
不同氮效率小麦品种苗期相关指标的杂种优势
2023-04-25 09:10王汉霞马巧云单福华田立平张胜全
麦类作物学报 2023年4期
王汉霞,马巧云,单福华,田立平,权 威,张胜全
(北京市农林科学院杂交小麦研究所/杂交小麦分子遗传北京市重点实验室,北京 100097 )
小麦是人类主要粮食作物之一,在世界各地被广泛种植。氮是作物生长发育不可或缺的营养元素,也是影响作物代谢的限制因素之一。人们为了提高作物产量而投入大量的氮肥到土壤中,张福锁等[1]研究表明,我国大多数农业生产区域施氮量过多,造成土壤和水污染,甚至使作物产量降低。因此,提高氮肥利用效率、选育氮高效小麦品种至关重要。关于小麦氮效率的研究已有较多报道,如张国平等[2]研究表明,不同基因型小麦的氮利用效率存在差异;赵 瑞等[3]对108份小麦的氮效率进行研究,发现不同基因型成株期的氮效率存在差异。前人研究发现,在特定氮素水平下,小麦的产量、群体质量与氮效率存在正相关关系[4];小麦单株总根长、根总表面积、单株根尖数、茎叶干重、根干重、叶面积、全氮含量可以作为小麦苗期不同氮效率类型的筛选指标[5-7];低氮胁迫使小麦叶片和根部的硝酸还原酶(NR)、谷氨酰胺合成酶(GS)活性及氮含量和氮积累量均降低[8]。
Kant等[9]和侯文通等[10]认为,选育氮高效品种可提高小麦氮肥利用效率。王琳琳等[11]发现,低氮高效和高氮高效小麦品种杂交,低氮环境有利于选择优良后代。我国小麦氮高效利用的研究主要集中在不同氮素利用效率品种的评价方法及指标的筛选,对不同氮水平下,不同氮效率品种的杂种优势研究较少。本研究选用5个不同氮效率小麦品种及其组配的6个杂交F1,在两个氮水平下对其苗期形态及氮代谢相关酶的活性进行分析,探讨不同氮效率小麦及其杂交F1苗期相关指标的差异,以期为小麦氮高效育种的亲本选配提供理论依据。
1 材料与方法
1.1 供试材料
