
小麦籽粒性状的遗传效应分析及其育种策略
2023-04-25 09:10赵永涛张中州甄士聪望俊森张立超张文斐
麦类作物学报 2023年4期
袁 谦,赵永涛,张中州,甄士聪,望俊森,张立超,张文斐
(1.漯河市农业科学院,河南漯河 462300; 2.中国农业科学院作物科学研究所,北京 100081)
小麦籽粒大小是千粒重的决定因素之一,直接影响小麦产量[1];同时小麦籽粒的形态与品质有较强的相关性[2],是小麦育种的重要目标性状。在小麦育种过程中,对籽粒形态的评价主要通过目测法和人工测量法,但采用目测法评价误差较大,不易量化;采用游标卡尺人工测量的方法效率低,不能批量处理,耗时费力。随着成像技术和图像分析处理技术的发展,借助图像分析处理软件批量且准确地测量籽粒形态成为可能[3-5]。目前,基于OpenCV、Matlab GUI等开发的图像采集分析系统[6-7]以及Image J软件[8]等已成功应用于多种作物的表型研究,但操作复杂、对程序算法也要有一定的基础,难以在育种科研人员中广泛推广;谷物籽粒分析仪[9]能够方便快捷地采集多个表型信息,但价格昂贵,对于基层育种单位负担较重。Tanabata等[10]开发的SmartGrain软件具有操作简单、成本低廉等优点,同时可实现高通量分析处理,是较理想的表型分析工具,目前已在小麦、水稻等作物籽粒研究中得到应用[11-12]。
小麦籽粒形态包括籽粒的长、宽、厚、长宽比、周长、面积以及圆度等因素,是重要的农艺性状[13],解析其遗传机制对小麦高产育种具有重要意义。主基因+多基因混合遗传分析方法可以鉴别主效基因和微效基因,同时明确基因的效应及基因间的效应[14],是一种广泛应用的数量性状遗传分析方法,已经用于小麦抗病性[15-17]、品质[18]、产量性状[19-21]及其他重要农艺性状[22-23]的遗传分析研究中。漯麦76是漯河市农业科学院利用人工合成小麦材料选育的小麦新品种,丰产性突出,具有大粒、大穗、多穗等优异特性,在河南省小麦统一试验中表现优异,品种比较试验、区域试验和生产试验均为产量第一,三年完成试验程序,并于2022年通过河南省初审。本研究以漯麦76为研究材料构建6世代联合群体,通过SmartGrain软件进行图像分析获得籽粒性状数据,利用主基因+多基因混合遗传分析方法明确大粒性状的遗传效应,以期为进一步利用小麦创新种质提供 指导。
