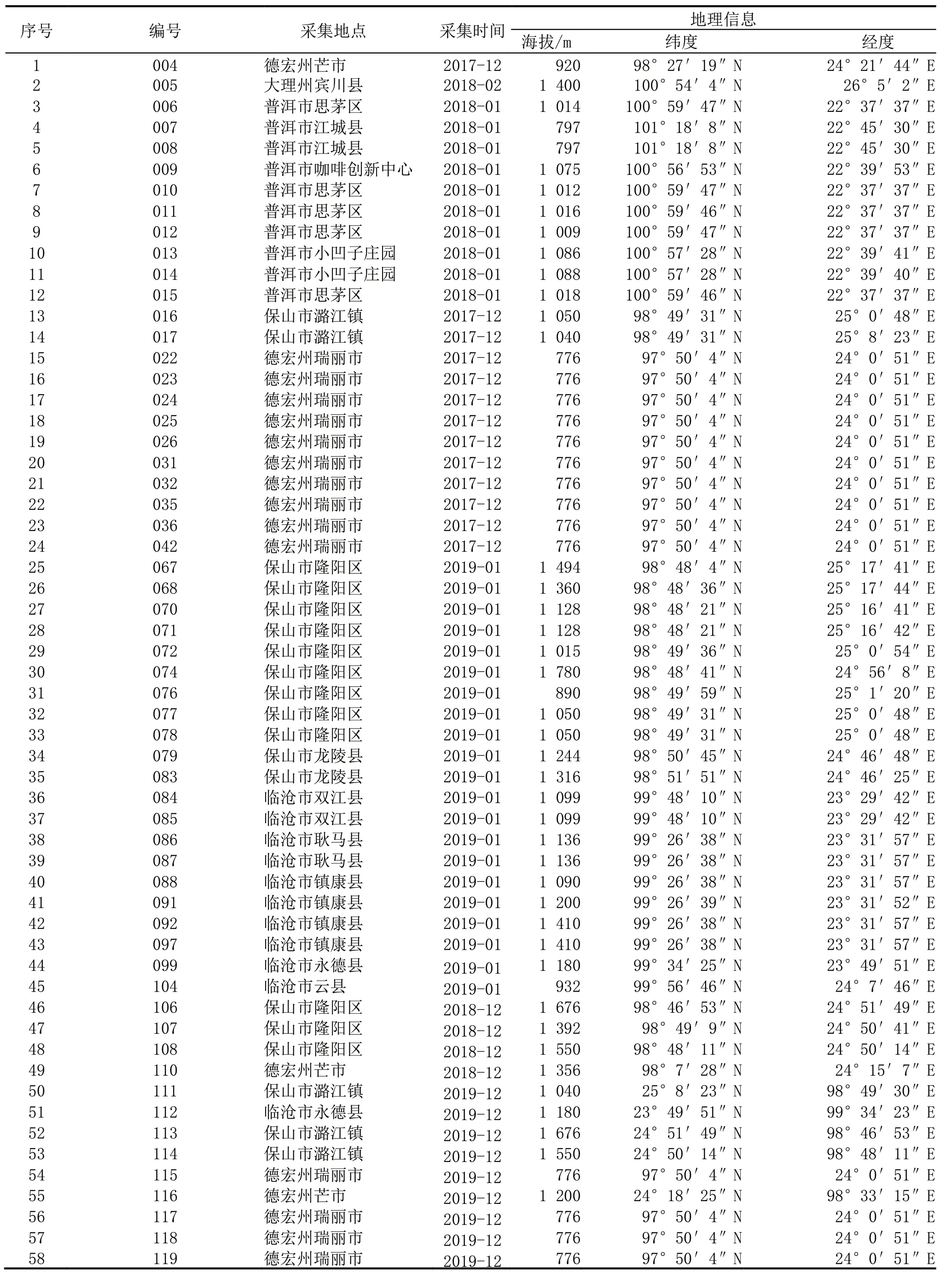
表1 咖啡锈菌样本采集点及寄主

表4 不同锈菌小种在鉴别寄主上的抗病类型
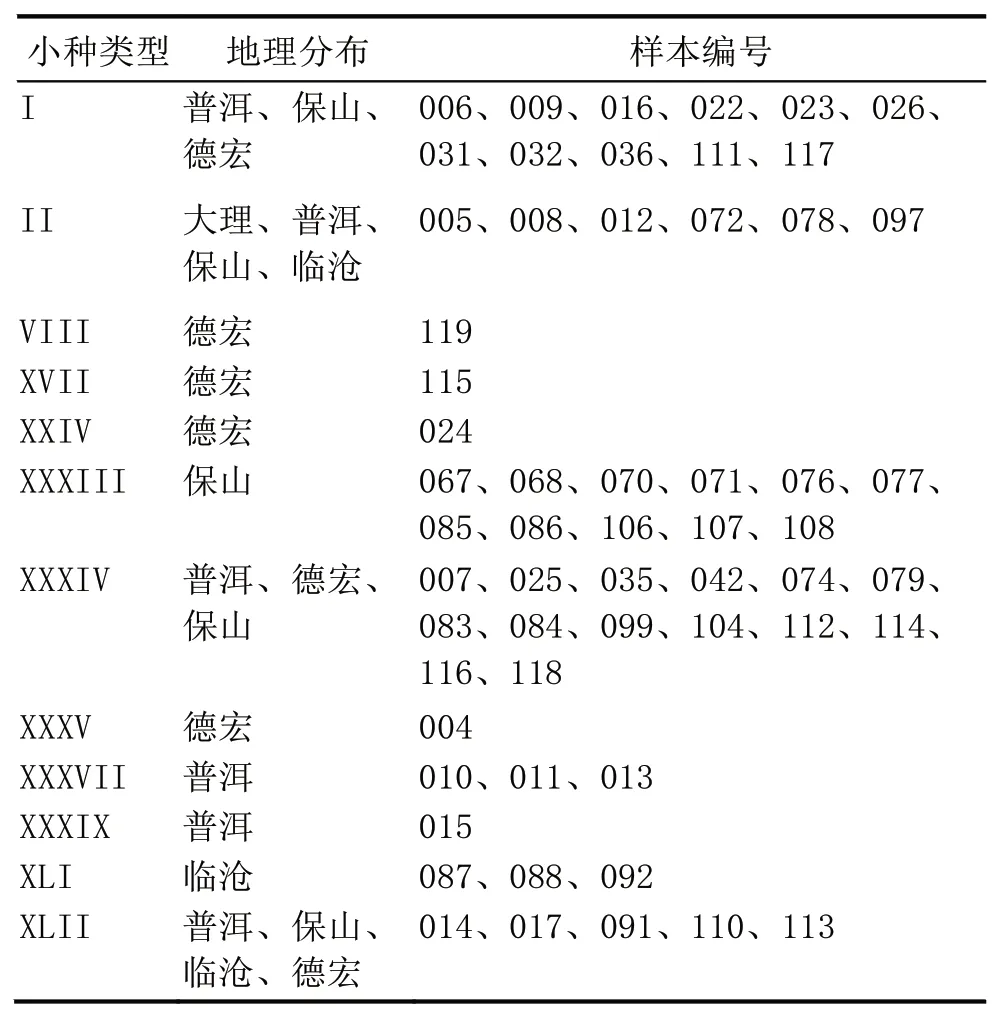
表5 锈菌小种类型及地理分布
2.2 锈菌生理小种出现频率及毒力基因种类
本次研究中对58 份锈菌样本进行鉴定,共鉴定出12 个生理小种。其中,XXXIV 型生理小种为优势小种,占样本的24.14%;I 型和XXXIII 型小种均占样本的18.97%;II 占样本的10.34%;XLII 占样本的8.62%;XXXVII 和XLI 均占样本的5.18%;VIII、XVII、XXIV、XXXV 和 XXXIX 分别占样本的1.72%(表6)。本次鉴定结果表明,云南咖啡主产区包含v1~v9 共9 个毒力基因。其中,含单个毒力基因(II 号生理小种)仅占样本总数的10.34%,其余均为双至多个毒力基因(表6)。
从地区看,德宏地区共17 个样本,I 型生理小种7 个,占德宏地区41.18%,保山地区19 个样本,XXXIII 型生理小种 9 个,占保山地区47.37%。普洱和临沧差距不大。与2018 年白学慧等的报道相比,新鉴定出6 个生理小种,分别为 I(v2,v5)、II(v5)、XVII(v1,v2,v5)、XXIV(v2,v4,v5)、XXXV(v2,v4,v5,v7,v9)和 XXXIX(v2,v4,v5,v6,v7,v8,v9)[13]。表明云南咖啡主产区锈菌小种类型呈现增加的趋势,对抗锈育种提出了新的挑战。
3 讨论