供试材料为本课题组前期筛选出的5个不同氮效率的小麦品种京冬8(JD8,氮低效)、矮抗58(AK58,氮低效)、京农18-8(JN18-8,氮高效)、京农18-96(JN18-96,氮高效)、郑麦366(ZM366,氮高效)及将其作为亲本组配成的6个杂交F1,组合分别为JN18-8/JD8、JN18-8/AK58、JN18-96/JD8、JN18-96/AK58、ZM366/JD8、ZM366/AK58。
1.2 幼苗培养方法
幼苗采用水培培养,设置两个氮水平:低氮(LN,0.4 mmol·L-1)和正常供氮(CK, 4.0 mmol·L-1),氮源为Ca(NO3)2。各材料均取100粒种子用5%的H2O2消毒5 min,用去离子水洗净;置于培养盒中,于光照培养箱中去离子水培养至一叶一心;将幼苗移入氮水平为0.2和2.0 mmol·L-1的1/2 Hoagland营养液中缓苗2 d,后分别转入氮含量0.4和4.0 mmol·L-1的Hoagland营养液中,继续培养25 d,期间每隔3 d换一次营养液,用电动气泵连续通气,每个处理三次重复。试验在光照培养室内进行,光照强度为 4.8×104Lx,光/暗时间为16 h/8 h,温度为 25 ℃/20 ℃,相对湿度为60%。
1.3 取样方法
将1.2中培养25 d的幼苗从培养盒中取出,用流动的去离子水冲洗干净,用吸水纸吸干附着水;每个处理取10株材料,按根、苗分开,测定鲜重;于105 ℃的烘箱中杀青30 min后80 ℃烘干至恒重,测定根、苗干重后,用于氮含量的测定。
按上述方法每个处理取样10株,分别取叶片和根,液氮速冻后置于-80 ℃保存,用于氮代谢相关酶活性的测定。
1.4 测定指标与方法
氮含量:采用半微量凯氏定氮法测定[12]。
采用南京建成生物工程研究所生产的试剂盒测定超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、硝酸还原酶(NR)活性、谷氨酰胺合成酶(GS)活性、谷氨酸脱氢酶(GDH)活性、丙二醛(MDA)含量。