1 材料与方法
1.1 试验材料
漯麦76和L529是由漯河市农业科学院分别利用人工合成小麦和偃麦草创制的小麦新种质。漯麦76具有千粒重高、籽粒商品性好、成穗率高等优异特性;L529具有赤霉病抗性好、多穗等优异特性,但籽粒性状一般。2019年以L529(P1)为母本、漯麦76(P2)为父本杂交获得F1群体;2020年F1自交获得F2群体,F1分别利用P1和P2回交获得B1和B2群体。
1.2 试验方法
2020年秋,在漯河市农业科学院试验田种植4个世代6个联合群体(P1、P2、F1、F2、B1和B2)。行长2.5 m,行距0.25 m,每行点播30粒种子,P1、P2和F1各种植3行,F2、B1和B2各种植24行,整个生育期常规田间管理。于小麦蜡熟期收获主茎穗,人工脱粒,保证净度,晾干后备用。分别获得P1、P2和F1群体材料31、32和40份,F2、B1和B2群体材料168、255和174份。
用SmartGrain软件测量获得小麦籽粒表型数据[24]。将种子均匀分散平铺于扫描仪面板,背景设置为黑色,扫描图片的分辨率设置为200 dpi,保存格式为TIFF,将所有图片保存于同一文件夹,图片不需要经过任何处理,直接用SmartGrain软件处理获得籽粒面积、粒长、粒宽、长宽比、周长和圆度以及穗粒数等表型数据。每份试验材料种子用电子天平称量获得单穗重,并结合穗粒数计算千粒重(1 000×单穗重/穗粒数)。
1.3 数据分析
利用R软件对6个群体的7个籽粒性状数据进行描述性统计分析,并采用Shapiro-Wilk检验方法计算W值和P值,进行正态性检验。
2 结果与分析
2.1 描述性统计分析结果
对P1、P2和F1群体的7个籽粒性状进行差异分析,由表1可知,亲本L529(P1)和漯麦76(P2)间籽粒的千粒重、面积、周长、粒长、粒宽5个性状差异均达到极显著水平,但长宽比和圆度差异不显著,说明两个亲本籽粒形状相似,但大小差异较大。L529和漯麦76的千粒重分别为37.44和52.44 g,籽粒面积分别为19.69和55.51 mm2,籽粒周长分别为18.31和19.58 mm,粒长分别为6.97和 7.45 mm,粒宽分别为3.75和 4.01 mm,F1的这5个籽粒性状均介于两个亲本之间,未表现出超亲分离现象,但均更接近大粒亲本漯麦76。