咖啡锈病(H. vastatrixBerkeley & Broome)对咖啡野生种和栽培种均有危害,对小粒种咖啡(Coffea arabica)的危害最为严重。1861年该病害被英国探险队首次在东非维多利亚湖畔发现,1868 年在斯里兰卡首次流行,目前已在全世界咖啡产区流行。种植抗锈品种是控制咖啡锈病最为经济有效的措施。1875 年起,咖啡研究者在田间寻找抗锈病植株,1927 年在Timor岛发现 HDT(C. canphora和C. arabica天然杂交),上世纪50 年代CIFC(葡萄牙咖啡锈病研究中心)以HDT 作为主要抗源与世界各主产国联合开展了抗锈咖啡品种的选育[22]。但是,在选育具有抗病性品种的同时,随之出现的锈菌新型小种逐渐克服了抗病咖啡品种的抗性,使其抗性仅在一定时间及空间内有效。如,‘S. 288’主要对II 型小种有抗性,引入云南种植14 年后抗性逐渐丧失。‘卡蒂姆CIFC7963’引入云南从推广到失去抗性仅 9 年时间[23]。Várzea 等[24]2005 年报道咖啡锈菌新小种产生的机制可能源自锈菌变异,Robinson[25]、Vanderplank[26]、Eskes[27]1983 年报道病原菌小种的形成、分布与该地区栽培品种的抗病基因相关,新小种是以克服品种抗病性为目的而形成的。本研究鉴定的 I(v2,v5)、II(v5)和 XXIV(v2,v4,v5)生理小种寄主均为铁皮卡(Typica)、波邦(Bourbon)、卡杜拉(Caturra)等感病品种(无抗源介入),XXXIII(v5,v7 或 v5,v7,v9)、XXXIV(v2,v5,7 或 v2,v5,v7,v9)、XXXV(v2,v4,v5,v7,v9)、XXXVII(v2,v5,v6,v7,v9)、XXXIX(v2,v4,v5,v6,v7,v8,v9)、XLI(v2,v5,v8)和 XLII(v2,v5,v7,v8 或 v2,v5,v7,v8,v9)等生理小种寄主均为卡蒂姆系列(Catimor)品种(有HDT 抗源介入),表明咖啡锈菌生理小种的形成、分布与栽培品种的抗病基因相关。