可溶性蛋白含量:采用考马斯亮蓝G-250法测定[13]。
1.5 数据统计与分析
中亲优势(mid-parent heterosis,MH)=(F1值-双亲平均值)/ 双亲平均值×100%;超高亲优势(better parent heterosis,BH)=(F1值-高值亲本值)/高值亲本值×100%;超低亲优势(lower parent heterosis,LH)=(F1值-低值亲本值)/低值亲本值×100%;偏低亲优势表示F1杂种优势大于超低亲优势但小于中亲优势;若F1值超过亲本均值,为正杂种优势,若低于亲本均值,则为负杂种优势。
数据用Excel 2007和DPS v 7.05进行分析。
2 结果与分析
2.1 不同氮水平下小麦苗期植株干鲜重的遗传分析
由表1可知,正常氮水平下小麦苗鲜重均高于低氮水平,不同材料差异程度不同;两个氮水平下,组合ZM366/AK58和JN18-8/AK58的F1均表现出正向杂种优势,其中组合ZM366/AK58的F1在正常供氮水平下表现出超亲优势;JN18-96/AK58的F1在正常供氮水平下表现中亲优势,组合JN18-8/JD8和JN18-96/JD8的F1在低氮水平下表现出中亲优势。
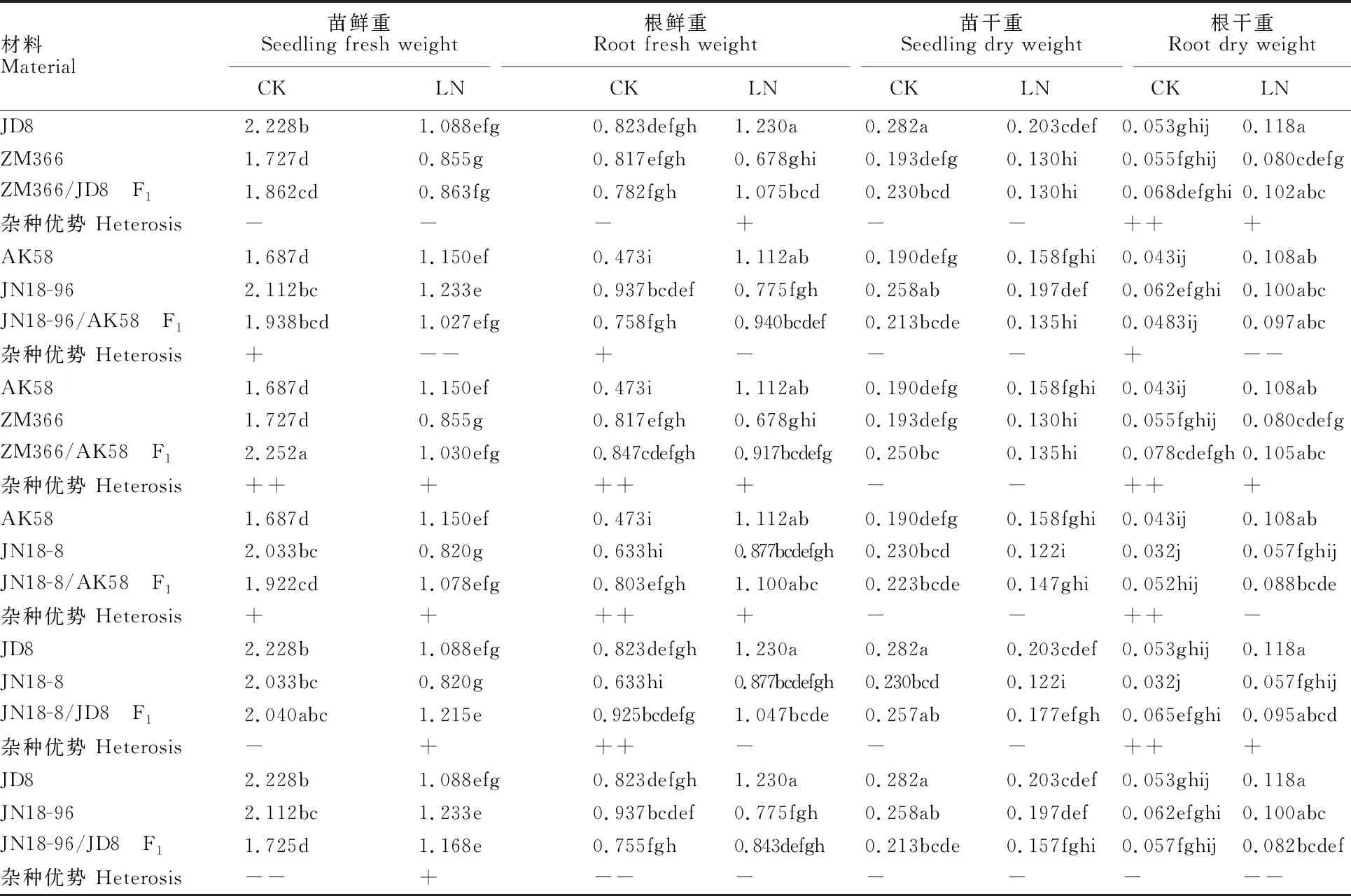
表1 不同氮水平下小麦苗期植株干鲜重及杂种优势
大部分供试材料在低氮条件下的根鲜重高于正常氮水平,不同基因型的增减幅度不同,其中,组合JN18-8/JD8的F1在两个氮水平下均表现为超亲优势,其他组合在低氮水平下表现为超中亲或偏低亲优势。6个F1中,仅有2个F1(JN18-8/AK58和JN18-8/JD8)的苗干重同时在两个氮水平下表现出超中亲优势;有3个F1(ZM366/JD8 、ZM366/AK58和JN18-8/JD8)的根干重同时在两个氮水平下表现出超中亲优势或超高亲优势。ZM366/AK58和 JN18-8/AK58的F1表现出较好的根、苗干重和鲜重杂种优势。