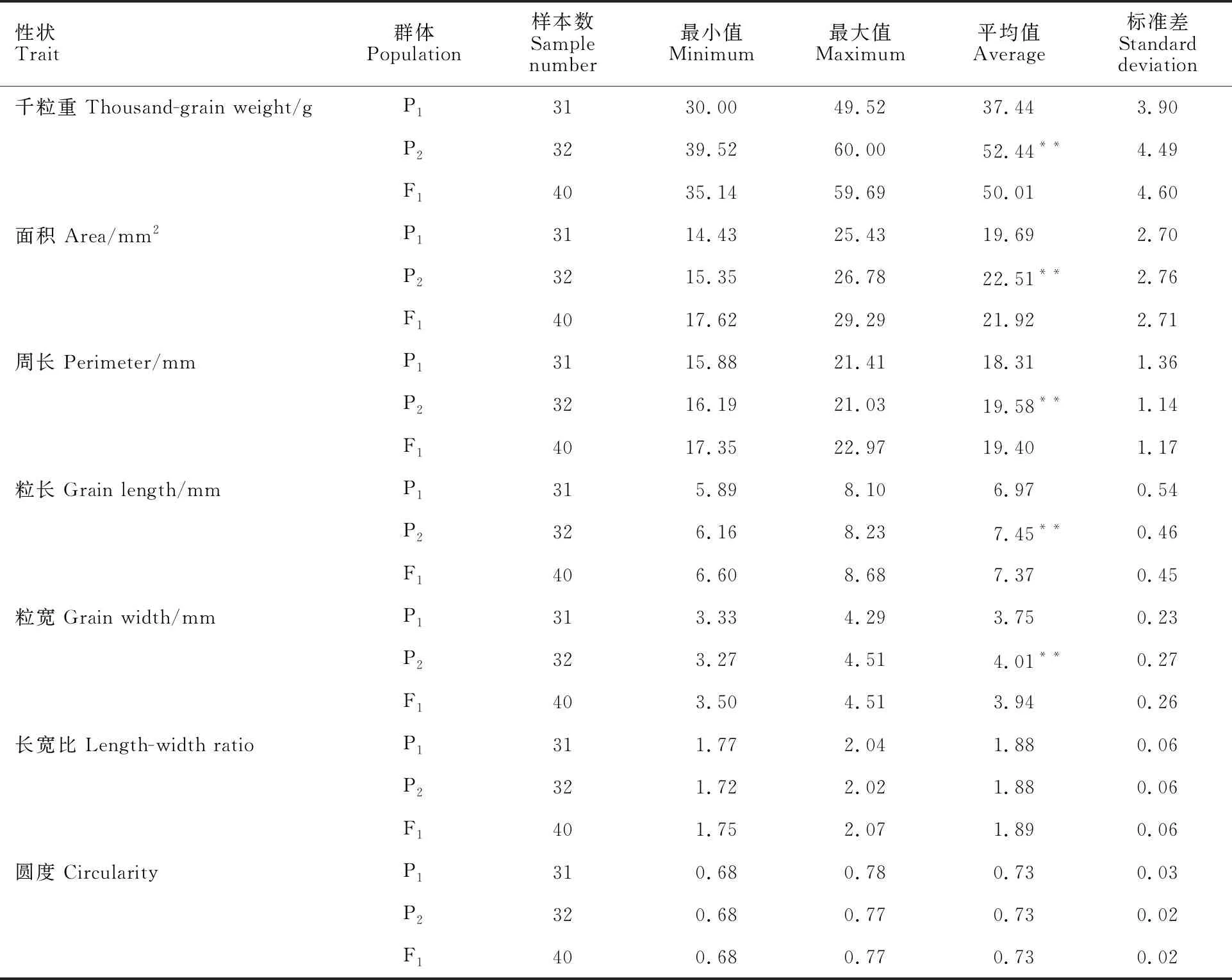
表1 不分离世代群体籽粒性状的描述性统计
对分离世代群体(F2、B1和B2)的千粒重、籽粒面积、籽粒周长、粒长、粒宽5个籽粒性状进行分析,由表2可知,分离世代群体中千粒重最大值与最小值差值介于43.91~54.19 g之间,变异系数介于15.04%~20.45%之间;籽粒面积最大值与最小值差值介于16.16~20.47 mm2之间,变异系数介于16.50%~17.53%之间;籽粒周长最大值与最小值差值介于8.27~9.70 mm之间,变异系数介于8.44%~9.14%之间;粒长最大值与最小值差值介于3.18~3.56 mm之间,变异系数介于8.59%~8.88%之间;粒宽最大值与最小值差值介于1.48~2.20 mm之间,变异系数介于 8.03%~9.90%之间。说明分离世代群体籽粒性状表型变异丰富,明显高于P2、P2和F1群体,且有超亲现象。分离世代偏度和峰度绝对值均小于1,初步判断服从正态分布。Shapiro-Wilk检验结果表明,P值均小于W值,所以分离世代数据均符合正态分布,适合进行遗传分析。
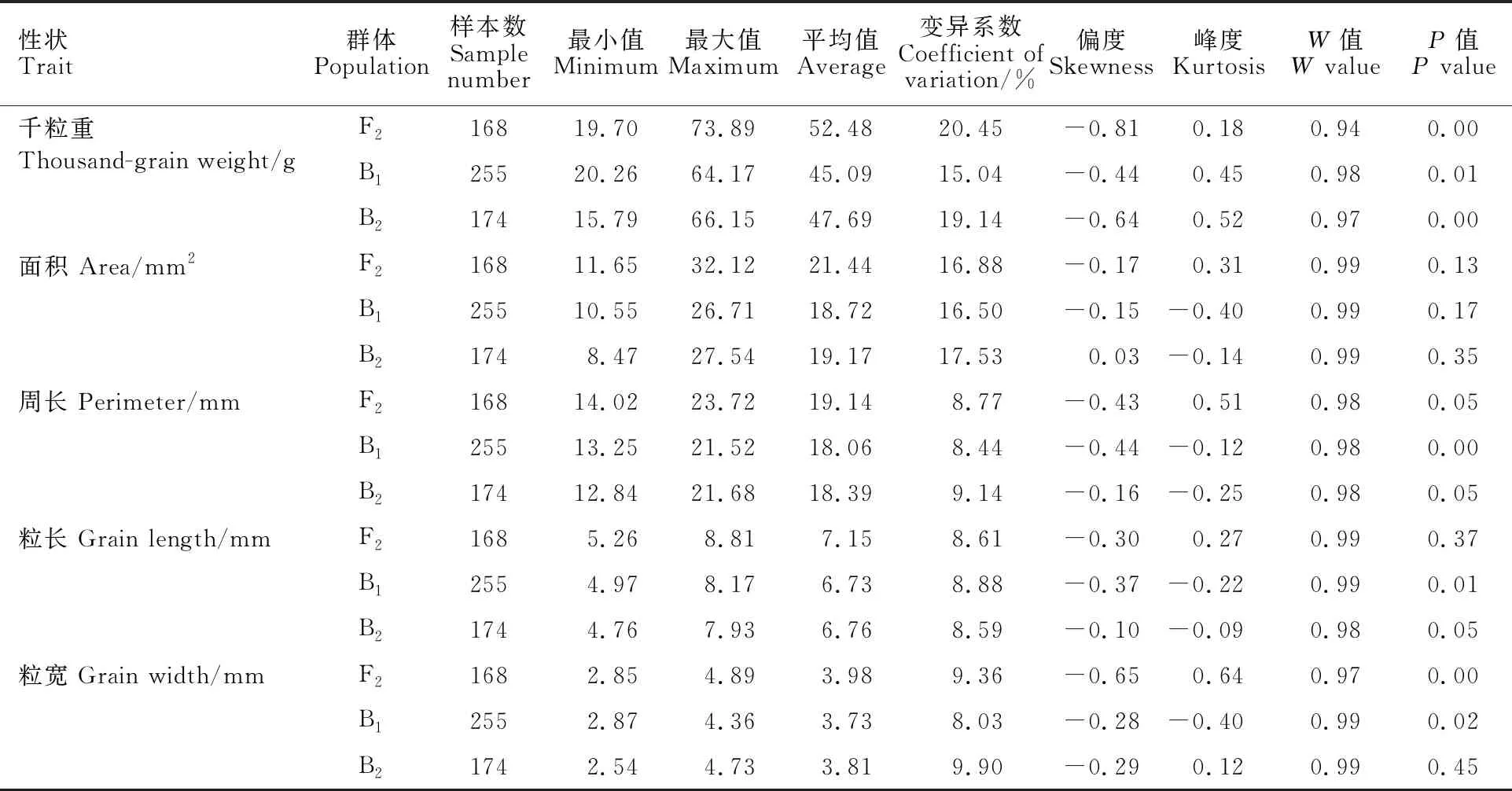
表2 分离世代群体籽粒性状的描述性统计
2.2 最优遗传模型的确定
对6个群体籽粒的千粒重、面积、周长、粒长和粒宽分别进行主基因+多基因混合遗传模型分析,得到1对主基因、2对主基因、多基因、1对主基因+多基因、2对主基因+多基因共5类24种遗传模型。根据最优模型的选取原则,选取AIC值最小或接近最小的3个模型作为备选模型(表3)。其中,千粒重的MX2-ADI-AD、2MG-ADI和MX2-ADI-ADI模型AIC值较小,分别为 4 837.53、4 842.01和4 844.49;籽粒面积的PG-ADI、MX1-AD-ADI和2MG-ADI模型AIC值较小,分别为3 636.19、3 640.19和3 