本次研究共鉴定出12 个生理小种,和陈振佳等[11]1998 年、白学慧等[12]2018 年的报道相比,云南咖啡植区新增4 个生理小种,分别为XVII、XXIV、XXXV 和XXXIX。本研究中锈菌包含的毒力基因为 v1~v9,与陈振佳等[11]1998 年的报道相比,增加毒力基因 v1、v3、v6、v8;与白学慧等[12]2018年的报道相比,增加毒力基因v4。综上,云南咖啡锈菌生理小种的类型处于动态变化中,锈菌的种类和毒力基因也在不断增加。这与Van der Vossen 等[14]2005 年报道世界不同咖啡种植区新型锈菌生理小种不断出现,咖啡抗锈基因SH1~SH9 已经被陆续攻克的研究也吻合。
我国目前选育和推广的咖啡抗锈品种的抗源基本都来自HDT。锈病抗源单一,亟待创新、引进新的锈病抗源,并在选育推广咖啡抗锈品种的过程中拓宽手段和策略。
猜你喜欢
山东畜牧兽医(2022年2期)2023-01-05 22:43:29
南方农业学报(2022年11期)2022-03-18 05:55:26
农业技术与装备(2021年12期)2021-12-02 17:24:28
现代园艺(2019年13期)2019-01-06 10:27:12
林业科学(2017年7期)2017-08-30 14:24:16
植物保护(2017年4期)2017-08-09 01:18:38
中国马铃薯(2017年1期)2017-03-02 09:15:51
作物学报(2016年10期)2016-10-19 04:14:09
陕西林业科技(2016年1期)2016-08-04 00:35:49
现代农业(2016年5期)2016-02-28 18:42:36