2.2 不同氮水平小麦苗期氮含量与氮积累量的遗传分析
由表2可以看出,低氮水平下,各材料地上部含氮量较正常供氮水平降低1.5~1.7倍,根部氮含量降低1.8~2.1倍;除组合ZM366/JD8外,其他组合F1的地上部含氮量均表现正向杂种优势。低氮较正常供氮水平,各供试材料地上部氮积累量降低1.9~3.2倍,根部氮积累量降低0.9~1.4倍。组合ZM366/JD8、ZM366/AK58和JN18-8/D8的F1地上部和根的氮积累量均表现正向杂种优势。正常氮水平下,组合JN18-96/AK58的F1地上部和根的氮积累量均表现为超中亲优势;在低氮条件下,组合JN18-8/JD8的F1地上部和根的氮积累量均表现为超中亲优势。所有组合中,只有JN18-96/JD8的F1地上部和根的氮积累量在低氮条件下表现为超低亲杂种优势。
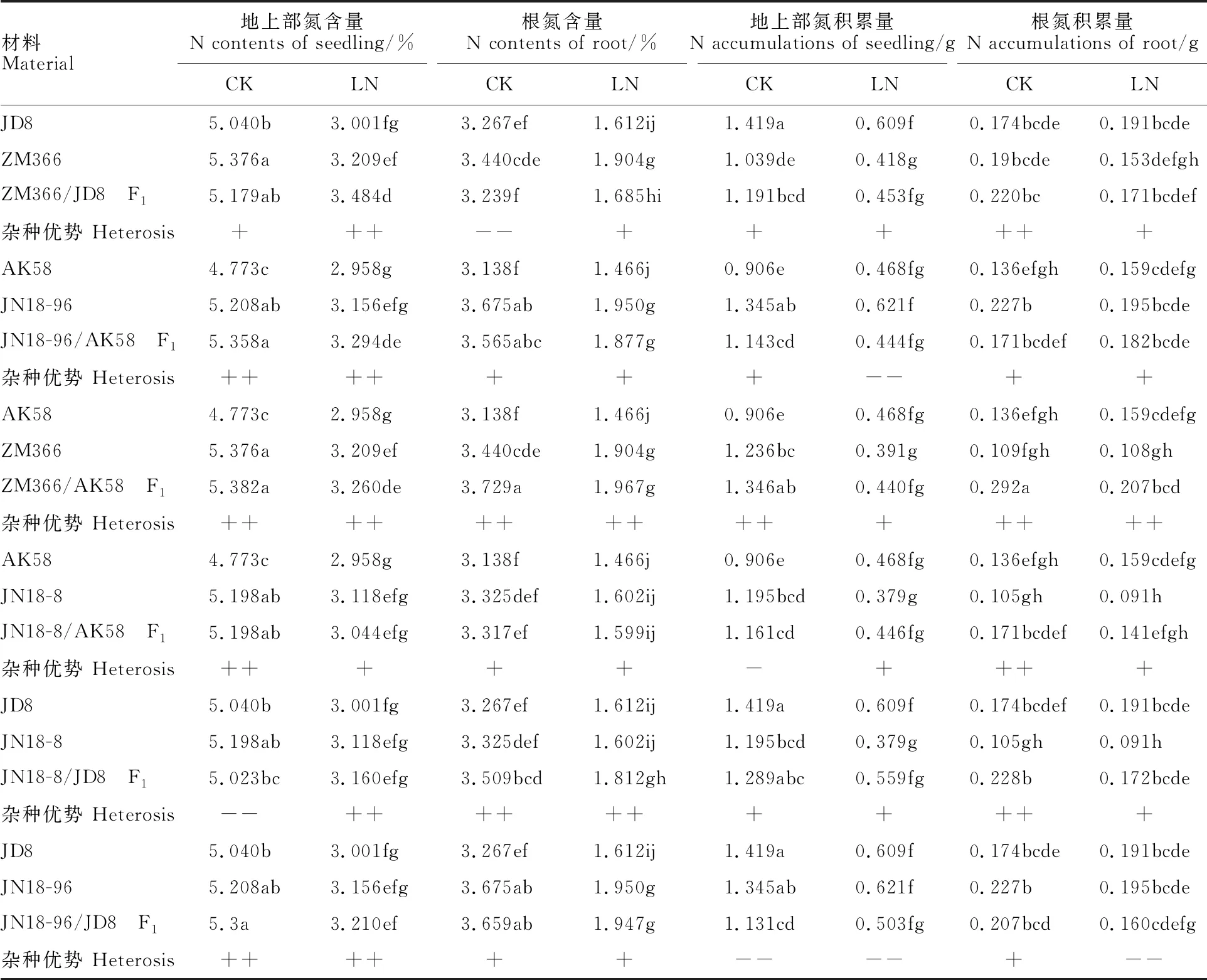
表2 不同氮水平下小麦苗期植株氮含量、氮积累量及杂种优势
2.3 不同氮水平下小麦苗期氮代谢相关酶活性的遗传分析
硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)是植物叶片涉及氮代谢的主要同化酶。由表3可见,正常氮水平下,各供试材料叶片中GS、GDH和NR活性整体高于低氮水平;除ZM366/JD8的F1中NR活性表现为偏低亲优势外,其他组合F1叶片中的GS、GDH和NR活性均表现超中亲甚至超高亲优势。低氮水平下,除JN18-8/AK58的F1中GS和GDH活性表现低亲优势外,其他组合F1的GS和GDH活性和NR活性均表现超亲或者超高亲优势。氮高效品种叶片氮代谢酶活性均高于氮低效品种。
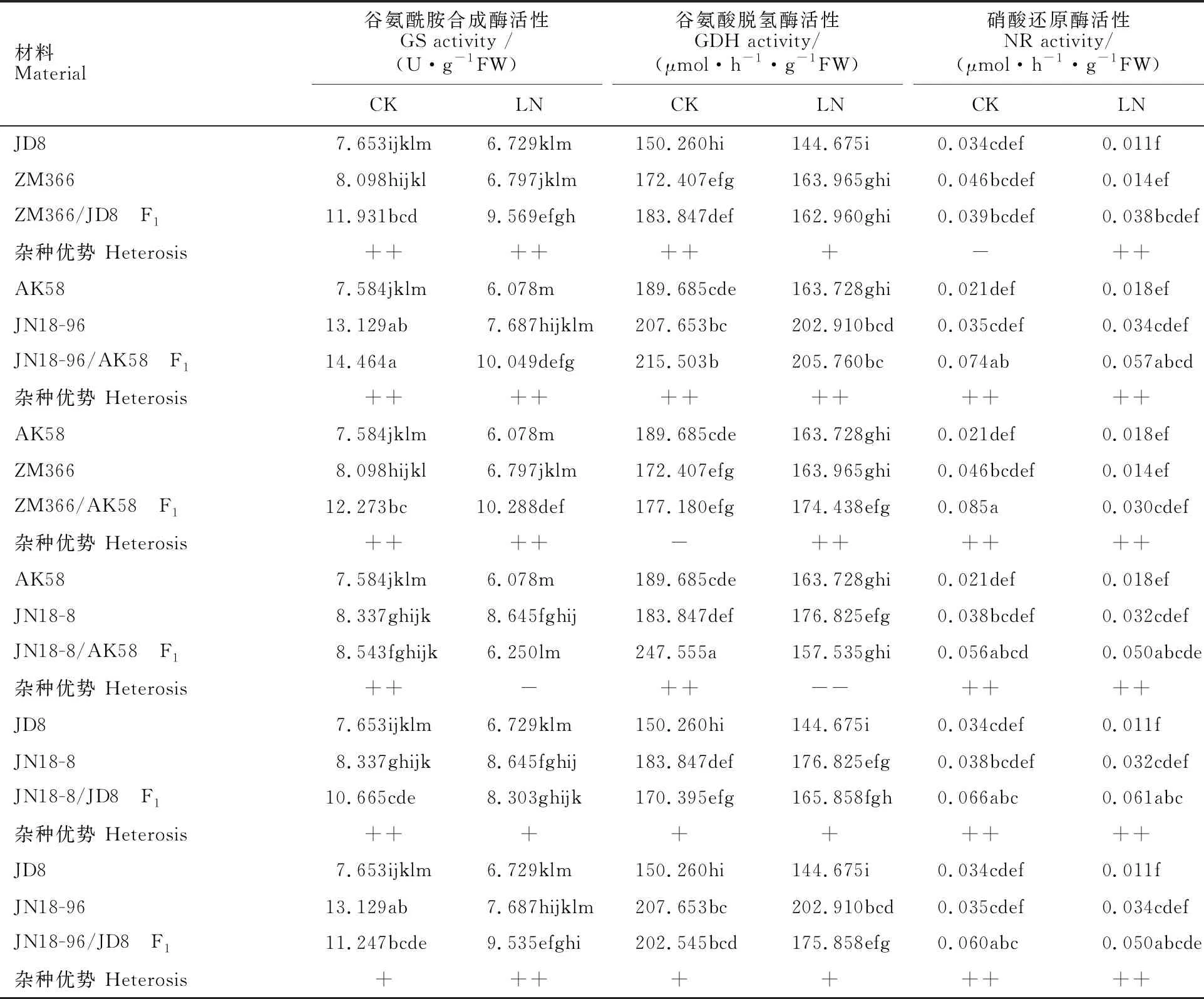
表3 不同氮水平下小麦苗期叶片氮代谢相关酶活性及杂种优势