644.31;籽粒周长的2MG-ADI、MX2-ADI-ADI和MX2-ADI-AD模型AIC值较小,分别为2 603.88、2 607.84和2 608.70;粒长的PG-ADI、2MG-ADI和MX2-ADI-ADI模型AIC值较小,分别为1 234.28、 1 234.35和1 235.55;粒宽的2MG-ADI、MX2-ADI-AD和PG-ADI模型AIC值较小,分别为429.47、429.60和432.76。
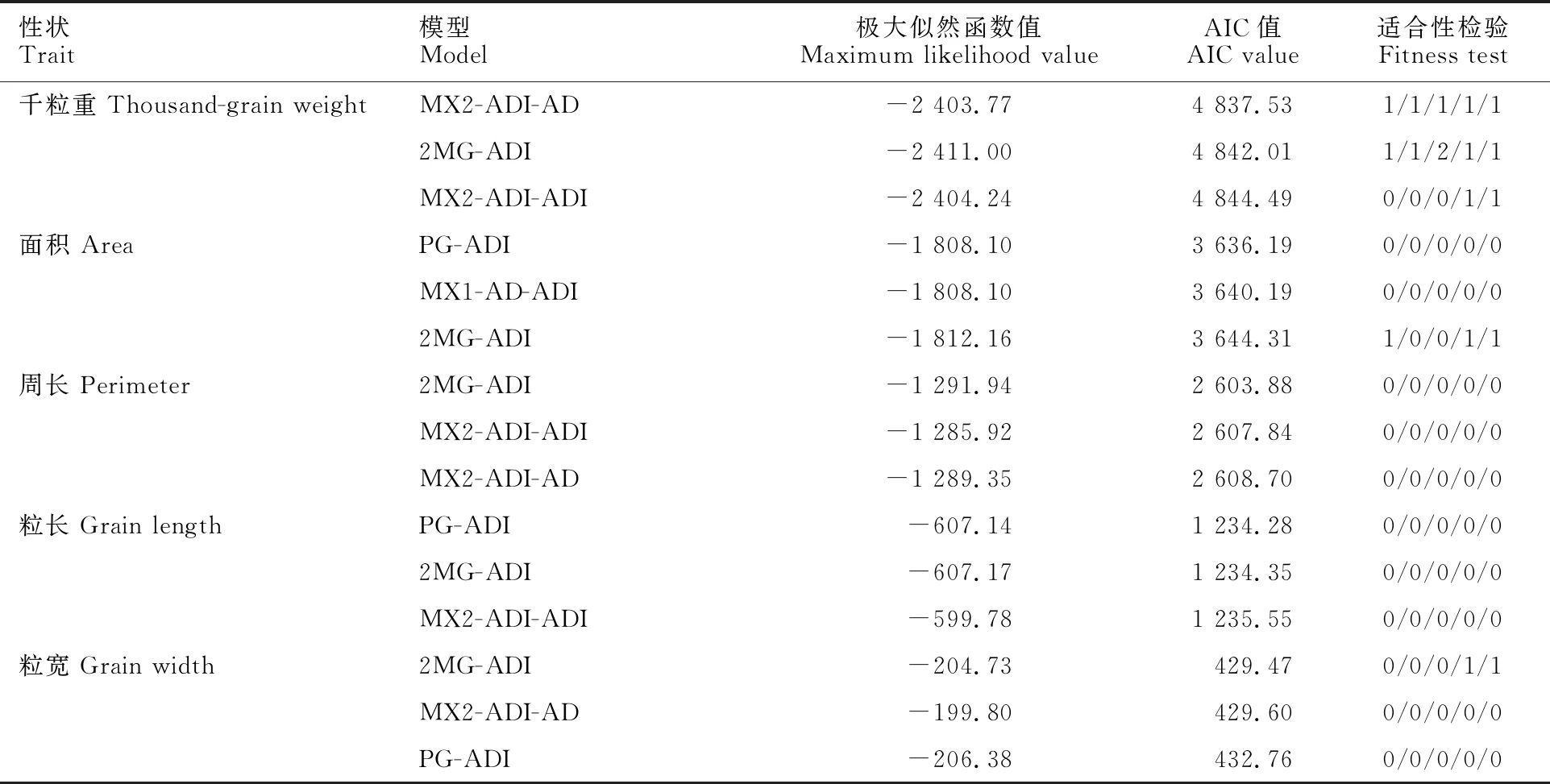
表3 籽粒性状备选模型的极大似然函数值和AIC值及其适合性检验
2.3 遗传参数估计
利用R软件包SEA v2.0根据最小二乘法原理对千粒重、籽粒面积、籽粒周长、粒长、粒宽5个籽粒性状的最优模型进行一阶遗传参数和二阶遗传参数估计(表4)。千粒重的最佳遗传模型为2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型(MX2-ADI-ADI)。由一阶遗传参数可知,控制千粒重的2对主基因为负向的加性效应,效应值均为-5.19;显性效应为正向效应,效应值分别为1.28和1.27,2对主基因热能比值(ha/da、hb/db)绝对值均小于1,说明2对主基因均以加性效应为主。2对主基因的加性×加性上位性效应和显性×显性上位性效应均为负值,且绝对值远大于加性×显性上位性效应和显性×加性上位性效应,进一步说明2对主基因的遗传效应以负向加性效应为主,说明小粒亲本对后代千粒重的负向影响较大,配制组合易采用“高粒重×高粒重”组合配制方案。由二阶遗传参数可知,3个分离世代(F2、B1和B2)千粒重的主基因+多基因遗传率分别为83.80%、59.44%和77.61%,说明千粒重主要受遗传因素控制;主基因遗传率分别为 66.88%、39.58%和59.38%,均远大于多基因遗传率,说明在千粒重的遗传贡献中以主基因为主。

表4 籽粒各性状最佳遗传模型的遗传参数
籽粒面积的最佳遗传模型为加性-显性-上位性多基因遗传模型(PG-ADI)。由二阶遗传参数可知,3个分离世代(F2、B1和B2)籽粒面积的多基因遗传率分别为44.81%、24.37%和35.95%,说明籽粒面积受环境因素影响较大,遗传因素起到一定作用,在F2和B2分离世代中遗传因素影响稍大。因此,宜采用多环境鉴定的方法进行选择。