由表4可知,正常氮水平下各供试材料根系GS、GDH和NR活性高于低氮水平。正常氮水平下,除组合JN18-8/AK58、JN18-8/JD8的F1中GDH活性表现为低亲优势外,其他组合F1的GS、GDH和NR活性均表现超中亲甚至超高亲优势。低氮水平下,除ZM366/JD8的F1根系中NR活性、JN18-96/AK58与ZM366/AK58的F1中GDH、NR活性表现低亲优势外,其他组合F1根系中的GS、GDH和NR活性均表现超中亲或者超高亲优势。氮高效品种根系GS、GDH活性高于氮低效品种。
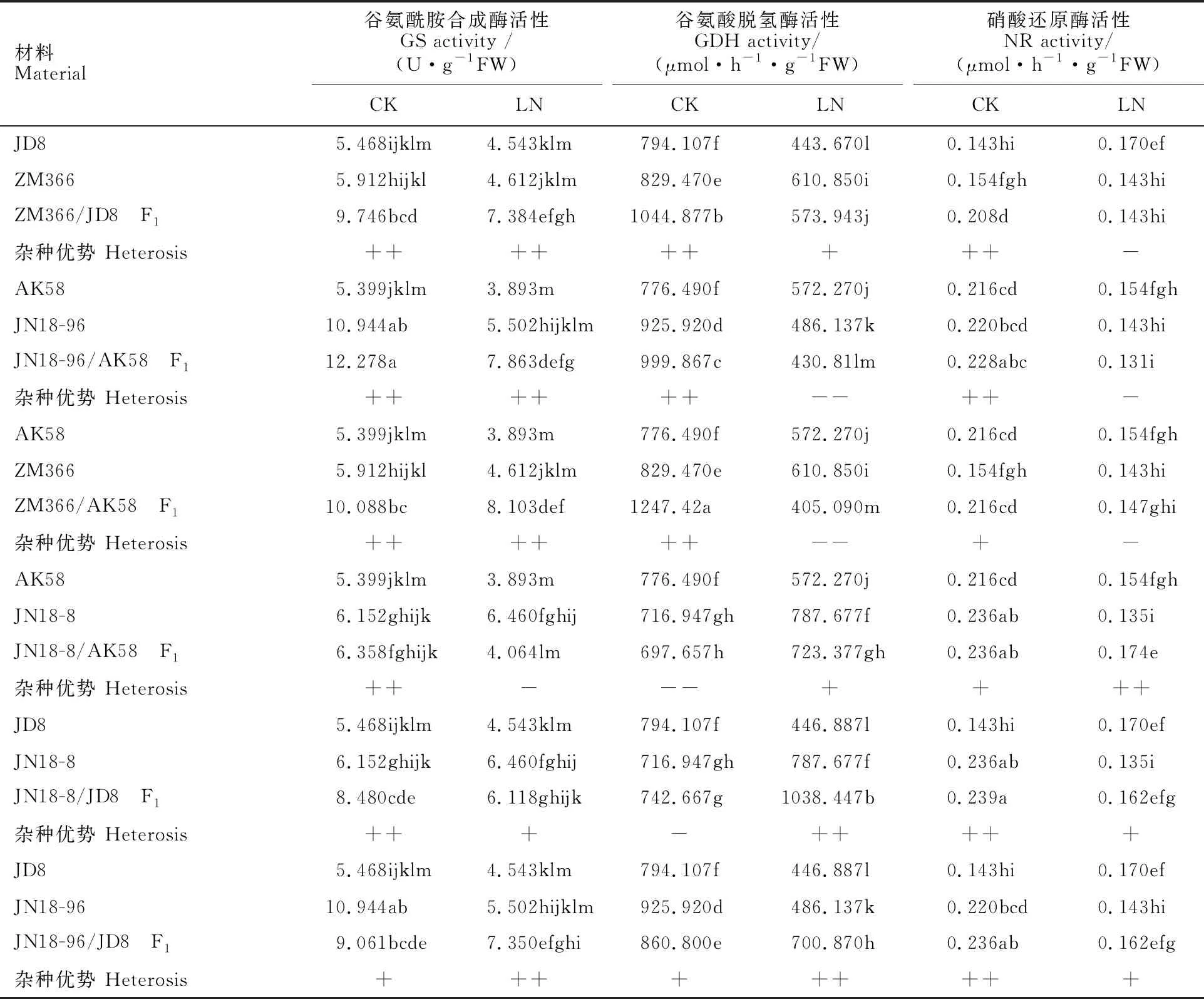
表4 不同氮水平下小麦苗期根系氮代谢相关酶活性及杂种优势
2.4 不同氮水平下小麦苗期抗氧化酶活性的遗传分析
SOD、POD、CAT是生物防御活性氧伤害的重要保护酶类,能有效的阻止活性氧的快速积累。由表5可以看出,正常氮水平下叶片中SOD、POD、CAT活性高于低氮水平,说明氮素水平升高可提高小麦幼苗叶片内的抗氧化酶活性。正常氮水平下,组合JN18-96/AK58、ZM366/AK58、 JN18-96/JD8的F1叶片中SOD活性表现为低亲优势,组合ZM366/JD8的F1的叶片POD活性表现为超低亲优势,其他组合F1的SOD活性、POD活性、CAT活性均表现为超亲优势或者超高亲优势。低氮条件下,除组合JN18-96/JD8的F1叶片POD活性表现为低亲优势外,其他组合F1的SOD、POD、CAT活性均表现为超亲优势或者超高亲优势。
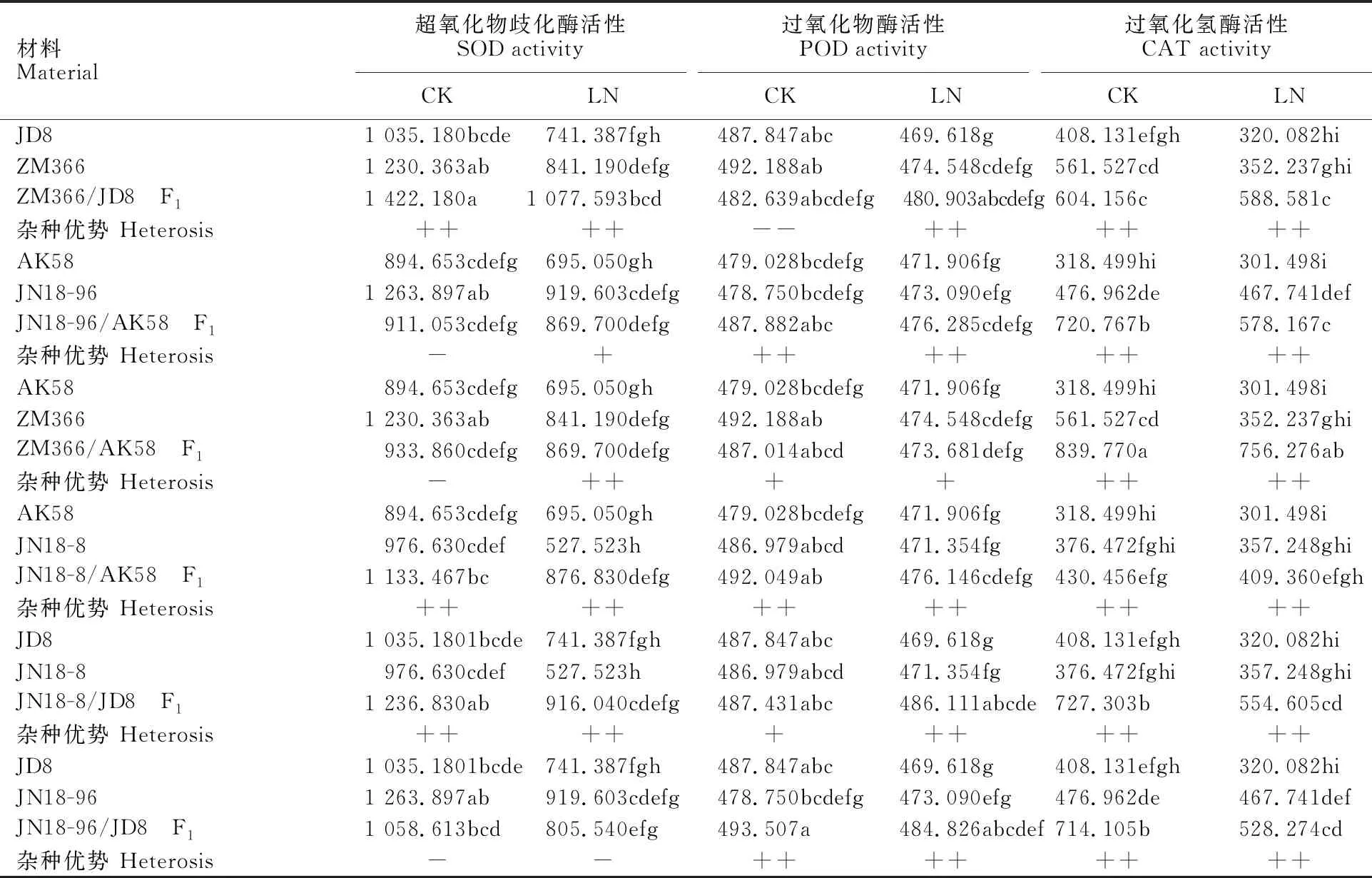
表5 不同氮水平下小麦苗期叶片抗氧化酶活性及杂种优势分析
由表6可以看出,正常氮水平下根系中SOD、POD、CAT活性高于低氮水平,说明氮素水平升高可提高小麦幼苗根系内抗氧化酶活性。正常氮水平下,杂交组合JN18-96/AK58的F1根系SOD活性、ZM366/AK58和JN18-8/AK58的F1根系SOD和POD活性、 JN18-8/JD8和JN18-96/JD8的F1根系的POD活性表现为低亲或超低亲优势外,其他组合F1的SOD、POD、CAT活性均表现为超亲优势或者超高亲优势。