籽粒周长的最佳遗传模型为2对加性-显性-上位性主基因遗传模型(2MG-ADI)。由一阶遗传参数可知,控制籽粒周长的2对主基因为负向的加性效应,效应值均为-0.35,显性效应也均为负向效应,效应值分别为-2.15和-2.68,2对主基因的势能比值(ha/da、hb/db)绝对值分别为 6.14和7.66,远大于1,说明2对主基因以负向的显性效应为主;上位性效应以显性×显性上位性效应为主,效应值为4.25,大于其他上位性效应,且大于其他上位性效应绝对值之和,该上位性效应为正向效应,说明2对主基因可能为隐性基因,稳定纯合后会获得较好的正向效应,宜在高代进行选择。由二阶遗传参数可知,3个分离世代(F2、B1和B2)籽粒周长的主基因遗传率分别为52.91%、42.91%和53.16%,说明遗传效应占主要作用,但受环境因素影响较大,在育种时应注重构建F2和B2分离世代进行选择。
粒长的最佳遗传模型为加性-显性-上位性多基因遗传模型(PG-ADI)。由二阶遗传参数可知,3个分离世代(F2、B1和B2)粒长的多基因遗传率分别为40.63%、37.08%和33.43%,说明粒长受环境因素影响也较大,但遗传因素也起到一定作用,在F2和B1分离世代中遗传因素影响稍大。因此,宜采用多环境鉴定的方法进行选择。
粒宽的最佳遗传模型为2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型(MX2-ADI-AD)。由一阶遗传参数可知,控制粒宽的2对主基因为正向加性效应,效应值均为0.09,显性效应均为负向,效应值分别为 -0.41和-0.33,显性×显性上位性效应为正向,为 0.71,大于其他遗传效应,说明2对主基因的遗传效应以负向显性效应为主,在高代基因纯合后进行选择可取得较好的效果。由二阶遗传参数可知,3个分离世代(F2、B1和B2)粒宽的主基因遗传率分别为60.24%、38.26%和53.14%,均远大于多基因遗传率(0.01%、0.01%和 8.10%),说明粒宽受主基因效应控制,同时也受环境因素影响,在F2和B2分离世代中具有较高的主基因遗传率,育种中应优先选择F2和B2分离世代。