低氮条件下,组合ZM366/JD8、JN18-96/AK58、ZM366/AK58、JN18-8/AK58的F1根系SOD活性、JN18-96/JD8的F1根系POD活性、JN18-96/AK58和JN18-96/JD8的F1根系CAT活性表现为低亲优势或超低亲优势,其他组合F1的SOD、POD、CAT活性均表现为超亲优势或者超高亲 优势。
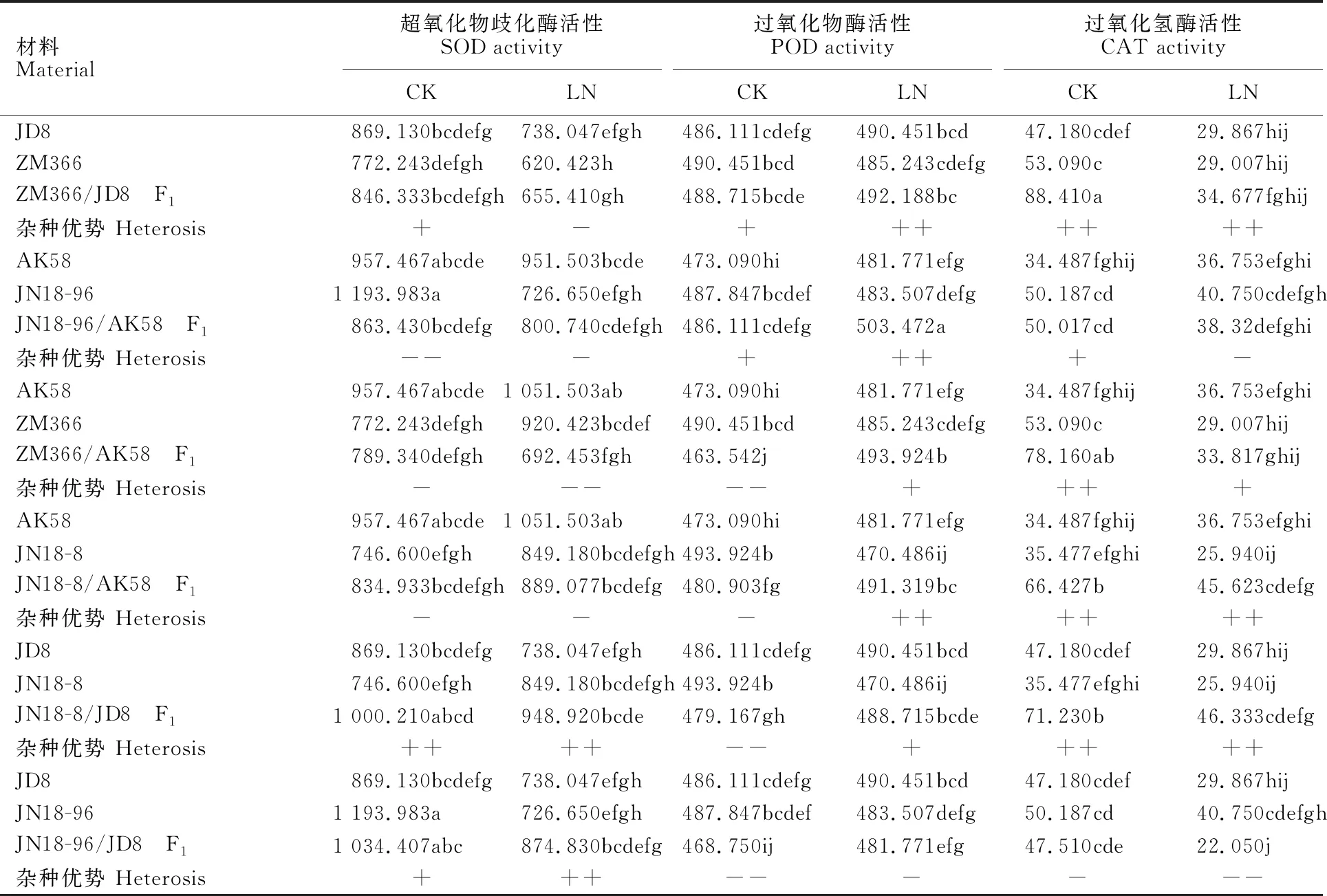
表6 不同氮水平下小麦苗期根系抗氧化酶活性及杂种优势分析
2.5 不同氮水平下小麦苗期叶片MDA含量与可溶性蛋白含量遗传分析
由表7可知,在低氮水平下叶片MDA含量较正常氮水平提高,且氮低效品种JD8的MDA含量最高,氮高效品种JN18-8的MDA含量最低,说明氮高效品种在低氮条件下能维持细胞膜的稳定状态。正常氮水平下,组合JN18-96/AK58、JN18-8/AK58的F1叶片MDA含量表现为偏低亲优势,组合ZM366/AK58、JN18-8/JD8的F1表现为超中亲优势,组合ZM366/JD8、JN18-96/JD8的F1表现为超高亲优势,但F1与其亲本间差异不显著。低氮条件下,组合ZM366/AK58的F1叶片MDA含量表现为超高亲优势,组合JN18-8/AK58、 JN18-8/JD8的F1表现为超中亲优势,组合ZM366/JD8、JN18-96/JD8的F1表现为偏低亲优势,组合JN18-96/AK58的F1表现为超低亲优势。
由表7可以看出,正常氮水平下,各供试材料叶片的可溶性蛋白含量高于低氮水平;除组合JN18-96/JD8的F1外,其他组合F1的叶片可溶性蛋白含量均表现出超中亲或超高亲优势。低氮条件下,氮高效品种的叶片可溶性蛋白含量稍高于氮低效品种,6个F1叶片的可溶性蛋白含量均表现超高亲优势。

表7 不同氮水平下小麦苗期叶片MDA、可溶性蛋白含量及杂种优势