3 讨 论
随着小麦基因组学[26]和快速加代技术[27]的发展,小麦育种效率逐步提高,而传统常规育种方法所采用的目测法效率低、工作量大、无法对表型进行量化分析[7],这成为小麦育种发展急需解决的问题。基于图像分析能够快速、准确、无损地获取表型信息。肖 杰等[24]采用SmartGrain软件法与游标卡尺测量法对小麦籽粒形态进行测量比较,结果显示,SmartGrain软件测量的粒长和粒宽的相对误差小于3%,且两种方法所测数据呈极显著线性相关。基于此,王 娜等[11]采用SmartGrain软件法解析了小麦NaN3诱变群体籽粒性状的遗传变异规律;张 健等[12]采用SmartGrain软件法采集了水稻RIL群体的籽粒表型数据,并定位了水稻籽粒大小相关性状QTL。本研究借鉴了前人的方法,采用SmartGrain软件对小麦籽粒图像进行处理分析,获得籽粒形态信息,同时应用于小麦育种工作,为小麦育种量化分析提供了指导。
小麦籽粒形态和千粒重是复杂的数量性状[13],籽粒面积、周长、粒长和粒宽是籽粒形态的重要决定因素,而大粒型小麦种质资源是小麦高产育种的重要基础。研究大粒型小麦新种质籽粒性状的遗传规律是提高高产育种效率的重要前提,为合理构建选择群体和指导后代选择提供重要依据。本研究对大粒型小麦新种质漯麦76和普通小麦种质L529构建的4个世代6个群体进行遗传分析,结果表明,千粒重最佳模型为两对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型,这与杨兴圣等[21]的研究结果一致,但2对主基因的遗传效应有所不同,本研究中控制千粒重的2对主基因是负向等加性效应。本研究发现,籽粒面积和粒长的最佳模型均为加性-显性-上位性多基因遗传模型,籽粒面积和粒长的多基因遗传率分别为24.37%~44.81%和33.43%~40.63%,受环境影响较大。余曼丽等[28]分别检测到有5和12个QTL控制籽粒面积和粒长;杨兴圣等[21]研究表明,粒长属于加性-显性-上位性多基因遗传模型,这与本研究结果一致;而陈佳慧等[29]检测到粒长受2个QTL控制。原因有待进一步研究。本研究中籽粒周长的最佳模型为两对加性-显性-上位性主基因遗传模型。余曼丽等[28]检测到有10个QTL控制籽粒周长,但有2个QTL能够在两年环境中检测到。本研究中粒宽的最佳模型为两对加性-显性-上位性主基因+加性显性多基因遗传模型,与杨兴圣等[21]的研究结果一致。