由表8可知,正常条件下,6个F1的根系MDA 含量表现为中亲、低亲或超低亲优势。在低氮条件下,供试材料根系的MDA含量增加,且氮低效品种根系的MDA含量较氮高效品种高,说明氮低效品种受到的伤害大,6个F1均表现为中亲或低亲优势。
由表8可以看出,正常条件下,除组合JN18-96/AK58的F1外,其他组合F1的根系的可溶性蛋白含量均表现出超中亲或超高亲优势,与叶片表现一致。在低氮条件下,除JN18-96/JD8外,其他组合的F1根系可溶性蛋白含量表现为偏低亲或者超低亲优势。
3 讨论与结论
氮素是作物生长发育的重要营养元素之一,选育氮高效小麦品种是提高小麦氮效率、实现减氮丰产的有效措施[14-17]。前人研究表明,不同基因型小麦具有不同的氮利用效率,低氮胁迫下,不同基因型小麦均表现出地上部生物量降低、根部生物量升高,且氮高效品种的根部生物量增幅更大[18-19];苗期根系的主要形态数量性状可作为衡量氮效率的筛选指标[7]。本研究表明,低氮水平下,各基因型小麦的根系鲜重高于正常氮水平下,这与前人的研究结果一致,这说明低氮可诱导根系的生长,在一定程度上促进根系对氮素的吸收,缓解氮胁迫对植株生长的不良影响。本研究中,组合ZM366/AK58、JN18-8/AK58的F1叶片和根系鲜重均表现为超中亲或超高亲优势。
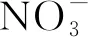
在氮胁迫下,小麦会产生更多的活性氧使小麦细胞膜脂过氧化,损伤细胞膜,抗氧化酶能快速清除细胞中的活性氧,保护细胞膜受到损伤[28]。有研究表明,小麦旗叶的SOD、POD、CAT活性在一定的范围内随施氮水平的增加而提高[29-30]。本研究表明,氮高效小麦品种叶片和根系的抗氧化酶活性高于氮低效小麦品种,更利于细胞完整性的维持;组合JN18-8/AK58和JN18-8/JD8的F1叶片中三种抗氧化酶活性在两个氮素水平下的均表现出超中亲或超高亲优势。
MDA含量是植物膜脂过氧化程度的体现,本研究发现,在低氮条件下,小麦MDA含量增加,且氮低效品种的MDA含量大于氮高效小麦品种,说明氮低效小麦品种的细胞膜受的伤害较大。可溶性蛋白是细胞重要的渗透调节物质,本研究中,叶片可溶性蛋白含量在正常施氮条件下高于低氮,且在低氮条件下,氮高效小麦品种的可溶性蛋白含量高于氮低效小麦品种,组合ZM366/AK58和JN18-8/JD8的F1叶片的MDA和可溶性蛋白含量均表现出了超中亲或超高亲优势,组合JN18-96/JD8的F1根系的MDA和可溶性蛋白含量均表现出了超中亲或超高亲优势。说明利用氮效率杂种优势配置杂交组合可以作为培育小麦氮高效小麦新品种的一种途径。
王琳琳等[11]研究发现,不同氮效率小麦品种配置的杂交组合在不同氮水平下,大多数指标表现为正向杂种优势。本研究中,6个杂交组合F1叶片的NR、GS、GDH、SOD、POD、CAT活性、可溶性蛋白含量大多存在中亲或高亲优势,根系的NR、GS、GDH、CAT活性、MDA含量大多存在中亲或高亲优势。正常施用氮肥能够有效提高小麦苗期叶片SOD、POD、CAT活性,增加可溶性蛋白含量,降低膜脂过氧化程度,提高NR、GS、GDH活性。在低氮条件下,氮高效品种比氮低效品种生长的更好,杂交F1也表现出了较好的耐低氮性,所以以氮高效小麦品种作为亲本配制杂交组合可以作为氮高效育种的有效方法。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
蔬菜(2021年7期)2021-11-27
今日农业(2021年9期)2021-11-26
四川劳动保障(2021年3期)2021-06-09
今日农业(2020年23期)2020-12-15
今日农业(2019年12期)2019-08-13
现代园艺(2017年13期)2018-01-19
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
山西农经(2016年5期)2016-02-28