4 结 论
通过对大粒型小麦新种质漯麦76配制的4个世代6个联合群体进行遗传分析,结果表明,千粒重的最佳模型为MX2-ADI-ADI(两对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型),2对主基因的遗传效应以负向等加性效应为主,以负向的显性×显性上位性效应为辅,说明千粒重性状受低粒重亲本影响较大,宜采用“高粒重×高粒重”的组合配制方案;籽粒面积和粒长的最佳模型均为PG-ADI(加性-显性-上位性多基因遗传模型),受环境因素影响较大,宜采用多年多点的种植方案进行选择;籽粒周长的最佳模型为2MG-ADI(两对加性-显性-上位性主基因遗传模型),2对主基因以负向的显性效应为主,正向的显性×显性上位性效应为辅,宜在高代基因纯合后进行选择;粒宽的最佳模型为MX2-ADI-AD(两对加性-显性-上位性主基因+加性显性多基因遗传模型),2对主基因以负向的显性效应为主,正向的显性×显性上位性效应为辅,同时主基因遗传率远大于多基因遗传率,宜在高代基因纯合后进行选择。综上,对籽粒性状的选择,应采用“高粒重×高粒重”的组合配制方案构建F2和B2选择群体,多环境鉴定,以低代宽高代严的选择标准,在高代对大粒性状进行选择。
猜你喜欢
阜阳师范大学学报(自然科学版)(2022年1期)2022-04-02
广东农业科学(2021年3期)2021-04-23
系统管理学报(2018年3期)2018-08-13
系统管理学报(2018年2期)2018-08-13
消费导刊(2017年24期)2018-01-31
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21
广东农业科学(2016年11期)2016-03-29
华北农学报(2016年1期)2016-03-18
农业科技与装备(2015年5期)2015-05-30
哈尔滨师范大学自然科学学报(2015年1期)2015-04-19
